
Diachroneity Rules the Mid-Latitudes: A Test Case Using Late Neogene Planktic Foraminifera across the Western Pacific

 ,
,
Abstract
:1. Introduction

2. Materials and Methods
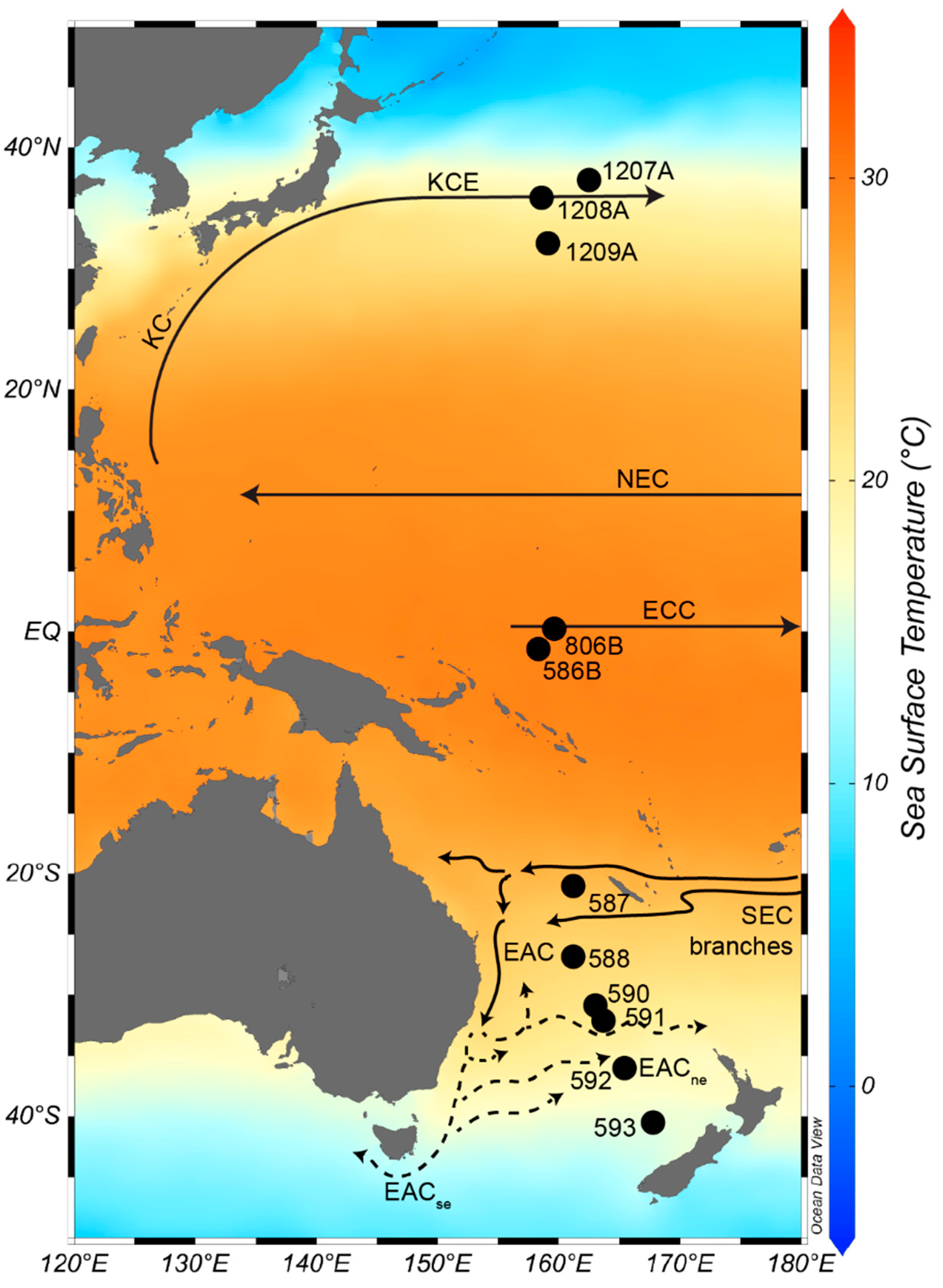
2.1. ODP and DSDP Sites Utilized
2.2. Foraminiferal Occurrence Data, Taxonomy, and Age Models
3. Results
3.1. Late Neogene Western Pacific Planktic Foraminiferal Biostratigraphy

3.1.1. Tropical Holes 806B and 586B
3.1.2. Warm Subtropical Sites 587 and 588
3.1.3. Cool Subtropical Sites 590, 591, and 592
3.1.4. Temperate Site 593
3.2. Western Pacific Plankton Biogeographic Patterns
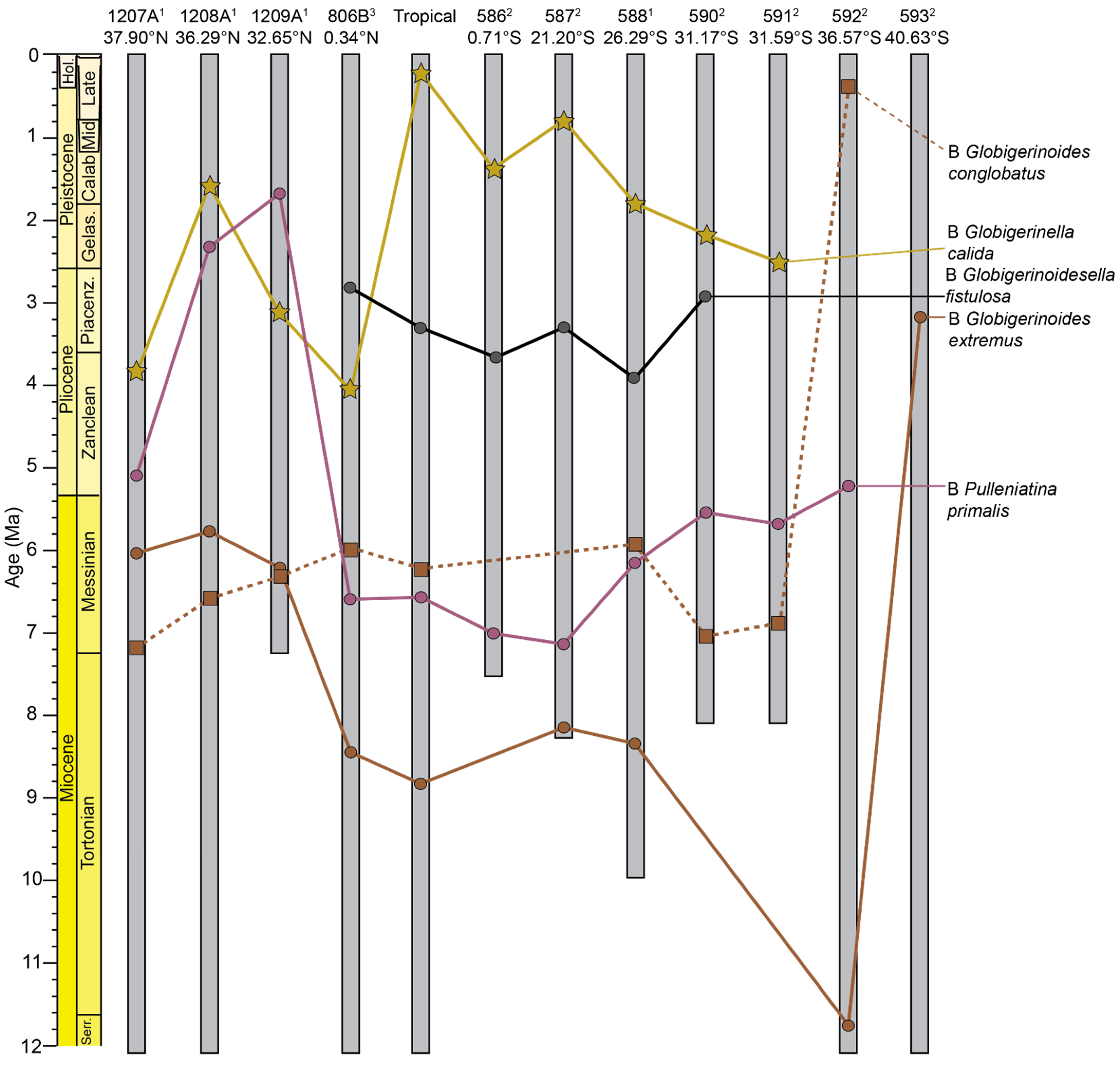
3.2.1. Globigerinella Biogeographic Patterns
3.2.2. Globigerinoides Biogeographic Patterns


3.2.3. Globigerinoidesella Biogeographic Patterns
3.2.4. Pulleniatina Biogeographic Patterns
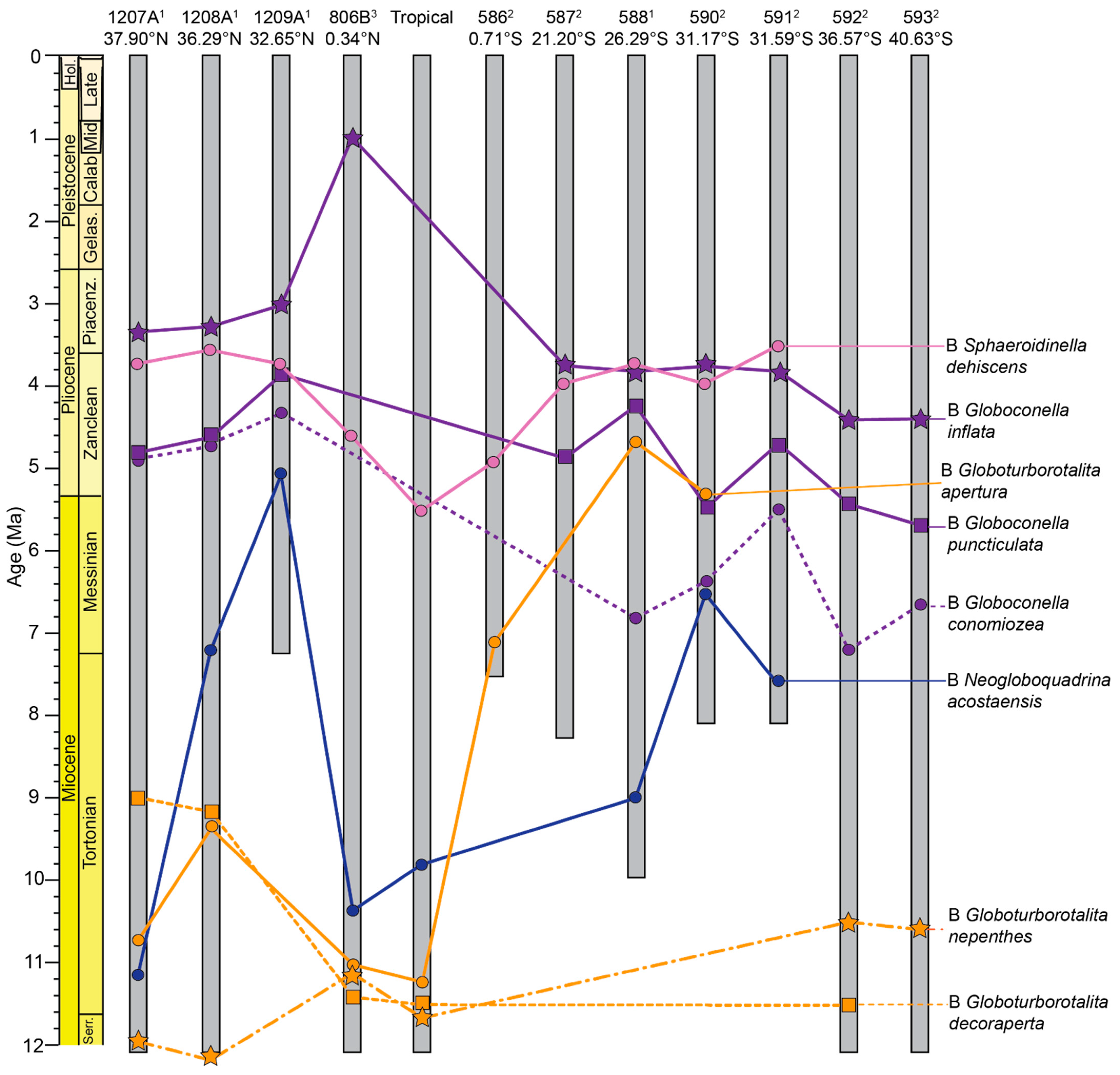
3.2.5. Globoquadrina Biogeographic Patterns
3.2.6. Sphaeroidinella Biogeographic Patterns
3.2.7. Sphaeroidinellopsis Biogeographic Patterns
3.2.8. Globoconella Biogeographic Patterns

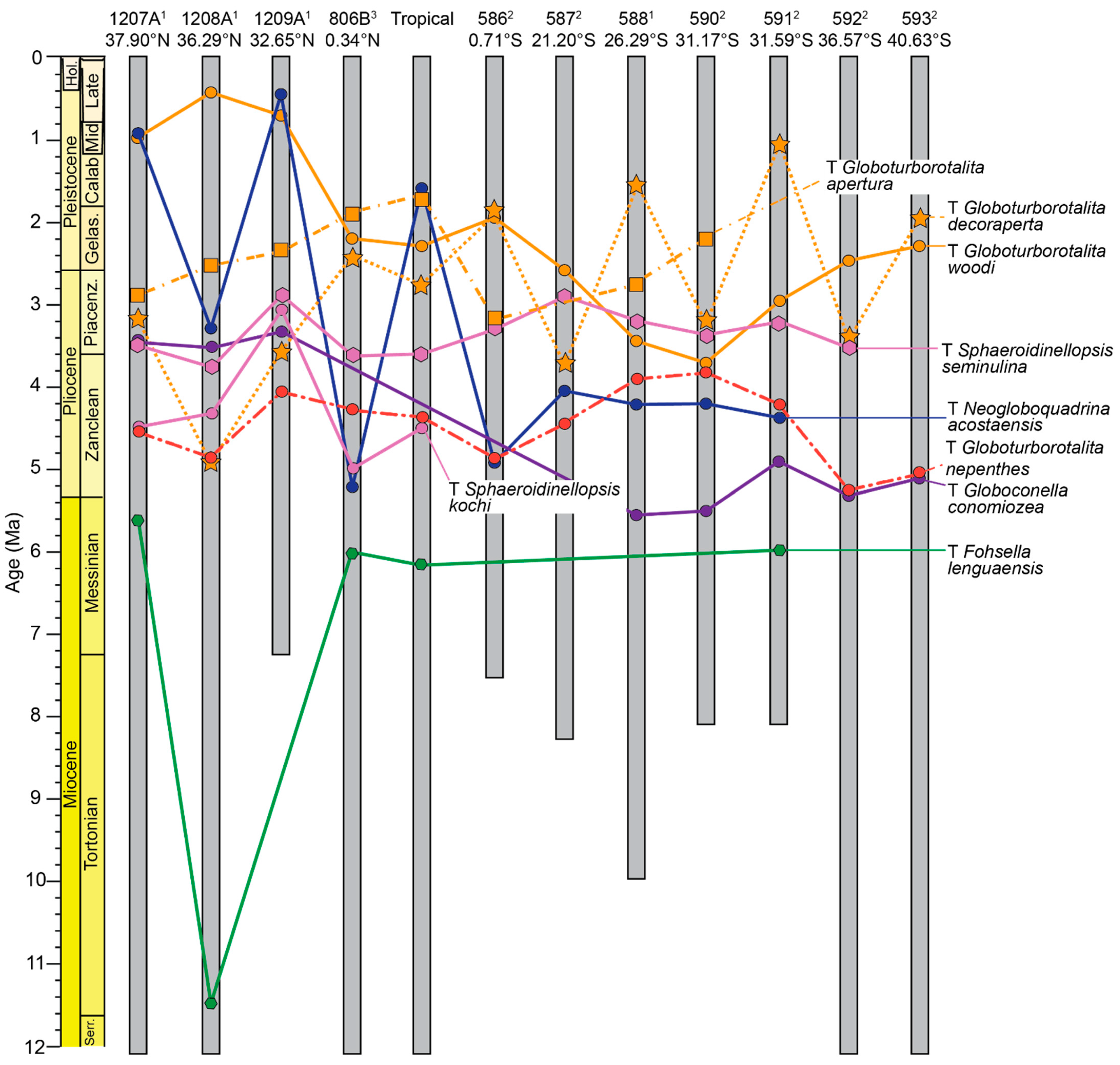
3.2.9. Globoturborotalita Biogeographic Patterns

3.2.10. Neogloboquadrina Biogeographic Patterns
3.2.11. Fohsella Biogeographic Patterns
3.2.12. Hirsutella Biogeographic Patterns
3.2.13. Truncorotalia Biogeographic Patterns

3.2.14. Globorotalia Biogeographic Patterns
3.2.15. Menardella Biogeographic Patterns

3.2.16. Dentoglobigerina Biogeographic Patterns
4. Discussion
4.1. Sources of Error
4.2. Biostratigraphy across the Western Pacific
4.3. Diachroneity of Planktic Foraminiferal Datums
4.3.1. Diachroneity between the Western Equatorial Pacific and Equatorial Atlantic Oceans
4.3.2. Diachroneity within the Northwest and Southwest Pacific
4.3.3. Diachroneity between the Northwest and Southwest Pacific
4.3.4. Diachroneity between the Western Equatorial and Eastern Equatorial Pacific, and the Eastern Equatorial Indian Ocean
5. Major Biogeographic Patterns and Their Oceanographic Drivers
6. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leckie, R.M. Seeking a better life in the plankton. Proc. Natl. Acad. Sci. USA 2009, 106, 14183–14184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraass, A.J.; Kelly, D.C.; Peters, S.E. Macroevolutionary history of the planktic foraminifera. Annu. Rev. Earth Planet. Sci. 2015, 43, 139–166. [Google Scholar] [CrossRef] [Green Version]
- Gradstein, F.; Waskowska, A.; Glinskikh, L. The First 40 Million Years of Planktonic Foraminifera. Geosciences 2021, 11, 85. [Google Scholar] [CrossRef]
- Lam, A.R.; Bauer, J.E.; Fraass, S.; Sheffield, S.; Limbeck, M.R.; Borden, R.M.; Bryant, R. Time Scavengers: An Educational Website to Communicate Climate Change and Evolutionary Theory to the Public through Blogs, Web Pages, and Social Media Platforms. J. STEM Outreach 2019, 7, 1–8. [Google Scholar] [CrossRef]
- Hsiang, A.Y.; Brombacher, A.; Rillo, M.C.; Mleneck-Vautravers, M.J.; Conn, S.; Lordsmith, S.; Hull, P.M. Endless Forams: >34,000 modern planktonic foraminiferal images for taxonomic training and automated species recognition using convolutional neural networks. Paleoceanogr. Paleoclimatol. 2019, 34, 1157–1177. [Google Scholar] [CrossRef] [Green Version]
- Pearson, P.N.; Shackleton, N.J.; Hall, M.A. Stable isotopic evidence for the sympatric divergence of Globigerinoides trilobus and Orbulina universa (planktonic foraminifera). J. Geol. Soc. 1997, 154, 295–302. [Google Scholar] [CrossRef]
- Darling, K.F.; Wade, C.M.; Stewart, I.A.; Kroon, D.; Dingle, R.; Brown, A.J.L. Molecular evidence for genetic mixing of Arctic and Antarctic subpolar populations of planktonic foraminifers. Nature 2000, 405, 43–47. [Google Scholar] [CrossRef]
- Norris, R.D.; de Vargas, C. Evolution all at sea. Nature 2000, 405, 23–24. [Google Scholar] [CrossRef]
- Brombacher, A.; Wilson, P.A.; Bailey, I.; Ezard, T.H. The breakdown of static and evolutionary allometries during climatic upheaval. Am. Nat. 2017, 190, 350–362. [Google Scholar] [CrossRef] [Green Version]
- Woodhouse, A.; Jackson, S.; Jamieson, R.; Newton, R.; Sexton, P.; Aze, T. Adaptive Ecological Niche Migration does not Negate Extinction Susceptibility. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Boscolo-Galazzo, F.; Jones, A.; Dunkley Jones, T.; Crichton, K.A.; Wade, B.S.; Pearson, P.N. Late Neogene evolution of modern deep-dwelling plankton. Biogeosciences 2022, 19, 743–762. [Google Scholar] [CrossRef]
- Wade, B.S.; Pearson, P.N.; Berggren, W.A.; Pälike, H. Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale. Earth-Sci. Rev. 2011, 104, 111–142. [Google Scholar] [CrossRef] [Green Version]
- King, D.J.; Wade, B.S.; Liska, R.D.; Miller, C.G. A review of the importance of the Caribbean region in Oligo-Miocene low latitude planktonic foraminiferal biostratigraphy and the implications for modern biogeochronological schemes. Earth-Sci. Rev. 2020, 202, 102968. [Google Scholar] [CrossRef]
