
Supplementation with Fermented Feedstuff Enhances Orexin Expression and Secretion Associated with Increased Feed Intake and Weight Gain in Weaned Pigs

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Fermented Feedstuff (FFS)
2.2. Experimental Design, Animals and Diets
2.3. Sampling
2.4. Growth Performance Estimation
2.5. Apparent Digestibility Estimation
2.6. Plasma Biochemical Profile Detection
2.7. Measurement of Serum Hormones
2.8. Extraction of RNA and cDNA Synthesis
2.9. Real-Time Fluorescence PCR for Quantification of mRNA Levels
2.10. Statistical Analysis
3. Results
3.1. The Effects of FFS on Growth Performance of Pigs
3.2. The Effects of FFS on the Apparent Nutrient Digestibility of Pigs
3.3. The Effects of FFS on the Serum Biochemical Parameters of Pigs
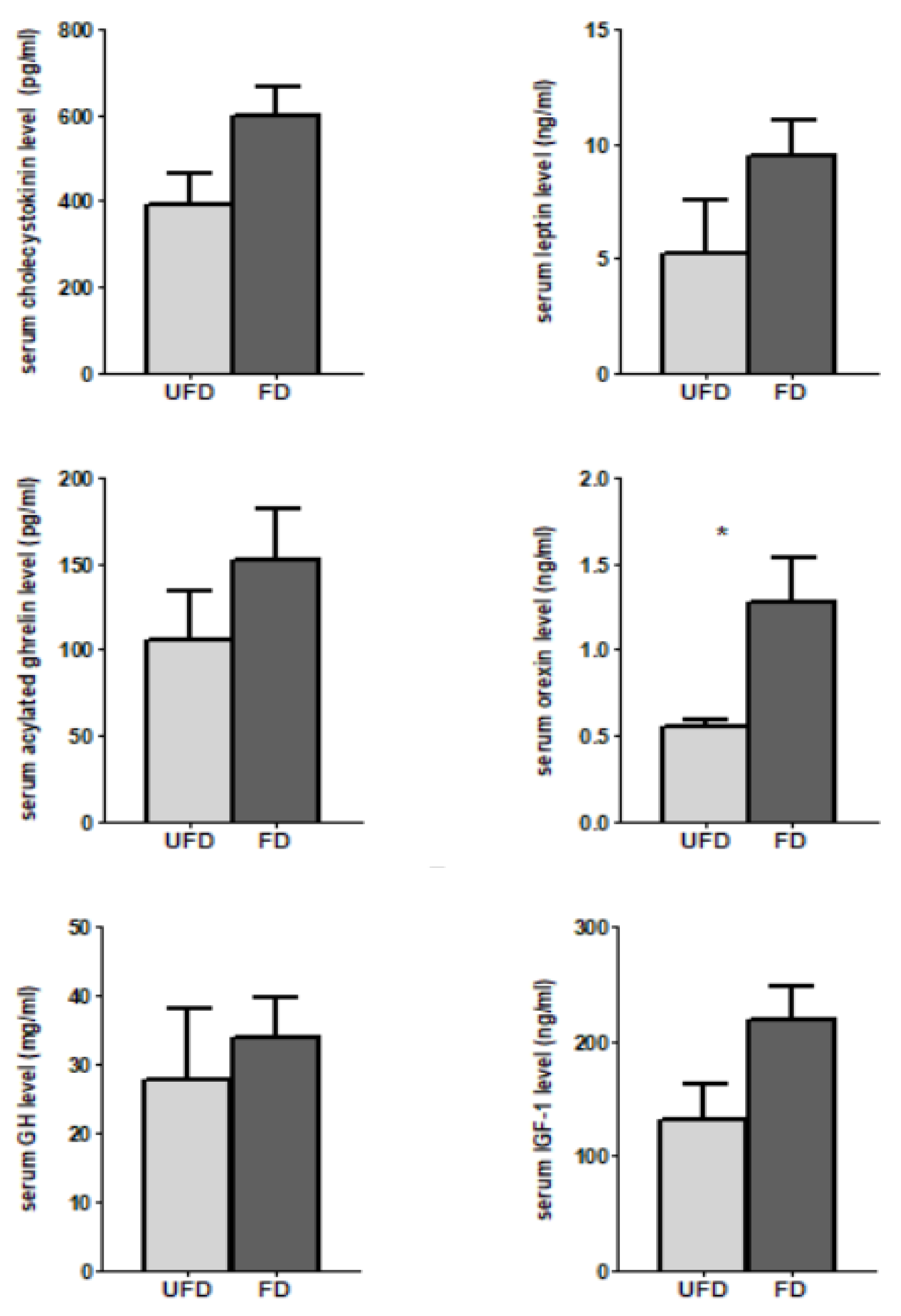
3.4. The Effects of FFS on the Serum Hormones Associated with Feed Intake and Growth of Pigs
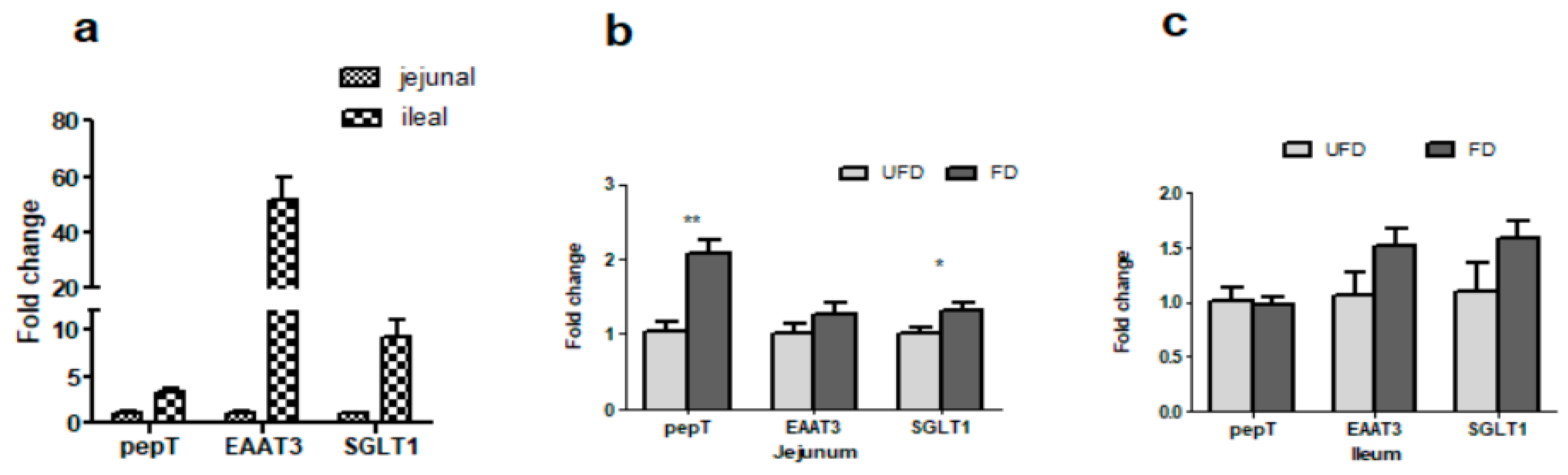
3.5. The Effects of FFS on the Gene Expressions of Nutrient Transporters of Pigs
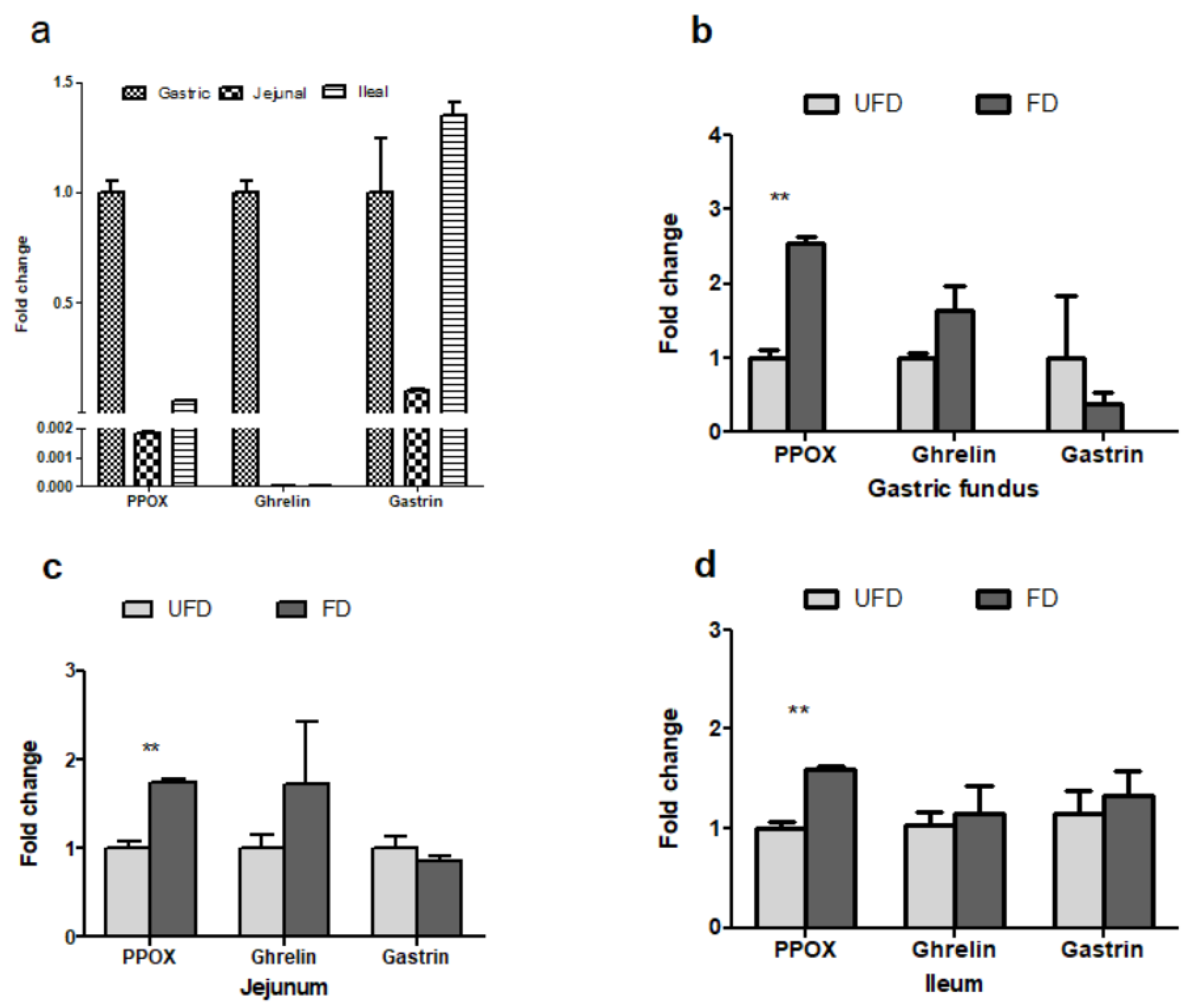
3.6. The Effects of FFS on the Gene Expressions Associated with Feed Intake of Pigs
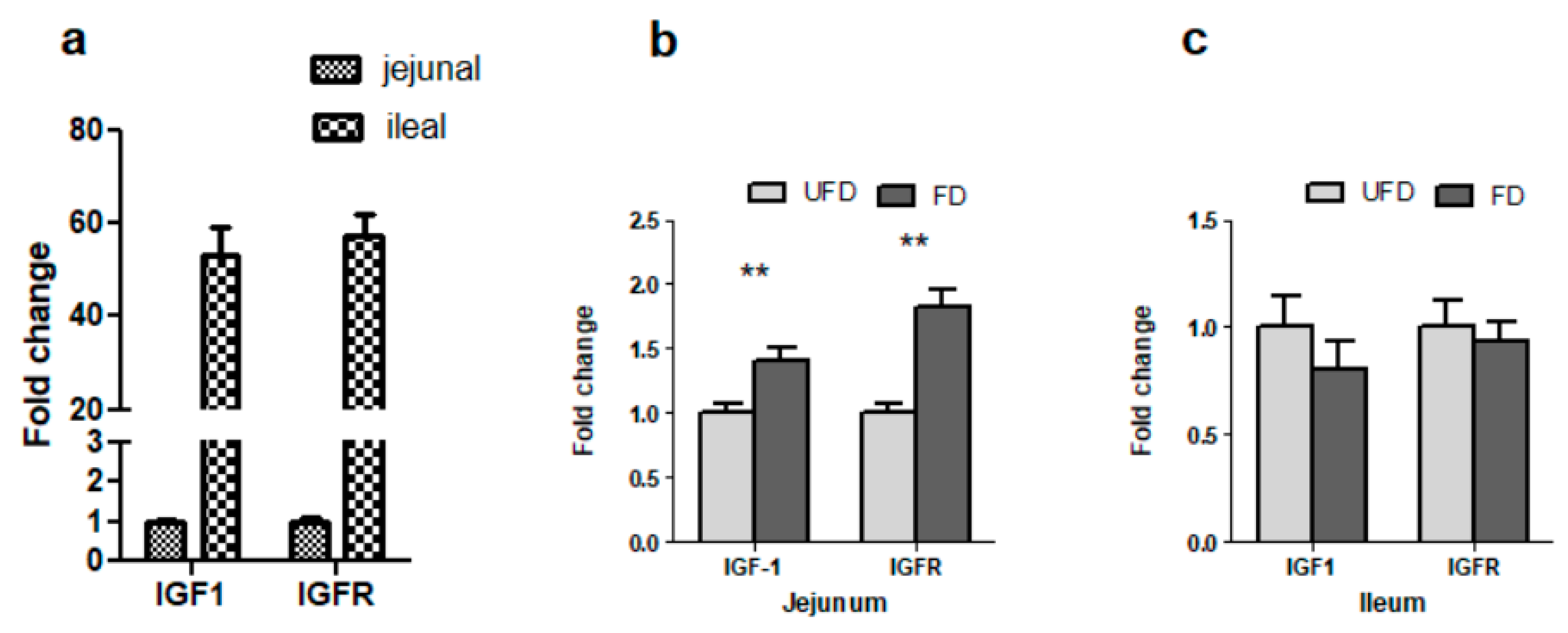
3.7. The Effects of FFS on the Gene Expressions of IGF-1 and IGFR in the Intestinal Mucosa of Pigs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Humer, E.; Wetscherek, W.; Schwarz, C.; Schedle, K. Effects of maize conservation techniques on the apparent total tract nutrient and mineral digestibility and microbial metabolites in the faeces of growing pigs. Anim. Feed Sci. Technol. 2014, 197, 176–184. [Google Scholar] [CrossRef]
- Koo, B.; Kim, J.W.; Nyachoti, C.M. Nutrient and energy digestibility, and microbial metabolites in weaned pigs fed diets containing Lactobacillus–fermented wheat. Anim. Feed Sci. Technol. 2018, 241, 27–37. [Google Scholar] [CrossRef]
