
A 20-SNP Panel as a Tool for Genetic Authentication and Traceability of Pig Breeds

 ,
,  , , , ,
, , , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Breeds, Sampling, and Genotyping
2.2. Data Analysis and SNP Selection
3. Results
3.1. Genotyping
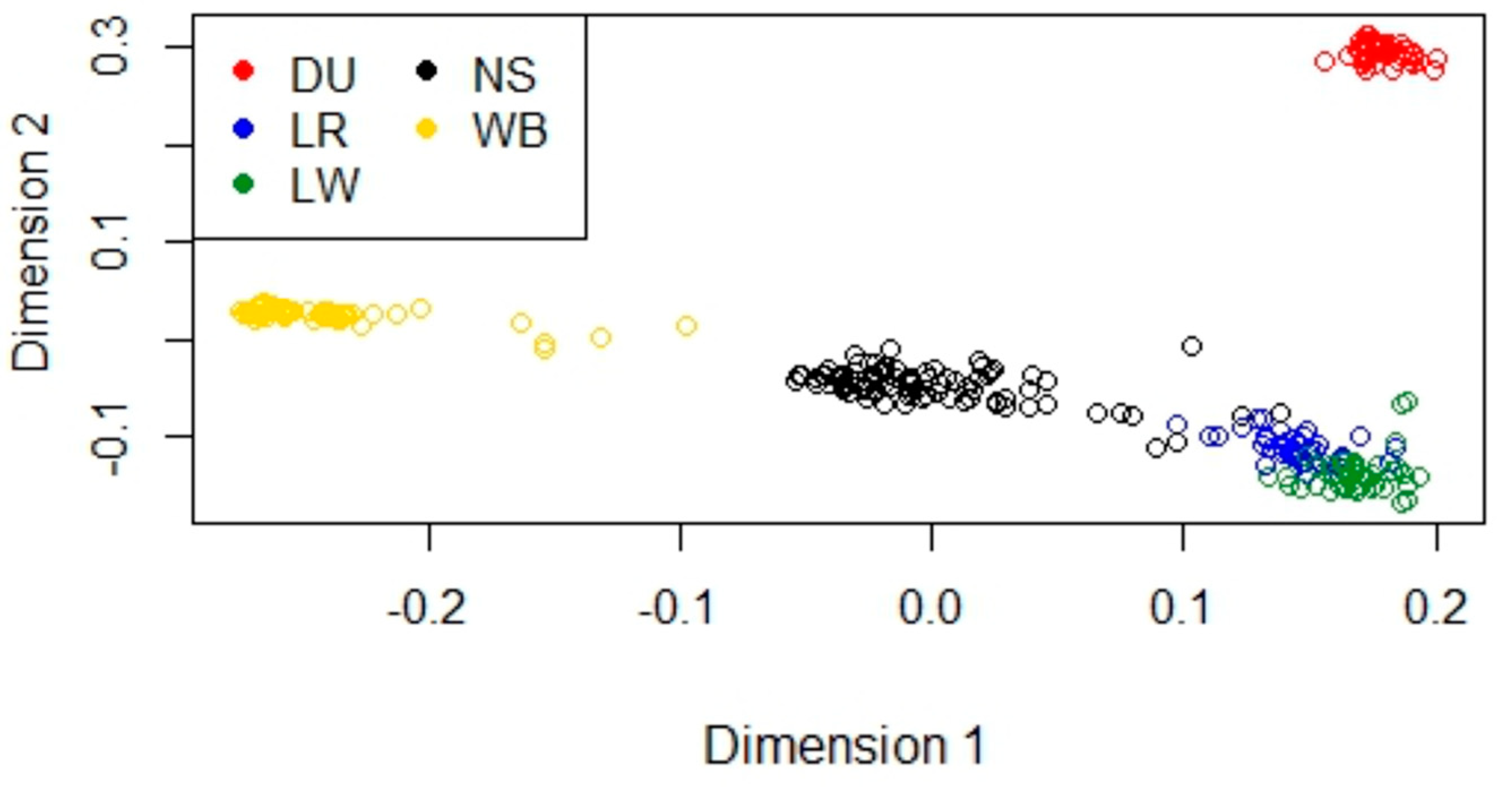
3.2. Multidimensional Scaling
3.3. SNP Selection
3.4. Canonical Discriminant Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Orrù, L.; Catillo, G.; Napolitano, F.; De Matteis, G.; Scatà, M.C.; Signorelli, F.; Moioli, B. Characterization of a SNPs panel for meat traceability in six cattle breeds. Food Control 2009, 20, 856–860. [Google Scholar] [CrossRef]
- Ammendrup, S.; Füssel, A.E. Legislative requirements for the identification and traceability of farm animals within the European Union. OIE Rev. Sci. Technol. 2001, 20, 437–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenstra, J.A. DNA methods for identifying plant and animal species in food. In Food Authenticity and Traceability; Lees, M., Ed.; Woodhead Publishing Ltd.: Cambridge, UK, 2003; pp. 34–53. [Google Scholar]
- Gvozdanović, K.; Kušec, I.D.; Margeta, P.; Salajpal, K.; Džijan, S.; Bošnjak, Z.; Kušec, G. Multiallelic marker system for traceability of Black Slavonian pig meat. Food Control 2020, 109, 106917. [Google Scholar] [CrossRef]
- Oh, J.-D.; Song, K.-D.; Seo, J.-H.; Kim, D.-K.; Kim, S.-H.; Seo, K.-S.; Lim, H.-T.; Lee, J.-B.; Park, H.-C.; Ryu, Y.-C.; et al. Genetic traceability of black pig meats using microsatellite markers. Asian Austral. J. Anim. Sci. 2014, 27, 926–931. [Google Scholar] [CrossRef] [Green Version]
- Gama, L.T.; Martínez, A.M.; Carolino, I.; Landi, V.; Delgado, J.V.; Vicente, A.A.; Vega-Pla, J.L.; Cortés, O.; Sousa, C.O. Genetic structure, relationships and admixture with wild relatives in native pig breeds from Iberia and its islands. Genet. Sel. Evol. 2013, 45, 18. [Google Scholar] [CrossRef] [Green Version]
- Ramos, A.M.; Megens, H.J.; Crooijmans, R.P.M.A.; Schook, L.B.; Groenen, M.A.M. Identification of high utility SNPs for population assignment and traceability purposes in the pig using high-throughput sequencing: Identification of high utility SNPs. Anim. Genet. 2011, 42, 613–620. [Google Scholar] [CrossRef]