- Kennett, J.P. Middle and late Cenozoic planktonic foraminiferal biostratigraphy of the south-west Pacific- DSDP Leg 21. In Initial Reports of the Deep Sea Drilling Project; Burns, R.E., Andrews, J.E., van der Lingen, G.J., Churkin, M., Jr., Galehouse, J.S., Packham, G.H., Davies, T.A., Kennett, J.P., Dumitrica, P., Edwards, A.R., et al., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1973; Volume 21, pp. 575–639. [Google Scholar]
- Srinivasan, M.S.; Kennett, J.P. Neogene planktonic foraminiferal biostratigraphy and evolution: Equatorial to subantarctic, South Pacific. Mar. Micropaleontol. 1981, 6, 499–533. [Google Scholar] [CrossRef]
- Srinivasan, M.S.; Kennett, J.P. A Review of Neogene Planktonic Foraminiferal Biostratigraphy Applications in the Equatorial and South Pacific; SEPM Special Publications: Tulsa, AK, USA, 1981; Volume 32, pp. 395–432. [Google Scholar]
- Kennett, J.P.; Srinivasan, M.S. Neogene Planktonic Foraminifera a Phylogenetic Atlas; Hutchinson Ross Publishing Co.: Hutchinson, KS, USA, 1983. [Google Scholar]
- Jenkins, D.G.; Srinivasan, M.S. Cenozoic Planktonic Foraminifers from the Equator to the sub-Antarctic of the Southwest Pacific. In Initial Reports of the Deep Sea Drilling Project; Kennett, J.P., von der Borch, C.C., Baker, P.A., Barton, C.E., Boersma, A., Caulet, J.P., Dudley, W.C., Jr., Gardner, J.V., Jenkins, D.G., Lohman, W.H., et al., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1986; Volume 90, pp. 795–834. [Google Scholar]
- Cifelli, R.; Scott, G. Stratigraphic record of the Neogene globorotalid radiation (Planktonic Foraminiferida). Smithson. Contrib. Paleobiol. 1986, 58. [Google Scholar] [CrossRef]
- Scott, G.H. Planktonic foraminiferal biostratigraphy (Altonian-Tongaporutuan Stages, Miocene) at DSDP Site 593, Challenger Plateau, Tasman Sea. N. Z. J. Geol. Geophys. 1992, 35, 501–513. [Google Scholar] [CrossRef]
- Keller, G. Planktonic foraminiferal biostratigraphy and paleoceanography of the Japan trench, Leg 57, Deep Sea Drilling Project. In Initial Reports of the Deep Sea Drilling Project; von Huene, R., Nasu, N., Arthur, M.A., Barron, J.A., Bell, G.D., Cadet, J.-P., Carson, B., Fujioka, K., Honza, E., Keller, G., et al., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1980; Volume 57, pp. 809–833. [Google Scholar]
- Weaver, P.P.E.; Clement, B.M. Magnetobiostratigraphy of planktonic foraminiferal datums: Deep Sea Drilling Project Leg 94, North Atlantic. In Initial Reports of the Deep Sea Drilling Project; Ruddiman, W.F., Kidd, R.B., Baldauf, J.G., Clement, B.M., Dolan, J.F., Eggers, M.R., Hill, M.R., Keigwin, L.D., Jr., Mitchell, M., Philipps, I., et al., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1986; Volume 94, pp. 815–829. [Google Scholar]
- Fraass, A.J.; Leckie, R.M.; Lowery, C.M.; DeConto, R. Precision in Biostratigraphy: Evidence for a temporary flow reversal in the Central American Seaway during or after the Oligocene-Miocene transition. J. Foraminifer. Res. 2019, 49, 357–366. [Google Scholar] [CrossRef]
- Scott, G.H. Divergences and phyletic transformations in the history of the Globorotalia inflata lineage. Paleobiology 1983, 9, 422–426. [Google Scholar] [CrossRef]
- Hodell, D.A.; Kennett, J.P. Late Miocene–early Pliocene stratigraphy and paleoceanography of the South Atlantic and southwest Pacific Oceans: A synthesis. Paleoceanography 1986, 1, 285–311. [Google Scholar] [CrossRef]
- Dowsett, H.J. Diachrony of late Neogene microfossils in the southwest Pacific Ocean: Application of the graphic correlation method. Paleoceanography 1988, 3, 209–222. [Google Scholar] [CrossRef]
- Dowsett, H.J. Improved dating of the Pliocene of the eastern South Atlantic using graphic correlation: Implications for paleobiogeography and paleoceanography. Micropaleontology 1989, 35, 279–292. [Google Scholar] [CrossRef]
- Srinivasan, M.S.; Sinha, D.K. Improved correlation of the late Neogene planktonic foraminiferal datums in the equatorial to cool subtropical DSDP sites, southwest Pacific: Application of the graphic correlation method. Mem. Geol. Soc. India 1991, 20, 55–93. [Google Scholar]
- Jenkins, D.G. The paleogeography, evolution and extinction of Late Miocene-Pleistocene planktonic foraminifera from the southwest Pacific. Centen. Jpn. Micropaleontol. 1992, 27, 1–35. [Google Scholar]
- Spencer-Cervato, C.; Thierstein, H.R.; Lazarus, D.B.; Beckmann, J.P. How synchronous are Neogene marine plankton events? Paleoceanography 1994, 9, 739–763. [Google Scholar] [CrossRef] [Green Version]
- Lam, A.R.; Leckie, R.M. Subtropical to temperate late Neogene to Quaternary planktic foraminiferal biostratigraphy across the Kuroshio Current Extension, Shatsky Rise, northwest Pacific Ocean. PLoS ONE 2020, 15, e0234351. [Google Scholar] [CrossRef]