- Yuan, L.; Chang, J.; Yin, Q.; Lu, M.; Di, Y.; Wang, P.; Wang, Z.; Wang, E.; Lu, F. Fermented soybean meal improves the growth performance, nutrient digestibility, and microbial flora in piglets. Anim. Nutr. 2017, 3, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Sholly, D.M.; Jorgensen, H.; Sutton, A.L.; Richert, B.T.; Bach Knudsen, K.E. Effect of fermentation of cereals on the degradation of polysaccharides and other macronutrients in the gastrointestinal tract of growing pigs. J. Anim. Sci. 2011, 89, 2096–2105. [Google Scholar] [CrossRef]
- Meinlschmidt, P.; Ueberham, E.; Lehmann, J.; Schweiggert-Weisz, U.; Eisner, P. Immunoreactivity, sensory and physicochemical properties of fermented soy protein isolate. Food Chem. 2016, 205, 229–238. [Google Scholar] [CrossRef]
- Scholten, R.H.J.; van der Peet-Schwering, C.M.C.; Verstegen, M.W.A.; den Hartog, L.A.; Schrama, J.W.; Vesseur, P.C. Fermented co-products and fermented compound diets for pigs: A review. Anim. Feed Sci. Technol. 1999, 82, 1–19. [Google Scholar] [CrossRef]
- Sugiharto, S.; Lauridsen, C.; Jensen, B.B. Gastrointestinal ecosystem and immunological responses in E. coli challenged pigs after weaning fed liquid diets containing whey permeate fermented with different lactic acid bacteria. Anim. Feed Sci. Technol. 2015, 207, 278–282. [Google Scholar] [CrossRef]
- Xu, B.; Zhu, L.; Fu, J.; Li, Z.; Wang, Y.; Jin, M. Overall assessment of fermented feed for pigs: A series of meta-analyses. J. Anim. Sci. 2019, 97, 4810–4821. [Google Scholar] [CrossRef]
- Geary, N. Endocrine controls of eating: CCK, leptin, and ghrelin. Physiol. Behav. 2004, 81, 719–733. [Google Scholar] [CrossRef]
- Barson, J.R. Orexin/hypocretin and dysregulated eating: Promotion of foraging behavior. Brain Res. 2020, 1731, 145915. [Google Scholar] [CrossRef]