- Alves, E.; Fernández, A.I.; Fernández-Rodríguez, A.; Pérez-Montarelo, D.; Benitez, R.; Óvilo, C.; Rodríguez, C.; Silió, L. Identification of mitochondrial markers for genetic traceability of European wild boars and Iberian and Duroc pigs. Animal 2009, 3, 1216–1223. [Google Scholar] [CrossRef] [Green Version]
- Ojeda, A.; Ramos-Onsins, S.E.; Marletta, D.; Huang, L.S.; Folch, J.M.; Pérez-Enciso, M. Evolutionary study of a potential selection target region in the pig. Heredity 2011, 106, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Misra, A. SNP genotyping: Technologies and biomedical applications. Annu. Rev. Biomed. Eng. 2007, 9, 289–320. [Google Scholar] [CrossRef]
- Negrini, R.; Nicoloso, L.; Crepaldi, P.; Milanesi, E.; Marino, R.; Perini, D.; Pariset, L.; Dunner, S.; Leveziel, H.; Williams, J.L.; et al. Traceability of four European Protected Geographic Indication (PGI) beef products using Single Nucleotide Polymorphisms (SNP) and Bayesian statistics. Meat Sci. 2008, 80, 1212–1217. [Google Scholar] [CrossRef]
- Ripoli, M.V.; Wei, S.; Rogberg-Muñoz, A.; Guo, B.L.; Goszczynski, D.E.; Fernandez, M.E.; Mellucci, L.; Lirón, J.P.; Villarreal, E.; Wei, Y.M.; et al. Evaluation of six nucleotide polymorphisms for bovine traceability in the context of the Argentine-Chinese beef trade. BAG J. Basic Appl. Genet. 2013, 24, 31–45. [Google Scholar]
- Bozzi, R.; García-Casco, J.; Núñez, Y.; Ribani, A.; Franci, O.; García, F.; Škrlep, M.; Schiavo, G.; Bovo, S. Genomic diversity, linkage disequilibrium and selection signatures in European local pig breeds assessed with a high-density SNP chip. Sci. Rep. 2019, 9, 13546. [Google Scholar]
- Wilkinson, S.; Wiener, P.; Archibald, A.L.; Law, A.; Schnabel, R.D.; McKay, S.D.; Taylor, J.F.; Ogden, R. Evaluation of approaches for identifying population informative markers from high density SNP Chips. BMC Genet. 2011, 12, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontanesi, L. Genetic authentication and traceability of food products of animal origin: New developments and perspectives. Ital. J. Anim. Sci. 2010, 8, 9–18. [Google Scholar] [CrossRef]
- Schiavo, G.; Bovo, S.; Tinarelli, S.; Gallo, M.; Dall’Olio, S.; Fontanesi, L. Genome-wide association analyses for coat colour patterns in the autochthonous Nero Siciliano pig breed. Livest. Sci. 2020, 236, 104015. [Google Scholar] [CrossRef]