- Lam, A.R.; Stigall, A.L. Pathways and mechanisms of Late Ordovician (Katian) faunal migrations of Laurentia and Baltica. Est. J. Earth Sci. 2015, 64, 62–67. [Google Scholar] [CrossRef]
- Lam, A.R.; Stigall, A.L.; Matzke, N.J. Dispersal in the Ordovician: Speciation patterns and paleobiogeographic analyses of brachiopods and trilobites. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 489, 147–165. [Google Scholar] [CrossRef] [Green Version]
- Lam, A.R.; Leckie, R.M.; Patterson, M.O. Illuminating the Past to See the Future of Western Boundary Currents. Oceanography 2020, 33, 65–67. [Google Scholar] [CrossRef]
- Zheng, H.; Sun, X.; Wang, P.; Chen, W.; Yue, J. Mesozoic tectonic evolution of the Proto-South China Sea: A perspective from radiolarian paleobiogeography. J. Asian Earth Sci. 2019, 179, 37–55. [Google Scholar] [CrossRef]
- Bauer, J.E. Paleobiogeography, paleoecology, diversity, and speciation patterns in the Eublastoidea (Blastozoa: Echinodermata). Paleobiology 2021, 47, 221–235. [Google Scholar] [CrossRef]
- Lam, A.R.; Sheffield, S.L.; Matzke, N.J. Estimating dispersal and evolutionary dynamics in diploporan blastozoans (Echinodermata) across the great Ordovician biodiversification event. Paleobiology 2021, 47, 198–220. [Google Scholar] [CrossRef]
- Petrizzo, M.R.; Huber, B.T.; Falzoni, F.; MacLeod, K.G. Changes in biogeographic distribution patterns of southern mid-to high latitude planktonic foraminifera during the Late Cretaceous hot to cool greenhouse climate transition. Cretac. Res. 2020, 115, 104547. [Google Scholar] [CrossRef]
- Brombacher, A.; Wilson, P.A.; Bailey, I.; Ezard, T.H. The Dynamics of Diachronous Extinction Associated with Climatic Deterioration near the Neogene/Quaternary Boundary. Paleoceanogr. Paleoclimatol. 2021, 36, e2020PA004205. [Google Scholar] [CrossRef]
- Huber, B.T.; Tur, N.A.; Self-Trail, J.; MacLeod, K.G. Calcareous plankton biostratigraphic fidelity and species richness during the last 10 my of the Cretaceous at Blake Plateau, subtropical North Atlantic. Cretac. Res. 2022, 131, 105095. [Google Scholar] [CrossRef]
- Sheffield, S.L.; Lam, A.R.; Philips, S.F.; Deline, B. Morphological Dynamics and Response Following the Dispersal of Ordovician-Silurian Diploporan Echinoderms to Laurentia; University of Michigan: Ann Arbor, MI, USA, 2022; in press. [Google Scholar]
- Lam, A.R.; Leckie, R.M. Late Neogene and Quaternary diversity and taxonomy of subtropical to temperate planktic foraminifera across the Kuroshio Current Extension, northwest Pacific Ocean. Micropaleontology 2020, 66, 177–268. [Google Scholar] [CrossRef]
- Chaisson, W.P.; Leckie, R.M. High-resolution Neogene planktonic foraminifer biostratigraphy of Site 806, Ontong Java Plateau (western equatorial Pacific). In Proceedings of the Ocean Drilling Program Scientific Results; Berger, W.H., Kroenke, L.W., Janecek, T.R., Backman, J., Bassinot, F., Corfield, R.M., Delaney, M.L., Hagen, R., Jansen, E., Krissek, L.A., et al., Eds.; Ocean Drilling Program: College Station, TX, USA, 1993; Volume 130, pp. 137–178. [Google Scholar]
- Martini, E. Standard Tertiary and Quaternary calcareous nannoplankton zonation. Proc. II Planktonic Conf. Rome Italy 1970 Tecnoscienz 1971, 2, 739–785. [Google Scholar]
- Backman, J.; Raffi, I.; Rio, D.; Fornaciari, E.; Pälike, H. Biozonation and biochronology of Miocene through Pleistocene calcareous nannofossils from low and middle latitudes. Newsl. Stratigr. 2012, 45, 221–244. [Google Scholar] [CrossRef]
- Nelson, C.S.; Cooke, P.J. History of oceanic front development in the New Zealand sector of the Southern Ocean during the Cenozoic—A synthesis. N. Z. J. Geol. Geophys. 2001, 44, 535–553. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, S.J.; Kitamura, A.; Iryu, Y.; Itaki, T.; Koizumi, I.; Hoiles, P.W. The Pliocene to recent history of the Kuroshio and Tsushima Currents: A multi-proxy approach. Prog. Earth Planet. Sci. 2015, 2, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Lam, A.R.; MacLeod, K.G.; Schilling, S.H.; Leckie, R.M.; Fraass, A.J.; Patterson, M.O.; Venti, N.L. Pliocene to earliest Pleistocene (5–2.5 Ma) Reconstruction of the Kuroshio Current Extension Reveals a Dynamic Current. Paleoceanogr. Paleoclimatol. 2021, 36, e2021PA004318. [Google Scholar] [CrossRef]
- Sutherland, R.; Dos Santos, Z.; Agnini, C.; Alegret, L.; Lam, A.R.; Asatryan, G. Neogene mass accumulation rate of carbonate sediment in Tasman Sea, southwest Pacific. Paleoceanogr. Paleoclimatol. 2022, 37, e2021PA004294. [Google Scholar] [CrossRef]
- Hills, S.J.; Thierstein, H.R. Plio-Pleistocene calcareous plankton biochronology. Mar. Micropaleontol. 1989, 14, 67–96. [Google Scholar] [CrossRef]
- Schlitzer, R. Ocean Data View. 2018. Available online: Httsp://odv.awi.de (accessed on 11 February 2021).