- Al Massadi, O.; Nogueiras, R.; Dieguez, C.; Girault, J.-A. Ghrelin and food reward. Neuropharmacology 2019, 148, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Laviano, A.; Di Lazzaro, L.; Koverech, A. Changes in eating behavior, taste and food preferences and the effects of gastrointestinal hormones. Clin. Uutr. Exp. 2018, 20, 65–70. [Google Scholar] [CrossRef]
- Moss, C.; Dhillo, W.S.; Frost, G.; Hickson, M. Gastrointestinal hormones: The regulation of appetite and the anorexia of ageing. J. Hum. Nutr. Diet. 2012, 25, 3–15. [Google Scholar] [CrossRef]
- Strader, A.D.; Woods, S.C. Gastrointestinal hormones and food intake. Gastroenterology 2005, 128, 175–191. [Google Scholar] [CrossRef] [PubMed]
- de Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.; Gautvik, V.T.; Bartlett, F.S.; et al. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc. Natl. Acad. Sci. USA 1998, 95, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Naslund, E.; Ehrstrom, M.; Ma, J.; Hellstrom, P.M.; Kirchgessner, A.L. Localization and effects of orexin on fasting motility in the rat duodenum. Am. J. Physiol. Gastrt. Liver Physiol. 2002, 282, G470-9. [Google Scholar] [CrossRef] [Green Version]
- Dall’Aglio, C.; Zannoni, A.; Forni, M.; Bacci, M.L.; Ceccarelli, P.; Boiti, C. Orexin system expression in the gastrointestinal tract of pigs. Res. Vet. Sci. 2013, 95, 8–14. [Google Scholar] [CrossRef]
- Sales, J.; Janssens, G.P. Acid-insoluble ash as a marker in digestibility studies: A review. J. Anim. Feed Sci. 2003, 12, 383–401. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis; AOAC: Washington, DC, USA, 2005. [Google Scholar]