- Chessa, S.; Bordonaro, S.; Moretti, R.; Criscione, A.; Marletta, D.; Castiglioni, B. Genomic analysis for the valorization of Nero Siciliano swine breed. Ital. J. Anim. Sci. 2013, 12, 68. [Google Scholar]
- Pizzi, F.; Turri, F.; Gliozzi, T.M.; Gandini, G. Use of epididymal semen cryoconservation in farm animal gene banking: Field applications in Brianzola sheep and Nero Siciliano pig breeds. Ital. J. Anim. Sci. 2013, 12, 83. [Google Scholar]
- Guastella, A.M.; Criscione, A.; Marletta, D.; Zuccaro, A.; Chies, L.; Bordonaro, S. Molecular characterization and genetic structure of the Nero Siciliano pig breed. Genet. Mol. Biol. 2010, 33, 650–656. [Google Scholar] [CrossRef] [Green Version]
- Chessa, S.; Stella, A.; Raschetti, M.; Passero, A.; Crepaldi, P.; Nicoloso, L.; Castiglioni, B.; Pagnacco, G. Genomic analysis for the valorization of an Italian local swine breed. Ital. J. Anim. Sci. 2011, 10, 131. [Google Scholar]
- Goedbloed, D.J.; Megens, H.J.; Van Hooft, P.; Herrero-Medrano, J.M.; Lutz, W.; Alexandri, P.; Crooijmans, R.P.M.A.; Groenen, M.; Van Wieren, S.E.; Ydenberg, R.C.; et al. Genome-wide single nucleotide polymorphism analysis reveals recent genetic introgression from domestic pigs into Northwest European wild boar populations. Mol. Ecol. 2013, 22, 856–866. [Google Scholar] [CrossRef]
- Nielsen, E.E.; Bach, L.A.; Kotlicki, P. Hybridlab (version 1.0): A program for generating simulated hybrids from population samples. Mol. Ecol. Notes 2006, 6, 971–973. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiggans, G.R.; Sonstegard, T.S.; Van Raden, P.M.; Matukumalli, L.K.; Schnabel, R.D.; Taylor, J.F.; Schenkel, F.S.; Van Tassell, C.P. Selection of single-nucleotide polymorphisms and quality of genotypes used in genomic evaluation of dairy cattle in the United States and Canada. J. Dairy Sci. 2009, 92, 3431–3436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. 2014. Available online: http://www.R-project.org/ (accessed on 22 March 2022).
- GenABEL Project Developers. GenABEL: Genome-Wide SNP Association Analysis. R Package Version 1.8-0. 2013. Available online: http://CRAN.R-project.org/package=GenABEL (accessed on 22 March 2022).