- Locarnini, R.A.; Mishonov, A.V.; Antonov, J.I.; Boyer, T.P.; Garcia, H.E.; Seidov, D. World Ocean Atlas 2013, Volume 1: Temperature; Levitus, S., Michonov, A., Eds.; NOAA Atlas NESDIS 73; Department of Commerce: Silver Spring, ML, USA, 2013.
- Bé, A.W. An ecological, zoogeographic and taxonomic review of recent planktonic foraminifera. Ocean. Micropaleontol. 1977, 1, 76–88. [Google Scholar]
- Vincent, E.; Berger, W.H. Planktonic foraminifera and their use in paleoceanography. In The Oceanic Lithosphere; Emiliani, C., Ed.; Wiley-Interscience: New York, NY, USA, 1981; Volume 7, pp. 1–100. [Google Scholar]
- Kučera, M. Planktonic foraminifera as tracers of past oceanic environments. Dev. Mar. Geol. 2007, 1, 213–262. [Google Scholar]
- Reynolds, R.W.; Rayner, N.A.; Smith, T.M.; Stokes, D.C.; Wang, W. An improved in situ and satellite SST analysis for climate. J. Clim. 2002, 15, 1609–1625. [Google Scholar] [CrossRef]
- National Geographic Society. Tropics. National Geographic Society. 9 October 2012. Available online: https://www.nationalgeographic.org/encyclopedia/tropics/ (accessed on 3 April 2022).
- American Meteorological Society. Subtropics—Glossary of Meteorology. 25 April 2012. Available online: https://glossary.ametsoc.org/wiki/Subtropics (accessed on 3 April 2022).
- Bralower, T.J.; Silva, I.P.; Malone, M.J. Proceedings of the Ocean Drilling Program, Initial Reports 198; Ocean Drilling Program: College Station, TX, USA, 2002. [Google Scholar] [CrossRef]
- Kroenke, L.W.; Berger, W.H.; Janecek, T.R.; Backman, J.; Bassinot, F.; Corfield, R.M.; Delaney, M.L.; Hagen, R.; Jansen, E.; Krissek, L.A.; et al. Proceedings of the Ocean Drilling Program, Initial Reports; Ocean Drilling Program: College Station, TX, USA, 1991; Volume 130. [Google Scholar] [CrossRef]
- Kennett, J.P.; von der Borch, C.C.; Baker, P.A.; Barton, C.E.; Boersma, A.; Caulet, J.P.; Dudley, W.C.; Gardner, J.V.; Jenkins, G.; Lohman, W.H.; et al. Initial Reports of the Deep Sea Drilling Project; U.S. Government Printing Office: Washington, DC, USA, 1986; Volume 90. [Google Scholar] [CrossRef]
- Cooke, P.J.; Nelson, C.S.; Crundwell, M.P. Miocene isotope zones, paleotemperatures, and carbon maxima events at intermediate water-depth, Site 593, Southwest Pacific. N. Z. J. Geol. Geophys. 2008, 51, 1–22. [Google Scholar] [CrossRef]
- Sager, W.W.; Sano, T.; Geldmacher, J. Formation and evolution of Shatsky Rise oceanic plateau: Insights from IODP Expedition 324 and recent geophysical cruises. Earth-Sci. Rev. 2016, 159, 306–336. [Google Scholar] [CrossRef] [Green Version]
- Sager, W.W.; Zhang, J.; Korenaga, J.; Sano, T.; Koppers, A.A.; Widdowson, M.; Mahoney, J.J. An immense shield volcano within the Shatsky Rise oceanic plateau, northwest Pacific Ocean. Nat. Geosci. 2013, 6, 976–981. [Google Scholar] [CrossRef]
- Takayama, T. Notes on Neogene calcareous nannofossil biostratigraphy of the Ontong Java Plateau and size variations of Reticulofenestra coccoliths. In Proceedings of the Ocean Drilling Program Scientific Results; Berger, W.H., Kroenke, L.W., Janecek, T.R., Backman, J., Bassinot, F., Corfield, R.M., Delaney, M.L., Hagen, R., Jansen, E., Krissek, L.A., et al., Eds.; Ocean Drilling Program: College Station, TX, USA, 1993; Volume 130, pp. 179–229. [Google Scholar]
- Kennett, J.P.; von der Borch, C.C.; Baker, P.A.; Barton, C.E.; Boersma, A.; Caulet, J.P.; Dudley, W.C.; Gardner, J.V.; Jenkins, G.; Lohman, W.H.; et al. (Eds.) Shipboard Scientific Party. Introduction and Objectives, Techniques, and Explanatory Notes. In Initial Reports of the Deep Sea Drilling Program; U.S. Government Printing Office: Washington, DC, USA, 1986; Volume 90, pp. 3–16. [Google Scholar]
- Oke, P.R.; Pilo, G.S.; Ridgway, K.; Kiss, A.; Rykova, T. A search for the Tasman Front. J. Mar. Syst. 2019, 199, 103217. [Google Scholar] [CrossRef]
- Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D.; Ogg, G.M. Geologic Time Scale 2020; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Sutherland, R.; Dickens, G.R.; Blum, P.; Agnini, C.; Alegret, L.; Zhou, X. Expedition 371 methods. In Proceedings of the International Ocean Discovery Program; Sutherland, R., Dickens, G.R., Blum, P., Agnini, C., Alegret, L., Bhattacharya, J., Bordenave, A., Chang, L., Collot, J., Cramwinckel, M., et al., Eds.; International Ocean Discovery Program: College Station, TX, USA, 2019; Volume 371. [Google Scholar] [CrossRef] [Green Version]
- Wade, B.S.; Olsson, R.K.; Pearson, P.N.; Huber, B.T.; Berggren, W.A. Atlas of Oligocene Planktonic Foraminifera; Cushman Foundation for Foraminiferal Research: Washington, DC, USA, 2018. [Google Scholar]
- Crundwell, M.P. New Zealand Late Miocene Biostratigraphy and Biochronology: Studies of Planktic Foraminifers and Bolboforms at Oceanic Sites 593 and 1123, and Selected Onland Sections. Ph.D. Thesis, The University of Waikato, Hamilton, New Zealand, 2004. Available online: https://hdl.handle.net/10289/13214 (accessed on 1 December 2021).
- Scott, G.H. Coiling excursions in Globorotalia miotumida: High resolution bioevents at the middle-upper Miocene boundary in southern temperate water masses? J. Foraminifer. Res. 1995, 25, 299–308. [Google Scholar] [CrossRef]
- Barton, C.E.; Bloemendal, J. Paleomagnetism of sediments collected during Leg 90, southwest Pacific. In Initial Reports of the Deep Sea Drilling Project; Kennett, J.P., von der Borch, C.C., Baker, P.A., Barton, C.E., Boersma, A., Caulet, J.P., Dudley, W.C., Gardner, J.V., Jenkins, G., Lohman, W.H., et al., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1986; Volume 90, pp. 1273–1316. [Google Scholar]
- Lohman, W.H. Calcareous nannoplankton biostratigraphy of the southern Coral Sea, Tasman Sea, and southwestern Pacific Ocean, Deep Sea Drilling Project Leg 90: Neogene and Quaternary. In Initial Reports of the Deep Sea Drilling Project; Kennett, J.P., von der Borch, C.C., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1986; Volume 90, pp. 763–793. [Google Scholar]
- Grützmacher, U.J. Die veränderungen der paläogeographischen verbreitung von Bolboforma—ein beitrag zur rekonstruktion und definition vor wassermassen in Tertiär. GEOMAR Res. Cent. Mar. Geosci. Kiel Rep. 1993, 22, 1–104. [Google Scholar]
- Crundwell, M.P.; Nelson, C.S. A magnetostratigraphically-constrained chronology for late Miocene bolboformids and planktic foraminifers in the temperate Southwest Pacific. Stratigraphy 2007, 4, 1–34. [Google Scholar]
- Raine, J.I.; Beu, A.G.; Boyes, A.F.; Campbell, H.J.; Cooper, R.A.; Crampton, J.S.; Crundwell, M.P.; Hollis, C.J.; Morgans, H.E.G. Revised Calibration of the New Zealand Geological Timescale: NZGT2015/1. GNS Sci. Rep. 2015, 39, 1–39. [Google Scholar]
- Coxall, H.K.; Pearson, P.N.; Wilson, P.A.; Sexton, P.F. Iterative evolution of digitate planktonic foraminifera. Paleobiology 2007, 33, 495–516. [Google Scholar] [CrossRef] [Green Version]
- Chaproniere, G.H.; Styzen, M.J.; Sager, W.W.; Nishi, H.; Quinterno, P.J. Late Neogene biostratigraphic and magnetostratigraphic synthesis, Leg 135. In Proceedings of the Ocean Drilling Program, Scientific Results; Hawkins, J., Parson, L., Allan, J., Abrahamsen, N., Bednarz, U., Blanc, G., Bloomer, S.H., Bøe, R., Bruns, T.R., Bryan, W.B., et al., Eds.; Ocean Drilling Program: College Station, TX, USA, 1994; Volume 135, pp. 857–877. [Google Scholar]
- Pearson, P.N.; Penny, L. Coiling directions in the planktonic foraminifer Pulleniatina: A complex eco-evolutionary dynamic spanning millions of years. PLoS ONE 2021, 16, e0249113. [Google Scholar] [CrossRef]
- Keigwin, L.D.; Prell, W.L.; Gardner, J.V. Neogene planktonic foraminifers from Deep Sea Drilling Project sites 502 and 503. In Initial Reports of the Deep Sea Drilling Project; Prell, W.L., Gardner, J.V., Adelseck, C.G., Jr., Blechschmidt, G., Fleet, A.J., Keigwin, L.D., Kent, D.V., Ledbetter, M.T., Mann, U., Mayer, L.A., et al., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1982; Volume 68, pp. 269–288. [Google Scholar]