- GB/T 6437. Determination of Total Phosphorus in Feed-Spectrophotometry; Standardization Administration of China: Beijing, China, 2018. [Google Scholar]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dall’Aglio, C.; Zannoni, A.; Mercati, F.; Forni, M.; Bacci, M.L.; Boiti, C. Differential gene expression and immune localization of the orexin system in the major salivary glands of pigs. Regul. Peptides 2011, 172, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Bedford, A. The Effect of Lactococcus Lactis-Produced Epidermal Growth Factor on the Growth and Intestine Development of Early-Weaned Pigs; The University of Guelph: Guelph, ON, Canada, 2014. [Google Scholar]
- Ye, J.; Gao, C.; Li, X.; Jin, C.; Wan, D.; Shu, G.; Wang, W.; Kong, X.; Yao, K.; Yan, H.; et al. EAAT3 promotes amino acid transport and proliferation of porcine intestinal epithelial cells. Oncotarget 2016, 7, 38681–38692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Wang, L.; Yi, H.; Wang, Z.; Yang, X.; Gao, K.; Wen, X.; Jiang, Z. Effects of Lactobacillus reuteri LR1 on serum biochemical parameters and intestinal nutrient transporter mRNA expression of weaned piglets. J. Anim. Nutr. 2018, 30, 4589–4600. [Google Scholar]
- Ashayerizadeh, A.; Dastar, B.; Shargh, M.S.; Mahoonak, A.R.S.; Zerehdaran, S. Effects of feeding fermented rapeseed meal on growth performance, gastrointestinal microflora population, blood metabolites, meat quality, and lipid metabolism in broiler chickens. Livest. Sci. 2018, 216, 183–190. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.T.; Wang, H.L.; Li, D.F.; Piao, X.S.; Lu, W.Q. Optimization of processing conditions for solid-state fermented soybean meal and its effects on growth performance and nutrient digestibility of weanling pigs. Livest. Sci. 2014, 170, 91–99. [Google Scholar] [CrossRef]
- Skrede, G.; Herstad, O.; Sahlstrøm, S.; Holck, A.; Slinde, E.; Skrede, A. Effects of lactic acid fermentation on wheat and barley carbohydrate composition and production performance in the chicken. Anim. Feed Sci. Technol. 2003, 105, 135–148. [Google Scholar] [CrossRef]