- Bowcock, A.M.; Ruiz-Linares, A.; Tomfohrde, J.; Minch, E.; Kidd, J.R.; Cavalli-Sforza, L.L. High resolution of human evolutionary trees with polymorphic microsatellites. Nature 1994, 368, 455–457. [Google Scholar] [CrossRef]
- Paschou, P.; Ziv, E.; Burchard, E.G.; Choudhry, S.; Rodriguez-Cintron, W.; Mahoney, M.W.; Drineas, P. PCA-Correlated SNPs for structure identification in worldwide human populations. PLoS Genet. 2007, 3, 1672–1686. [Google Scholar] [CrossRef] [Green Version]
- Shriver, M.D.; Smith, M.W.; Jin, L.; Marcini, A.; Akey, J.M.; Deka, R.; Ferrell, R.E. Ethnic-affiliation estimation by use of population-specific DNA markers. Am. J. Hum. Genet. 1997, 60, 957–964. [Google Scholar]
- Wilkinson, S.; Archibald, A.L.; Haley, C.S.; Megens, H.J.; Crooijmans, R.P.; Groenen, M.A.; Wiener, P.; Ogden, R. Development of a genetic tool for product regulation in the diverse British pig breed market. BMC Genom. 2012, 13, 580. [Google Scholar] [CrossRef] [Green Version]
- Dimauro, C.; Cellesi, M.; Steri, R.; Gaspa, G.; Sorbolini, S.; Stella, A.; Macciotta, N.P.P. Use of the canonical discriminant analysis to select SNP markers for bovine breed assignment and traceability purposes. Anim. Genet. 2013, 44, 377–382. [Google Scholar] [CrossRef]
- Friendly, M.; Fox, J. Candisc: Visualizing Generalized Canonical Discriminant and Canonical Correlation Analysis. R package Version 0.6-7. 2015. Available online: http://CRAN.R-project.org/package=candisc (accessed on 22 March 2022).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Ramos, A.M.; Crooijmans, R.P.M.A.; Affara, N.A.; Amaral, A.J.; Archibald, A.L.; Beever, J.E.; Bendixen, C.; Churcher, C.; Clark, R.; Dehais, P.; et al. Design of a high density SNP genotyping assay in the pig using SNPs identified and characterized by next generation sequencing technology. PLoS ONE 2009, 4, e6524. [Google Scholar] [CrossRef] [Green Version]
- Geibel, J.; Reimer, C.; Weigend, S.; Weigend, A.; Pook, T.; Simianer, H. How array design creates SNP ascertainment bias. PLoS ONE 2021, 16, e0245178. [Google Scholar]
- Muñoz, M.; Bozzi, R.; García, F.; Núñez, Y.; Geraci, C.; Crovetti, A.; García-Casco, J.; Alves, E.; Škrlep, M.; Charneca, R.; et al. Diversity across major and candidate genes in European local pig breeds. PLoS ONE 2018, 13, e0207475. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| SNP | SNP ID | Chromosome | Position | Breed 1 | Δ with NS |
|---|---|---|---|---|---|
| MARC0027620 | rs81222690 | 1 | 17,004,757 | DU | 0.621 |
| INRA0002279 | rs326314161 | 1 | 57,648,581 | LW | 0.546 |
| ASGA0004735 | rs80788426 | 1 | 159,034,424 | LW | 0.783 |
| ASGA0007653 | rs81352517 | 1 | 300,830,758 | LW | 0.506 |
| H3GA0006564 2 | rs81357620 | 2 | 42,213,270 | NS | 0.701 |
| CASI0009067 | rs329578899 | 2 | 108,053,445 | WB | 1.000 |
| ASGA0021239 | rs81001550 | 4 | 105,898,886 | DU | 0.939 |
| ALGA0027544 | rs80958781 | 4 | 113,430,367 | LW | 0.731 |
| M1GA0006536 | rs80935048 | 4 | 133,828,395 | LR | 0.536 |
| H3GA0016973 | rs81385751 | 5 | 85,729,923 | LW | 0.723 |
| MARC0038980 | rs81232179 | 8 | 51,070,662 | LR | 0.761 |
| INRA0029816 | rs327195280 | 8 | 51,235,665 | LR | 0.763 |
| ALGA0047912 | rs81400622 | 8 | 57,588,232 | LW | 0.743 |
| MARC0042228 | rs81234311 | 8 | 139,592,943 | WB | 0.491 |
| ALGA0065765 2 | rs81433050 | 12 | 27,112,308 | NS | 0.758 |