- Morgans, H.E.; Edwards, A.R.; Scott, G.H.; Graham, I.J.; Kamp, P.J.; Mumme, T.C.; Wilson, G.S. Integrated stratigraphy of the Waitakian-Otaian Stage boundary stratotype, Early Miocene, New Zealand. N. Z. J. Geol. Geophys. 1999, 42, 581–614. [Google Scholar] [CrossRef] [Green Version]
- Chaisson, W.P.; Pearson, P.N. Planktonic foraminifer biostratigraphy at Site 925: Middle Miocene-Pliocene. In Proceedings of the Ocean Drilling Program Scientific Results; Shackleton, N.J., Curry, W.B., Richter, C., Bralower, T.J., Eds.; Ocean Drilling Program: Station, TX, USA, 1997; Volume 154, pp. 3–31. [Google Scholar]
- Aze, T.; Ezard, T.H.; Purvis, A.; Coxall, H.K.; Stewart, D.R.; Wade, B.S.; Pearson, P.N. A phylogeny of Cenozoic macroperforate planktonic foraminifera from fossil data. Biol. Rev. 2011, 86, 900–927. [Google Scholar] [CrossRef]
- Fabbrini, A.; Zaminga, I.; Ezard, T.H.; Wade, B.S. Systematic taxonomy of middle Miocene Sphaeroidinellopsis (planktonic foraminifera). J. Syst. Palaeontol. 2021, 19, 953–968. [Google Scholar] [CrossRef]
- Wei, K.-Y.; Kennett, J.P. Phyletic gradualism and punctuated equilibrium in the Late Neogene planktic foraminiferal clade Globoconella. Paleobiology 1988, 14, 345–363. [Google Scholar] [CrossRef]
- Chapman, M.R.; Funnell, B.M.; Weaver, P.P.E. Isolation, extinction and migration within Late Pliocene populations of the planktonic foraminiferal lineage Globorotalia (Globoconella) in the North Atlantic. Mar. Micropaleontol. 1998, 33, 203–222. [Google Scholar] [CrossRef]
- Scott, G.H.; Bishop, S.; Burt, B.J. Guide to some Neogene Globorotalids (Foraminiferida) from New Zealand; New Zealand Geological Survey: Lower Hutt, New Zealand, 1990. [Google Scholar]
- Jenkins, D.G. Planktonic foraminiferal zones and new taxa from the lower Miocene to the Pleistocene of New Zealand. N. Z. J. Geol. Geophys. 1967, 10, 1064–1078. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, D.G. New Zealand Cenozoic planktonic foraminifera. Palaeont. Bull. N. Z. Geol. Surv. 1971, 42, 1–278. [Google Scholar]
- Hornibrook, N.D.B. Globorotalia (planktic Foraminiferida) in the late Pliocene and early Pleistocene of New Zealand. N. Z. J. Geol. Geophys. 1981, 24, 263–292. [Google Scholar] [CrossRef]
- Hornibrook ND, B. Late Miocene to Pleistocene Globorotalia (Foraminiferida) from DSDP Leg 29, Site 284, Southwest Pacific. N. Z. J. Geol. Geophys. 1982, 25, 83–99. [Google Scholar] [CrossRef]
- Olsson, R.K.; Hemleben, C.; Huber, B.T.; Berggren, W.A. Taxonomy, biostratigraphy, and phylogeny of Eocene Globigerina, Globoturborotalita, Subbotina, and Turborotalita. In Atlas of Eocene Planktonic Foraminifera; Pearson, P.N., Olsson, R.K., Huber, B.T., Hemleben, C., Berggren, W.A., Eds.; Cushman Foundation for Foraminiferal Research Special Publication: Washington, DC, USA, 2006; Volume 41, pp. 111–168. [Google Scholar]
- Molnar, P. Closing of the Central American Seaway and the Ice Age: A critical review. Paleoceanography 2008, 23, PA22001. [Google Scholar] [CrossRef]
- Poore, R.Z.; Tauxe, L.; Percival, S.F., Jr.; Labrecque, J.L.; Wright, R.; Petersen, N.P.; Hsü, K. Late Cretaceous—Cenozoic magnetostratigraphic and biostratigraphic correlations of the South Atlantic Ocean: DSDP Leg 73. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1983, 42, 127–149. [Google Scholar] [CrossRef]
- Pujol, C.; Duprat, J. Quaternary planktonic foraminifers of the southwestern Atlantic (Rio Grande Rise) Deep Sea Drilling Project Leg 72. In Initial Reports of the Deep Sea Drilling Project; Barker, P.F., Carlson, R.L., Johnson, D.A., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1983; Volume 72, pp. 601–622. [Google Scholar]
- Grazzini, C.V. Non-equilibrium isotopic compositions of shells of planktonic foraminifera in the Mediterranean Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1976, 20, 263–276. [Google Scholar] [CrossRef]
- Shackleton, N.J.; Vincent, E. Oxygen and carbon isotope studies in recent foraminifera from the southwest Indian Ocean. Mar. Micropaleontol. 1978, 3, 1–13. [Google Scholar] [CrossRef]
- Hays, J.D.; Saito, T.; Opdyke, N.D.; Burckle, L.H. Pliocene-Pleistocene sediments of the equatorial Pacific: Their paleomagnetic, biostratigraphic, and climatic record. Geol. Soc. Am. Bull. 1969, 80, 1481–1514. [Google Scholar] [CrossRef]
- Mayer, L.; Pisias, N.; Janecek, T. (Eds.) Shipboard Scientific Party. Site 846. In Proceedings of the Ocean Drilling Program Initial Reports; Ocean Drilling Program: College Station, TX, USA, 1992; Volume 138, pp. 265–333. [Google Scholar]
- Pearson, P.N.; Shackleton, N.J. Neogene multispecies planktonic foraminifer stable isotope record, Site 871, Limalok Guyot. In Proceedings of the Ocean Drilling Program Scientific Results; Silva, I.P., Haggerty, J., Rack, F., Arnaud-Vanneau, A., Bergersen, D.D., Bogdanov, Y., Bohrmann, H.W., Buchardt, B., Camoin, G., Christie, D.M., et al., Eds.; Ocean Drilling Program: College Station, TX, USA, 1995; Volume 144, pp. 401–410. [Google Scholar]
- Stewart, D.R.M. Evolution of Neogene Globorotaliid Foraminifera and Miocene Climate Change. Ph.D. Thesis, University of Bristol, Bristol, UK, 2003. [Google Scholar]
- Nathan, S.A.; Leckie, R.M. Early history of the Western Pacific Warm Pool during the middle to late Miocene (~13.2–5.8 Ma): Role of sea-level change and implications for equatorial circulation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 274, 140–159. [Google Scholar] [CrossRef]
- Venti, N.L.; Billups, K. Stable-isotope stratigraphy of the Pliocene–Pleistocene climate transition in the northwestern subtropical Pacific. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 326, 54–65. [Google Scholar] [CrossRef]
- Carter, R.M.; McCave, I.N.; Richter, C.; Carter, L.; Aita, Y.; Buret, C.; Di Stefano, A.; Fenner, J.; Fothergill, F.; Gradstein, F.; et al. Proceedings of the Ocean Drilling Program; Initial Reports Volume 181; Ocean Drilling Program: College Station, TX, USA, 1999. [Google Scholar]
- Al-Sabouni, N.; Fenton, I.S.; Telford, R.J.; Kučera, M. Reproducibility of species recognition in modern planktonic foraminifera and its implications for analyses of community structure. J. Micropalaeontol. 2018, 37, 519–534. [Google Scholar] [CrossRef]
- Fenton, I.S.; Baranowski, U.; Boscolo-Galazzo, F.; Cheales, H.; Fox, L.; King, D.J.; Purvis, A. Factors affecting consistency and accuracy in identifying modern macroperforate planktonic foraminifera. J. Micropalaeontol. 2018, 37, 431–443. [Google Scholar] [CrossRef]
- Boltovskoy, E. Twilight of foraminiferology. J. Paleontol. 1965, 39, 383–390. [Google Scholar]
- Leckie, R.M.; Wade, B.S.; Pearson, P.N.; Fraass, A.J.; King, D.J.; Olsson, R.K.; Berggren, W.A. Taxonomy, biostratigraphy, and phylogeny of Oligocene and Early Miocene Paragloborotalia and Parasubbotina. In Atlas of Oligocene Planktonic Foraminifera; Wade, B.S., Olsson, P.N., Pearson, P.N., Huber, B.T., Berggren, W.A., Eds.; Cushman Foundation for Foraminiferal Research: Lawrence, KS, USA, 2018; Volume 46, pp. 125–178. [Google Scholar]
- Huber, B.T.; Petrizzo, M.R.; Young, J.R.; Falzoni, F.; Gilardoni, S.E.; Bown, P.R.; Wade, B.S. Pforams@ microtax. Micropaleontology 2016, 62, 429–438. [Google Scholar] [CrossRef]
- Olsson, R.K.; Berggren, W.A.; Hemleben, C.; Huber, B.T. Atlas of Paleocene Planktonic Foraminifera; Smithsonian Institute Press: Washington, DC, USA, 1999. [Google Scholar]
- Pearson, P.N.; Olsson, R.K.; Huber, B.T.; Hemleben, C.; Berggren, W.A. Atlas of Eocene Planktonic Foraminifera; Cushman Foundation for Foraminiferal Research Special Publication: Washington, DC, USA, 2005. [Google Scholar]
- Fraass, A.J.; Leckie, R.M. Oligocene Planktic Foraminiferal Taxonomy and Evolution: An Illustrated Revision of Ocean Drilling Program Site 803. J. Foraminifer. Res. 2021, 51, 139–164. [Google Scholar] [CrossRef]
- Fehrenbacher, J.; Fritz-Enders, T.; Schell, F.; Meyer, G.; Lane, M.K.; Kelly, J. Foraminarium. Available online: https://www.foraminarium.com/ (accessed on 12 December 2021).