- Xin, H.; Wang, M.; Xia, Z.; Yu, B.; He, J.; Yu, J.; Mao, X.; Huang, Z.; Luo, Y.; Luo, J.; et al. Fermented diet liquid feeding improves growth performance and intestinal function of pigs. Animals 2021, 11, 1452. [Google Scholar] [CrossRef]
- Ding, Z.; Chang, K.H.; In-Ho, K. Effects of fermented soybean meal on growth performance, nutrients digestibility, blood profile and fecal microflora in weaning pigs. Korean J. Agric. Sci. 2020, 47, 1–10. [Google Scholar]
- Hu, P.; Wang, L.; Hu, Z.; Jiang, L.; Hu, H.; Rao, Z.; Wu, L.; Tang, Z. Effects of multi-bacteria solid-state fermented diets with different crude fiber levels on growth performance, nutrient digestibility, and microbial flora of finishing pigs. Animals 2021, 11, 3079. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, J.; Sun, Y.; Wang, Y.; He, Z. Prodrug design targeting intestinal PepT1 for improved oral absorption: Design and performance. Curr. Drug Metab. 2013, 14, 675–687. [Google Scholar] [CrossRef]
- Gorboulev, V.; Schuermann, A.; Vallon, V.; Kipp, H.; Jaschke, A.; Klessen, D.; Friedrich, A.; Scherneck, S.; Rieg, T.; Cunard, R.; et al. Na+-d-glucose cotransporter SGLT1 is pivotal for intestinal glucose- absorption and glucose-dependent incretin secretion. Diabetes 2012, 61, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.-Y.; Xu, J.; Tang, S.-Q.; Li, H.-Y.; Jiang, Q.-Y.; Zou, X.-T. Ghrelin and its biological effects on pigs. Peptides 2009, 30, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Salfen, B.E.; Carroll, J.A.; Keisler, D.H.; Strauch, T.A. Effects of exogenous ghrelin on feed intake, weight gain, behavior, and endocrine responses in weanling pigs. J. Anim. Sci. 2004, 82, 1957–1966. [Google Scholar] [CrossRef] [PubMed]