| ASGA0069722 | rs81453203 | 15 | 63,655,984 | DU | 0.932 |
| ALGA0107321 | rs81335037 | 15 | 141,212,829 | DU | 0.779 |
| ASGA0095426 | rs81314826 | 15 | 149,495,100 | WB | 0.953 |
| INRA0052808 | rs342665431 | 17 | 17,548,564 | LR | 0.607 |
| DBMA0000205 | rs45432506 | 17 | 20,690,224 | LR | 0.788 |
| Breed 1 | 12 SNP | 20 SNP | ||||||
|---|---|---|---|---|---|---|---|---|
| PTP 2 | PVP 2 | HTP 2 | HVP 2 | PTP 2 | PVP 2 | HTP 2 | HVP 2 | |
| NS | 92/92 (100%) | 40/40 (100%) | 89/92 (96.7%) | 40/40 (100%) | 92/92 (100%) | 40/40 (100%) | 87/92 (94.6%) | 40/40 (100%) |
| LW | 34/36 (94.4%) | 36/40 (90%) | 33/36 (91.7%) | 33/40 (82.5%) | 36/36 (100%) | 40/40 (100%) | 36/36 (100%) | 40/40 (100%) |
| LR | 34/36 (94.4%) | 38/40 (95%) | 32/36 (88.9%) | 38/40 (95%) | 36/36 (100%) | 40/40 (100%) | 34/36 (94.4%) | 40/40 (100%) |
| DU | 39/40 (97.5%) | 40/40 (100%) | 37/40 (92.5%) | 36/40 (90%) | 40/40 (100%) | 40/40 (100%) | 39/40 (97.5%) | 40/40 (100%) |
| WB | 71/71 (100%) | 40/40 (100%) | 71/71 (100%) | 39/40 (97.5%) | 71/71 (100%) | 40/40 (100%) | 71/71 (100%) | 40/40 (100%) |
| HY | - | - | 143/200 (71.5%) | 75/100 (75%) | - | - | 171/200 (85.5%) | 84/100 (84%) |
| Purebred individuals | * | * | 262/275 (95.3%) | 186/200 (93%) | * | * | 267/275 (97.1%) | 200/200 (100%) |
| Entire population | 270/275 (98.2%) | 194/200 (97%) | 405/475 (85.3%) | 261/300 (87%) | 275/275 (100%) | 200/200 (100%) | 438/475 (92.2%) | 284/300 (94.7%) |
| Breed 1 | CACDA | Wrongly Assigned Breed | Mean Assignation Probability | Lowest Assignation Probability | CAT65 | Wrongly Assigned Breed |
|---|---|---|---|---|---|---|
| NS | 40/40 | 0.981 | 0.771 | 40/40 | ||
| DU | 40/40 | 0.980 | 0.757 | 40/40 | ||
| LR | 40/40 | 0.957 | 0.568 | 39/40 | HY | |
| LW | 40/40 | 0.979 | 0.618 | 39/40 | HY | |
| WB | 40/40 | 0.999 | 0.982 | 40/40 | ||
| NS × DU | 9/10 | DU | 0.842 | 0.390 | 10/10 | |
| NS × LR | 9/10 | NS | 0.786 | 0.303 | 10/10 | |
| NS × LW | 10/10 | 0.775 | 0.566 | 10/10 | ||
| NS × WB | 6/10 | 2 NS and 2 WB | 0.652 | 0.360 | 10/10 | |
| DU × LR | 10/10 | 0.948 | 0.831 | 10/10 | ||
| DU × LW | 10/10 | 0.827 | 0.534 | 10/10 | ||
| DU × WB | 10/10 | 0.924 | 0.865 | 10/10 | ||
| LR × LW | 4/10 | 3 LR and 3 LW | 0.493 | 0.054 | 7/10 | 1 LR and 2 LW |
| LR × WB | 7/10 | 1 LR and 2 WB | 0.600 | 0.096 | 8/10 | 1 LR and 1 WB |
| LW × WB | 9/10 | WB | 0.818 | 0.366 | 10/10 | |
| Whole population | 284/300 (94.7%) | 293/300 (97.7%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moretti, R.; Criscione, A.; Turri, F.; Bordonaro, S.; Marletta, D.; Castiglioni, B.; Chessa, S. A 20-SNP Panel as a Tool for Genetic Authentication and Traceability of Pig Breeds. Animals 2022, 12, 1335. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12111335
Moretti R, Criscione A, Turri F, Bordonaro S, Marletta D, Castiglioni B, Chessa S. A 20-SNP Panel as a Tool for Genetic Authentication and Traceability of Pig Breeds. Animals. 2022; 12(11):1335. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12111335
Chicago/Turabian StyleMoretti, Riccardo, Andrea Criscione, Federica Turri, Salvatore Bordonaro, Donata Marletta, Bianca Castiglioni, and Stefania Chessa. 2022. "A 20-SNP Panel as a Tool for Genetic Authentication and Traceability of Pig Breeds" Animals 12, no. 11: 1335. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12111335