- Keller, G. Late Neogene biostratigraphy and paleoceanography of DSDP Site 310 Central North Pacific and correlation with the Southwest Pacific. Mar. Micropaleontol. 1978, 3, 97–119. [Google Scholar] [CrossRef]
- Blow, W.H. Late Middle Eocene to Recent planktonic foraminiferal biostratigraphy. In Proceedings of the First International Conference Planktonic Microfossils 1967; EJ Brill: Leiden, The Netherlands, 1969; Volume 1, pp. 199–242. [Google Scholar]
- Jenkins, D.G. Planktonic foraminifera and Tertiary intercontinental correlations. Micropaleontology 1965, 11, 265–277. [Google Scholar] [CrossRef]
- Saito, T. Geologic significance of coiling direction in the planktonic foraminifera Pulleniatina. Geology 1976, 4, 305–309. [Google Scholar] [CrossRef]
- Hornibrook, N.D.B. Correlation of Pliocene biostratigraphy, magnetostratigraphy and O18 fluctuations in New Zealand and DSDP Site 284. Newsl. Stratigr. 1980, 9, 114–120. [Google Scholar] [CrossRef]
- Crundwell, M.P.; Woodhouse, A. A detailed biostratigraphic framework for 0–1.2 Ma Quaternary sediments of north-eastern Zealandia. N. Z. J. Geol. Geophys. 2022, 1–14. [Google Scholar] [CrossRef]
- Scott, G.H.; Kennett, J.P.; Wilson, K.J.; Hayward, B.W. Globorotalia puncticulata: Population divergence, dispersal and extinction related to Pliocene–Quaternary water masses. Mar. Micropaleontol. 2007, 62, 235–253. [Google Scholar] [CrossRef]
- Hornibrook ND, B. Globorotalia (planktic foraminifera) at the Miocene/Pliocene boundary in New Zealand. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1984, 46, 107–117. [Google Scholar] [CrossRef]
- Wilkens, R.H.; Westerhold, T.; Drury, A.J.; Lyle, M.; Gorgas, T.; Tian, J. Revisiting the Ceara Rise, equatorial Atlantic Ocean: Isotope stratigraphy of ODP Leg 154 from 0 to 5 Ma. Clim. Past 2017, 13, 779–793. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, H.; Idemitsu, K.; Wade, B.S.; Idehara, Y.; Kimoto, K.; Nishi, H.; Matsui, H. Middle Miocene to Pleistocene planktonic foraminiferal biostratigraphy in the eastern equatorial Pacific Ocean. Paleontol. Res. 2013, 17, 91–109. [Google Scholar] [CrossRef]
- Ogg, J.G.; Ogg, G.M.; Gradstein, F.M. Neogene. In A Concise Geologic Time Scale; Ogg, J.G., Ogg, G.M., Gradstein, F.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 201–203. [Google Scholar]
- Drury, A.J.; Westerhold, T.; Frederichs, T.; Tian, J.; Wilkens, R.; Channell, J.E.; Röhl, U. Late Miocene climate and time scale reconciliation: Accurate orbital calibration from a deep-sea perspective. Earth Planet. Sci. Lett. 2017, 475, 254–266. [Google Scholar] [CrossRef]
- Kuhnt, W.; Holbourn, A.; Hall, R.; Zuvela, M.; Käse, R. Neogene history of the Indonesian throughflow. In Continent-Ocean Interactions within East Asian Marginal Seas. Geophysical Monograph; Clift, P.D., Wang, P., Kuhnt, W., Hayes, D.E., Eds.; American Geophysical Union: Washington, DC, USA, 2004; Volume 149, pp. 299–320. [Google Scholar]
- Vincent, E. Indian Ocean Neogene planktonic foraminiferal biostratigraphy and its paleoceanographic implications. Indian Ocean. Geol. Biostratigraphy 1977, 20, 469–584. [Google Scholar]
- Srinivasan, M.S. Recent advances in Neogene planktonic foraminiferal biostratigraphy, chemostratigraphy and paleoceanography, Northern Indian Ocean. J. Palaeontol. Soc. India 1989, 34, 1–18. [Google Scholar]
- Sinha, D.K.; Singh, A.K. Late Neogene planktic foraminiferal biochronology of the ODP Site 763a, Exmouth Plateau, southeast Indian Ocean. J. Foraminifer. Res. 2008, 38, 251–270. [Google Scholar] [CrossRef]
- Groeneveld, J.; De Vleeschouwer, D.; McCaffrey, J.C.; Gallagher, S.J. Dating the northwest shelf of Australia since the Pliocene. Geochem. Geophys. Geosyst. 2021, 22, e2020GC009418. [Google Scholar] [CrossRef]
- de Vargas, C.; Norris, R.; Zaninetti, L.; Gibb, S.W.; Pawlowski, J. Molecular evidence of cryptic speciation in planktonic foraminifers and their relation to oceanic provinces. Proc. Natl. Acad. Sci. USA 1999, 96, 2864–2868. [Google Scholar] [CrossRef] [Green Version]
- Norris, R.D. Hydrographic and tectonic control of plankton distribution and evolution. In Reconstructing Ocean History; Abrantes, F., Mix, A.C., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 173–193. [Google Scholar]
- Darling, K.F.; Wade, C.M.; Kroon, D.; Brown AJ, L.; Bijma, J. The diversity and distribution of modern planktic foraminiferal small subunit ribosomal RNA genotypes and their potential as tracers of present and past ocean circulations. Paleoceanography 1999, 14, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Norris, R.D. Species diversity, biogeography, and evolution in the pelagic realm. In Deep Time: Paleobiology’s Perspective; Erwin, D.H., Wing, S.L., Eds.; Paleontological Society: Baltimore, MA, USA, 2000; pp. 237–258. [Google Scholar]
- de Vargas, C.; Bonzon, M.; Rees, N.W.; Pawlowski, J.; Zaninetti, L. A molecular approach to biodiversity and biogeography in the planktonic foraminifer Globigerinella siphonifera (d’Orbigny). Mar. Micropaleontol. 2002, 45, 101–116. [Google Scholar] [CrossRef]
- Sexton, P.F.; Norris, R.D. Dispersal and biogeography of marine plankton: Long-distance dispersal of the foraminifer Truncorotalia truncatulinoides. Geology 2008, 36, 899–902. [Google Scholar] [CrossRef]
- Bradshaw, J.S. Ecology of living planktonic foraminifera in the north and equatorial Pacific Ocean. Contrib. Cushman Found. Foraminifer. Res. 1959, 10, 25–64. [Google Scholar]
- Schneider, C.E.; Kennett, J.P. Isotopic evidence for interspecies habitat differences during evolution of the Neogene planktonic foraminiferal clade Globoconella. Paleobiology 1996, 22, 282–303. [Google Scholar] [CrossRef]
- Rutherford, S.; D’Hondt, S.; Prell, W. Environmental controls on the geographic distribution of zooplankton diversity. Nature 1999, 400, 749–753. [Google Scholar] [CrossRef]
- Tittensor, D.P.; Mora, C.; Jetz, W.; Lotze, H.K.; Ricard, D.; Berghe, E.V.; Worm, B. Global patterns and predictors of marine biodiversity across taxa. Nature 2010, 466, 1098–1101. [Google Scholar] [CrossRef]
- Yasuhara, M.; Hunt, G.; Dowsett, H.J.; Robinson, M.M.; Stoll, D.K. Latitudinal species diversity gradient of marine zooplankton for the last three million years. Ecol. Lett. 2012, 15, 1174–1179. [Google Scholar] [CrossRef]
- Fenton, I.S.; Pearson, P.N.; Dunkley Jones, T.; Purvis, A. Environmental predictors of diversity in recent planktonic foraminifera as recorded in marine sediments. PLoS ONE 2016, 11, e0165522. [Google Scholar] [CrossRef] [Green Version]
- Fenton, I.S.; Pearson, P.N.; Jones, T.D.; Farnsworth, A.; Lunt, D.J.; Markwick, P.; Purvis, A. The impact of Cenozoic cooling on assemblage diversity in planktonic foraminifera. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150224. [Google Scholar] [CrossRef] [Green Version]