- Lewis, L.D.; Williams, J.A. Regulation of cholecystokinin secretion by food, hormones, and neural pathways in the rat. Am. J. Physiol. 1990, 258 Pt 1, G512–G518. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, J.; Young, R.C.; Smith, G.P. Cholecystokinin decreases food intake in rats. J. Comp. Physiol. Psychol. 1973, 84, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Crawley, J.N.; Beinfeld, M.C. Rapid development of tolerance to the behavioural actions of cholecystokinin. Nature 1983, 302, 703–706. [Google Scholar] [CrossRef]
- Tsunoda, Y.; Yao, H.; Park, J.; Owyang, C. Cholecystokinin synthesizes and secretes leptin in isolated canine gastric chief cells. Biochem. Biophys. Res. Commun. 2003, 310, 681–684. [Google Scholar] [CrossRef]
- Barb, C.R.; Hausman, G.J.; Houseknecht, K.L. Biology of leptin in the pig. Domest. Anim. Endocrin. 2001, 21, 297–317. [Google Scholar] [CrossRef]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and orexin receptors: A family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998, 92, 573–585. [Google Scholar] [CrossRef] [Green Version]
- Saleri, R.; Baratta, M.; Mainardi, G.L.; Renaville, R.; Giustina, A.; Quintavalla, F.; Tamanini, C. IGF-I, IGFBP-2 and-3 but not GH concentrations are different in normal and poor growing piglets. Reprod. Nutr. Dev. 2001, 41, 163–172. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | UFD d | FD d |
|---|---|---|
| Corn | 61.56 | 60.06 |
| Soybean meal | 13.8 | 11.3 |
| Wheat bran | 1 | 0 |
| FFS a | 0 | 7.5 |
| Fish meal | 2 | 2 |
| Extruded soybean | 10 | 10 |
| Soybean oil | 2.5 | 2.5 |
| Soybean protein | 2.5 | 2.5 |
| Sugar | 2 | 2 |
| Acidifier | 0.4 | 0.40 |
| Sodium chloride | 0.38 | 0.38 |
| Calcium carbonate | 0.7 | 0.7 |
| Calcium phosphate | 1.3 | 1.30 |
| Lysine HCl | 0.6 | 0.6 |
| L-Threonine | 0.13 | 0.13 |
| DL-Methionine | 0.1 | 0.1 |
| Tryptophan | 0.03 | 0.03 |
| Premix b | 1 | 1 |
| Nutrition level c | ||