- Scott, G.H. Globorotalia inflata lineage and G. crassaformis from Blind River, New Zealand: Recognition, relationship, and use in uppermost Miocene-lower Pliocene biostratigraphy. N. Z. J. Geol. Geophys. 1980, 23, 665–677. [Google Scholar]
- DeConto, R.M.; Pollard, D. Rapid Cenozoic glaciation of Antarctica induced by declining atmospheric CO2. Nature 2003, 421, 245–249. [Google Scholar] [CrossRef]
- Kennett, J.P. Cenozoic evolution of Antarctic glaciation, the circum-Antarctic Ocean, and their impact on global paleoceanography. J. Geophys. Res. 1977, 82, 3843–3860. [Google Scholar] [CrossRef]
- Kennett, J.P. The development of planktonic biogeography in the Southern Ocean during the Cenozoic. Mar. Micropaleontol. 1978, 3, 301–345. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X.; Dong, C. Anatomy of a cyclonic eddy in the Kuroshio extension based on high-resolution observations. Atmosphere 2019, 10, 553. [Google Scholar] [CrossRef] [Green Version]
- Kennett, J.P.; Keller, G.; Srinivasan, M.S. Miocene planktonic foraminiferal biogeography and pale-oceanographic development of the Indo-Pacific region. In The Miocene Ocean: Paleoceanography and Biogeography; Kennett, J.P., Ed.; Geological Society of America: Boulder, CO, USA, 1985; Volume 163, pp. 197–236. [Google Scholar]
- Wu, L.; Cai, W.; Zhang, L.; Nakamura, H.; Timmermann, A.; Joyce, T.; Giese, B. Enhanced warming over the global subtropical western boundary currents. Nat. Clim. Chang. 2012, 2, 161–166. [Google Scholar] [CrossRef]
- Matsui, H.; Horikawa, K.; Chiyonobu, S.; Itaki, T.; Ikehara, M.; Kawagata, S.; Okazaki, Y. Integrated Neogene biochemostratigraphy at DSDP Site 296 on the Kyushu-Palau Ridge in the western North Pacific. Newsl. Stratigr. 2020, 53, 313–331. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Latitude | Longitude | Water Depth (m) | Core Section, Interval (cm) | Depth in Cores (mbsf) | Age (Ma) |
|---|---|---|---|---|---|---|
| 1207A | 37.90° N | 162.76° E | 3100.8 | 1H-1, 29–31 to 18H-4, 78–80 | 0.29–162.08 | 0.018–12.304 |
| 1208A | 36.29° N | 158.22° E | 3345.7 | 1H-1, 77–79 to 35X-2, 77–79 | 0.78–317.58 | 0.014–15.117 |
| 1209A | 32.65° N | 158.50° E | 2387.2 | 1H-1, 27–29 to 11H-1, 77–79 | 0.27–94.48 | 0.019–7.161 |
| 806B | 0.34° N | 159.53° E | 2519.9 | 1H-2, 12–14 to 66X-CC | 1.62–627.51 | 0.230–22.800 |
| 586B | 0.71° S | 158.71° E | 2207.0 | 1H-1, 51–53 to 21H-5, 50–52 | 1.92–197.31 | 0.097–7.391 |
| 587 | 21.20° S | 161.59° E | 1101.0 | 1H-1, 5–6 to 10H-6, 17–18 | 0.05–87.57 | 0.002–8.250 |
| 588 | 26.29° S | 161.38° E | 1533.0 | 1H-CC to 25H-CC | 5.43–235.90 | 0.395–9.401 |
| 590 | 31.17° S | 163.49° E | 1299.0 | 1H-CC to 27H-CC | 6.86–280.80 | 0.487–7.251 |
| 591 | 31.59° S | 164.68° E | 2131.0 | 1H-CC to 31H-CC | 3.19–282.83 | 0.147–7.626 |
| 592 | 36.57° S | 165.58° E | 1088.0 | 1H-CC to 29X-CC | 4.27–272.44 | 0.303–11.925 |
| 593 | 40.63° S | 167.79° E | 1068.0 | 1H-2, 87–89 to 45X-CC | 2.38–427.39 | 0.112–18.049 |
| Species | Evolutionary Event | 1207A Age | 1208A Age | 1209A Age | 806B Age | 586B Age | 587 Age | 588 Age | 590 Age | 591 Age | 592 Age | 593 Age | Tropical Age |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Globigerinella calida | Base | 3.870 ± 0.088 | 1.589 ± 0.018 | 3.139 ± 0.102 | 4.062 ± 0.072 | 1.369 ± 0.035 | 0.795 0.041 | 1.801 ± 0.056 | 2.178 ± 0.253 | 2.509 ± 0.160 | - | - | 0.22 |
| Globigerinoides subquadratus | Top | 8.470 ± 0.213 | 8.443 ± 0.178 | 6.480 ± 0.043 | 11.056 ± 0.011 | - | - | 9.010 ± 0.041 | - | - | 11.963 ± 0.017 | 8.130 ± 0.165 | 11.57 |
| Globigerinoides extremus | Top | 2.093 ± 0.097 | 1.625 ± 0.018 | 1.583 ± 0.032 | 2.172 ± 0.092 | 1.921 ± 0.010 | 2.458 ± 0.018 | 2.671 ± 0.035 | 2.917 ± 0.155 | 2.825 ± 0.018 | 3.945 ± 0.140 | 2.662 ± 0.269 | 1.97 |
| Base | 6.054 ± 0.101 | 5.777 ± 0.036 | 6.209 ± 0.008 | 8.438 ± 0.040 | - | 8.147 ± 0.103 | 8.342 ± 0.049 | - | - | 11.746 ± 0.079 | 3.170 ± 0.238 | 8.83 | |
| Globigerinoides conglobatus | Base | 7.203 ± 0.009 | 6.561 ± 0.091 | 6.299 ± 0.048 | 5.953 ± 0.014 | - | - | 5.849 ± 0.161 | 7.042 ± 0.093 | 6.882 ± 0.143 | 0.397 ± 0.094 | - | 6.21 |
| Globigerinoides obliquus | Top | 0.293 ± 0.011 | 1.668 ± 0.024 | 1.328 ± 0.033 | 1.821 ± 0.064 | 2.978 ± 0.051 | 3.964 ± 0.118 | 3.891 ± 0.008 | 6.238 ± 0.030 | - | - | - | 1.3 |
| Globigerinoidesella fistulosa | Top | - | - | - | 1.736 ± 0.019 | 1.736 ± 0.052 | 2.093 ± 0.134 | 1.632 ± 0.113 | 1.765 ± 0.161 | - | - | - | 1.88 |
| Base | - | - | - | 2.801 ± 0.086 | 3.740 ± 0.354 | 3.266 ± 0.091 | 3.866 ± 0.016 | 2.917 ± 0.155 | - | - | - | 3.33 | |
| Pulleniatina primalis | Top | 0.900 ± 0.001 | 1.906 ± 0 | 1.583 ± 0.032 | 2.042 ± 0.036 | 1.866 ± 0.007 | 3.266 ± 0.091 | 3.515 ± 0.056 | 3.577 ± 0.056 | 3.596 ± 0.072 | 5.177 ± 0.024 | - | 3.66 |
| Base | 5.095 ± 0.297 | 2.335 ± 0.023 | 1.678 ± 0.160 | 6.587 ± 0.015 | 6.998 ± 0.040 | 7.138 ± 0.023 | 6.154 ± 0.145 | 5.547 ± 0.015 | 5.680 ± 0.124 | 5.223 ± 0.022 | - | 6.57 | |
| Globoquadrina dehiscens | Top | 8.972 ± 0.791 | 9.053 ± 0.097 | 6.747 ± 0.149 | 5.686 ± 0.040 | 5.345 ± 0.015 | 7.941 ± 0.103 | 7.964 ± 0.285 | - | 6.882 ± 0.143 | 9.557 ± 0.184 | 8.861 ± 0.015 | 5.91 |
| Sphaeroidinella dehiscens | Base | 3.732 ± 0.126 | 3.490 ± 0.022 | 3.718 ± 0.181 | 4.585 ± 0.400 | 4.904 ± 0.013 | 3.964 ± 0.118 | 3.745 ± 0.025 | 3.970 ± 0.081 | 3.515 ± 0.008 | - | - | 5.54 |
| Sphaeroidinellopsis kochi | Top | 4.471 ± 0.031 | 4.324 ± 0.024 | 3.058 ± 0.058 | 4.987 ± 0.072 | - | - | - | - | - | - | - | 4.49 |
| Sphaeroidinellopsis seminulina | Top | 3.487 ± 0.033 | 3.762 ± 0.028 | 2.913 ± 0.010 | 3.625 ± 0.032 | 3.270 ± 0.029 | 2.899 ± 0.099 | 3.177 ± 0.208 | 3.243 ± 0.171 | 3.151 ± 0.187 | 3.463 ± 0.341 | 12.250 ± 0.256 | 3.59 |
| Globoconella conomiozea | Top | 3.434 ± 0.008 | 3.490 ± 0.022 | 3.281 ± 0.022 | - | - | - | 5.532 ± 0.156 | 5.475 ± 0.058 | 4.898 ± 0.021 | 5.277 ± 0.032 | 5.145 ± 0.105 | - |
| Base | 4.915 ± 0.063 | 4.753 ± 0.033 | 4.341 ± 0.034 | - | - | - | 6.807 ± 0.012 | 6.371 ± 0.104 | 5.498 ± 0.058 | 7.198 ± 0.141 | 6.650 ± 0.343 | - | |
| Globoconella puncticulata | Base | 4.821 ± 0.025 | 4.632 ± 0 | 3.862 ± 0.036 | - | - | 4.885 ± 0.094 | 4.245 ± 0.021 | 5.475 ± 0.058 | 4.705 ± 0.019 | 5.420 ± 0.048 | 5.730 ± 0.006 | - |
| Globoconella inflata | Base | 3.364 ± 0.016 | 3.289 ± 0.029 | 3.018 ± 0.007 | 0.978 ± 0.085 | - | 3.758 ± 0.087 | 3.832 ± 0.018 | 3.770 ± 0.015 | 3.827 ± 0.007 | 4.412 ± 0.092 | 4.395 ± 0.247 | - |
| Globoturborotalita nepenthes | Top | 4.498 ± 0.022 | 4.889 ± 0.012 | 4.025 ± 0.072 | 4.276 ± 0.021 | 4.904 ± 0.013 | 4.203 ± 0.121 | 3.891 ± 0.008 | 3.837 ± 0.052 | 4.215 ± 0.006 | 5.341 ± 0.032 | 5.043 ± 0.003 | 4.38 |
| Base | 11.967 ± 0.096 | 12.371 ± 0.110 | - | 11.112 ± 0.043 | - | - | - | - | - | 10.506 ± 0.090 | 10.619 ± 0.133 | 11.67 | |
| Globoturborotalita decoraperta | Top | 3.163 ± 0.114 | 4.926 ± 0.025 | 3.535 ± 0.023 | 2.399 ± 0.007 | 1.892 ± 0.019 | 3.758 ± 0.087 | 1.476 ± 0.042 | 3.243 ± 0.171 | 1.002 ± 0.025 | 3.463 ± 0.341 | 1.929 ± 0.172 | 2.74 |