| ME (Mcal/kg) | 3.51 | 3.51 |
| Crude protein% | 18.74 | 18.98 |
| Lysine% | 1.57 | 1.57 |
| Calcium% | 0.74 | 0.74 |
| Phosphorus% | 0.59 | 0.59 |
| Gene Name | Primer Sequence (5′–3′) | Product Length (bp) | Ref |
|---|---|---|---|
| PPOX 1 | For.: TTCCTGGAGACCCCACTCTG | 101 | [23] |
| Rev.: GGCAGCAACAGTAGCAGAAG | |||
| Ghrelin | For.: GCTCCTCATGGCAGACTT | 92 | |
| Rev.: CTGGCTTCTTGGACTCCTT | |||
| Gastrin | For.: TCCTCAGCACTGCGGCGG | 87 | |
| Rev.: ATGGAGGAGGAAGAAGAAGC | [24] | ||
| IGF-1 | For.: GCACATCACATCCTCTTCGC | 164 | |
| Rev.: ACCCTGTGGGCTTGTTGAAA | |||
| IGF-1R | For.: CTGTGGGGGCTCCTGTTTTT | 200 | |
| Rev.: GTGAGCTTGGGAAAGCGGTA | |||
| EAAT3 | For.: GGCACCGCACTCTACGAAGCA | 177 | [25] |
| Rev.: GCCCACGGCACTTAGCACGA | |||
| PepT1 | For.: GGTTTAGGCATCGGAGTAAGAAGT | 156 | [26] |
| Rev.: GGTCAAACAAAGCCCAGAACAT | |||
| SGLT1 | For.: GGCTGGCGAAGTATGGTGT | 153 | |
| Rev.: ACAACCACCCAAATCAGAGC | |||
| YWHAZ | For.: TGATGATAAGAAAGGGATTGTGG | 203 | |
| Rev.: GTTCAGCAATGGCTTCATCA | |||
| GAPDH | For.: GTTCCAGTATGATTCCACCCACGGCA | 147 | |
| Rev.: TGCCAGCCCCAGCATCAAAGGTAGAA |
| Phase | Item | UFD | FD | SEM | p Value |
|---|---|---|---|---|---|
| BW, Kg | Initial | 7.41 | 7.35 | 0.35 | 0.912 |
| d 7 | 8.49 | 8.61 | 0.38 | 0.319 | |
| d 21 | 11.68 | 12.14 | 0.48 | 0.009 | |
| d 1–7 | ADFI 1,g/d | 174 | 188 | 6.10 | 0.002 |
| ADG,g/d | 134 | 161 | 4.04 | 0.017 | |
| FCR | 1.29 | 1.19 | 0.06 | 0.146 | |
| d 8–21 | ADFI,g/d | 387 | 418 | 18.09 | 0.026 |
| ADG,g/d | 262 | 327 | 15.58 | 0.006 | |
| FCR | 1.48 | 1.31 | 0.06 | 0.040 | |
| Overall (d 1–21) | ADFI,g/d | 312 | 338 | 8.16 | 0.028 |
| ADG,g/d | 216 | 245 | 9.55 | <0.001 | |
| FCR | 1.45 | 1.38 | 0.03 | 0.120 |
| Item | UFD | FD | SEM | p Value |
|---|---|---|---|---|
| Dry matter (%) | 83.85 | 84.05 | 0.68 | 0.847 |
| Crude protein (%) | 76.65 | 79.61 | 0.98 | 0.054 |
| Crude fiber (%) | 64.56 | 56.66 | 2.60 | 0.076 |
| Phosphorus (%) | 42.59 | 53.46 | 2.74 | 0.025 |
| Item | Unit | UFD | FD | SEM | p Value |
|---|---|---|---|---|---|
| TP | g/L | 51.56 | 49.77 | 1.55 | 0.443 |
| ALB | g/L | 30.69 | 29.41 | 0.90 | 0.339 |
| GLB | g/L | 20.86 | 20.43 | 1.44 | 0.839 |
| ALB/GLB | 1.49 | 1.50 | 0.12 | 0.949 | |
| Glucose | mmol/L | 6.41 | 6.77 | 0.50 | 0.629 |
| Urea | mmol/L | 5.89 | 5.34 | 0.39 | 0.333 |
| Phosphorus | g/L | 2.27 | 2.31 | 0.07 | 0.716 |
| GGT | U/L | 43.29 | 41.86 | 4.35 | 0.832 |
| AST | U/L | 133.40 | 180.30 | 16.29 | 0.077 |
| ALT | U/L | 48.00 | 50.00 | 4.47 | 0.757 |
| TG | mM | 1.58 | 1.66 | 0.07 | 0.481 |
| TCHO | mmol/L | 1.58 | 1.80 | 0.10 | 0.141 |
| TBA | μmol/L | 19.47 | 31.32 | 4.57 | 0.093 |
| LDL-C | mM | 0.80 | 0.94 | 0.06 | 0.137 |
| HDL-C | mM | 0.60 | 0.66 | 0.05 | 0.366 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Zhang, R.; Lei, H.; Hang, Y.; Xue, H.; Cai, X.; Lu, Y. Supplementation with Fermented Feedstuff Enhances Orexin Expression and Secretion Associated with Increased Feed Intake and Weight Gain in Weaned Pigs. Animals 2022, 12, 1329. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101329
Lu Y, Zhang R, Lei H, Hang Y, Xue H, Cai X, Lu Y. Supplementation with Fermented Feedstuff Enhances Orexin Expression and Secretion Associated with Increased Feed Intake and Weight Gain in Weaned Pigs. Animals. 2022; 12(10):1329. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101329
Chicago/Turabian StyleLu, Yang, Ruiyang Zhang, Hulong Lei, Yiqiong Hang, Huiqin Xue, Xuan Cai, and Yonghong Lu. 2022. "Supplementation with Fermented Feedstuff Enhances Orexin Expression and Secretion Associated with Increased Feed Intake and Weight Gain in Weaned Pigs" Animals 12, no. 10: 1329. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101329