| Base | 8.972 ± 0.791 | 9.178 ± 0.028 | - | 11.408 ± 0.047 | - | - | - | - | - | 11.564 ± 0.286 | 13.356 ± 0.181 | 11.51 | |
| Globoturborotalita apertura | Top | 2.873 ± 0.035 | 2.517 ± 0.018 | 2.372 ± 0.281 | 1.821 ± 0.064 | 3.206 ± 0.035 | - | 2.671 ± 0.035 | 2.198 ± 0.155 | - | - | - | 1.64 |
| Base | 10.74 ± 0.061 | 9.354 ± 0.148 | - | 11.056 ± 0.011 | 7.094 ± 0.056 | - | 4.689 ± 0.231 | 5.318 ± 0.100 | - | - | - | 11.24 | |
| Globoturborotalita woodi | Top | 0.914 ± 0.027 | 0.427 ± 0.018 | 0.708 ± 0.035 | 2.172 ± 0.092 | 1.921 ± 0.010 | 2.568 ± 0.091 | 3.422 ± 0.037 | 3.681 ± 0.048 | 2.948 ± 0.016 | 2.461 ± 0.126 | 2.247 ± 0.146 | 2.3 |
| Neogloboquadrina acostaensis | Top | 0.914 ± 0.027 | 3.289 ± 0.029 | 0.451 ± 0.035 | 5.213 ± 0.039 | 4.904 ± 0.013 | 3.964 ± 0.118 | 4.193 ± 0.030 | 4.176 ± 0.124 | 4.368 ± 0.062 | - | - | 1.58 |
| Base | 11.156 ± 0.064 | 7.205 ± 0.004 | 4.970 ± 0.041 | 10.355 ± 0.039 | - | - | 9.010 ± 0.041 | 6.525 ± 0.050 | 7.538 ± 0.087 | - | - | 9.81 | |
| Fohsella lenguaensis | Top | 5.629 ± 0.614 | 11.469 ± 0.107 | - | 6.018 ± 0.050 | - | - | - | - | 5.971 ± 0.166 | - | - | 6.14 |
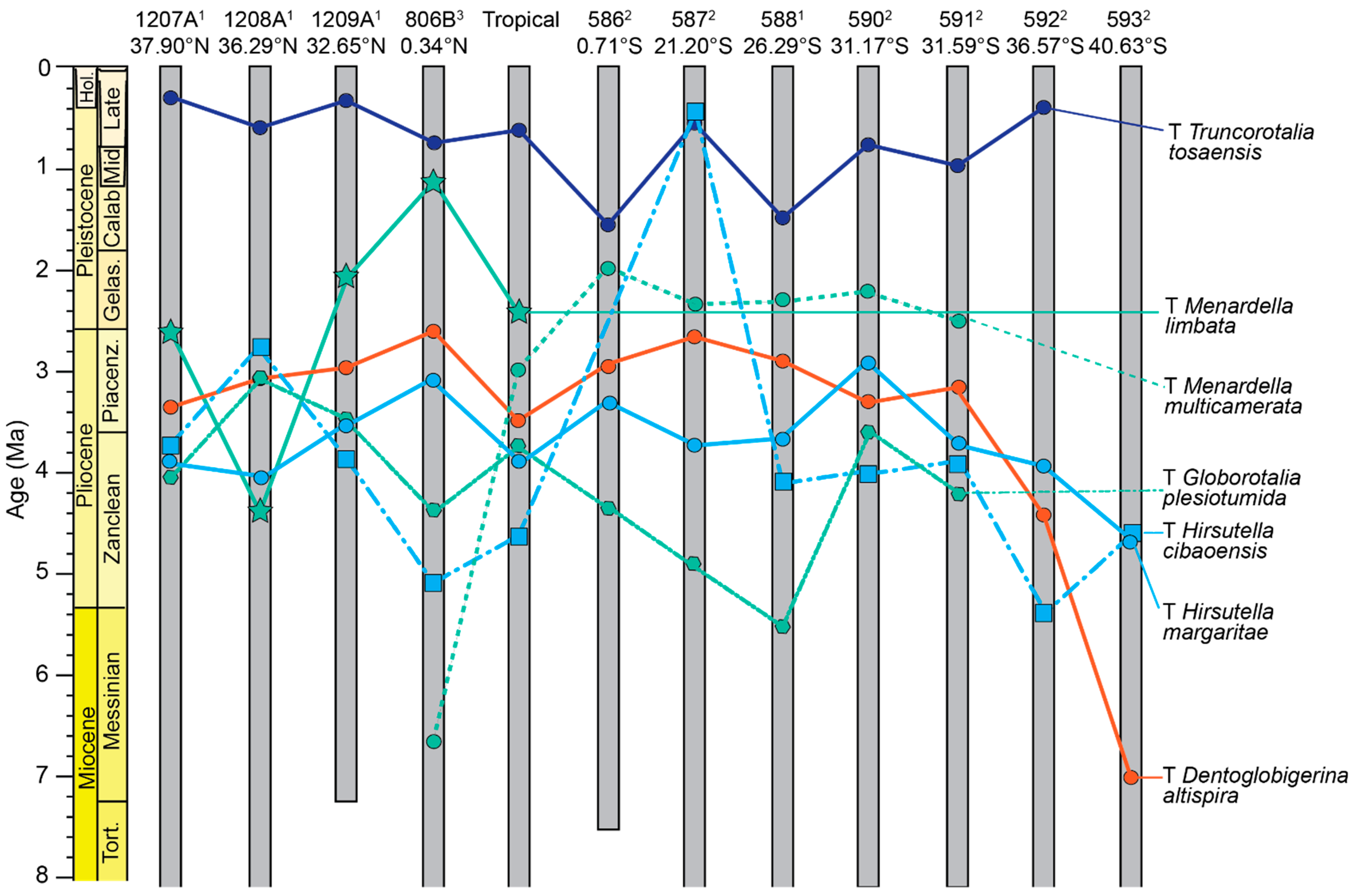
| Hirsutella margaritae | Top | 3.969 ± 0.022 | 4.009 ± 0.029 | 3.473 ±0.041 | 3.075 ± 0.111 | 3.343 ± 0.029 | 3.758 ± 0.087 | 3.745 ± 0.025 | 2.918 ± 0.155 | 3.720 ± 0.053 | 3.945 ± 0.140 | 4.671 ± 0.029 | 3.83 |
| Base | 6.707 ± 0.183 | 6.561 ± 0.091 | 4.970 ± 0.041 | 5.686 ± 0.040 | 5.082 ± 0.004 | 5.119 ± 0.029 | 6.779 ± 0.016 | 5.547 ± 0.015 | 5.680 ± 0.124 | 5.341 ± 0.032 | 4.793 ± 0.093 | 6.09 | |
| Hirsutella cibaoensis | Top | 3.732 ± 0.126 | 2.747 ± 0.018 | 3.862 ± 0.036 | 5.118 ± 0.032 | - | 0.431 ± 0.005 | 4.088 ± 0.013 | 3.970 ± 0.081 | 3.875 ± 0.007 | 5.341 ± 0.032 | 4.671 ± 0.029 | 4.61 |
| Base | 7.773 ± 0.522 | 8.120 ± 0.017 | 4.970 ± 0.041 | 8.013 ± 0.039 | - | - | 8.256 ± 0.007 | 7.193 ± 0.058 | 6.550 ± 0.189 | 8.100 ± 0.186 | 8.972 ± 0.207 | 9.44 | |
| Hirsutella hirsuta | Base | 1.123 ± 0.046 | 0.755 ± 0.027 | 0.673 ± 0.035 | - | - | 0.582 ± 0.026 | - | - | - | 2.216 ± 0.119 | 0.115 ± 0.007 | 0.45 |
| Truncorotalia crassaformis | Base | 3.969 ± 0.022 | 4.224 ± 0.021 | 3.535 ± 0.023 | 3.675 ± 0.17 | 3.740 ± 0.354 | 4.440 ± 0.115 | 5.148 ± 0.228 | 4.982 ± 0.071 | 4.239 ± 0.018 | 5.003 ± 0.028 | 4.671 ± 0.029 | 4.3 |
| Truncorotalia tosaensis | Top | 0.293 ± 0.011 | 0.586 ± 0.032 | 0.417 ± 0.035 | 0.735 ± 0.084 | 1.539 ± 0.035 | 0.530 ± 0.026 | 1.476 ± 0.042 | 0.736 ± 0.249 | 1.002 ± 0.025 | 0.397 ± 0.094 | - | 0.61 |
| Base | 2.150 ± 0.018 | 3.328 ± 0.009 | 2.947 ± 0.019 | 3.075 ± 0.111 | 3.270 ± 0.029 | 3.266 ± 0.091 | 3.515 ± 0.056 | 3.578 ± 0.056 | 3.935 ± 0.052 | 2.663 ± 0.076 | - | 3.35 | |
| Truncorotalia truncatulinoides | Base | 2.150 ± 0.018 | 1.994 ± 0.019 | 2.372 ± 0.281 | 2.332 ± 0.067 | 2.262 ± 0.002 | 2.899 ± 0.099 | 2.869 ± 0.100 | 2.597 ± 0.165 | 2.825 ± 0.018 | 2.663 ± 0.076 | 2.556 ± 0.004 | 1.92 |
| Globorotalia tumida | Base | 5.629 ± 0.614 | 5.383 ± 0.036 | 4.474 ± 0.101 | 5.806 ± 0.050 | 5.345 ± 0.015 | 5.257 ± 0.061 | 5.849 ± 0.161 | 4.852 ± 0.058 | 5.035 ± 0.116 | 5.177 ± 0.024 | - | 5.57 |
| Globorotalia plesiotumida | Top | 4.036 ± 0.022 | 3.025 ± 0.026 | 3.473 ± 0.041 | 4.370 ± 0.072 | 4.339 ± 0.030 | 4.885 ± 0.094 | 5.532 ± 0.156 | 3.578 ± 0.056 | 4.215 ± 0.006 | - | - | 3.72 |
| Base | 8.972 ± 0.791 | 6.561 ± 0.091 | 7.151 ± 0.019 | 7.924 ± 0.050 | - | - | 7.583 ± 0.096 | 5.902 ± 0.100 | - | - | - | 8.77 | |
| Menardella multicamerata | Top | - | - | - | 6.653 ± 0.050 | 1.921 ± 0.010 | 2.333 ± 0.107 | 2.273 ± 0.107 | 2.179 ± 0.253 | 2.509 ± 0.160 | - | - | 2.97 |
| Menardella limbata | Top | 2.595 ± 0.020 | 4.372 ± 0.024 | 2.082 ± 0.300 | 1.126 ± 0.114 | - | - | - | - | - | - | - | 2.37 |
| Dentoglobigerina altispira | Top | 3.348 ± 0.016 | 3.074 ± 0.022 | 2.928 ± 0.020 | 2.603 ± 0.111 | 2.978 ± 0.051 | 2.568 ± 0.091 | 2.869 ± 0.100 | 3.243 ± 0.171 | 3.151 ± 0.187 | 4.412 ± 0.092 | 7.002 ± 0.180 | 3.47 |
| Species | Evolutionary Event | NW Pacific Diachroneity (Myr) | SW Pacific Diachroneity (Myr) | WE Pacific Diachroneity (Myr) |
|---|---|---|---|---|
| Globigerinella calida | Base | 2.281 | 1.714 | 2.693 |
| Globigerinoides subquadratus | Top | 1.990 | 3.883 | - |
| Globigerinoides extremus | Top | 0.510 | 1.487 | 0.251 |
| Base | 0.432 | 8.576 | - | |
| Globigerinoides conglobatus | Base | 0.904 | 6.645 | - |
| Globigerinoides obliquus | Top | 1.375 | 2.274 | 1.157 |
| Globigerinoidesella fistulosa | Top | - | 0.461 | 0.000 |
| Base | - | 0.949 | 0.939 | |
| Pulleniatina primalis | Top | 1.006 | 0.311 | 0.176 |
| Base | 3.417 | 1.915 | 0.411 | |
| Globoquadrina dehiscens | Top | 2.306 | 2.675 | 0.341 |
| Sphaeroidinella dehiscens | Base | 0.242 | 0.455 | 0.319 |
| Sphaeroidinellopsis kochi | Top | 1.413 | - | - |
| Sphaeroidinellopsis seminulina | Top | 0.574 | 9.351 | 0.355 |
| Globoconella conomiozea | Top | 0.209 | 0.634 | - |
| Base | 0.574 | 1.700 | - | |
| Globoconella puncticulata | Base | 0.959 | 1.485 | - |
| Globoconella inflata | Base | 0.346 | 0.654 | - |
| Globoturborotalita nepenthes | Top | 0.864 | 1.504 | 0.628 |
| Base | 0.404 | 0.113 | - | |
| Globoturborotalita decoraperta | Top | 1.763 | 2.756 | 0.507 |
| Base | 0.206 | 1.792 | - | |
| Globoturborotalita apertura | Top | 0.501 | 0.473 | 1.385 |
| Base | 1.386 | 0.629 | 3.962 | |
| Globoturborotalita woodi | Top | 0.487 | 1.434 | 0.251 |
| Neogloboquadrina acostaensis | Top | 2.838 | 0.404 | 0.309 |
| Base | 6.186 | 2.485 | - | |
| Fohsella lenguaensis | Top | 5.840 | - | - |
| Hirsutella margaritae | Top | 0.536 | 1.753 | 0.268 |
| Base | 1.737 | 1.986 | 0.604 | |
| Hirsutella cibaoensis | Top | 1.115 | 4.910 | - |
| Base | 3.150 | 2.422 | - | |
| Hirsutella hirsuta | Base | 0.450 | 2.101 | - |
| Truncorotalia crassaformis | Base | 0.689 | 0.764 | 0.065 |
| Truncorotalia tosaensis | Top | 0.293 | 1.079 | 0.804 |
| Base | 1.178 | 1.272 | 0.195 | |
| Truncorotalia truncatulinoides | Base | 0.378 | 0.343 | 0.070 |
| Globorotalia tumida | Base | 1.155 | 0.997 | 0.461 |
| Globorotalia plesiotumida | Top | 1.011 | 1.307 | 0.031 |
| Base | 2.411 | 1.681 | - | |
| Menardella multicamerata | Top | - | 0.330 | 4.732 |
| Menardella limbata | Top | 2.290 | - | - |
| Dentoglobigerina altispira | Top | 0.420 | 4.434 | 0.375 |
| Average diachroneity | 1.396 | 2.052 | 0.819 | |
| Species | Evolutionary Event | U1463 (IO) Age | 806B (WEP) Age | Tropical Age | IO-WEP Diachroneity (Myr) | IO-Tropical Diachroneity (Myr) |
|---|---|---|---|---|---|---|
| Globigerinoides extremus | Top | 1.973 | 2.172 | 1.97 | 0.199 | 0.003 |
| Globigerinoidesella fistulosa | Top | 1.685 | 1.736 | 1.88 | 0.051 | 0.195 |
| Base | 3.155 | 2.801 | 3.33 | 0.354 | 0.175 | |
| Pulleniatina primalis | Top | 3.358 | 2.042 | 3.66 | 1.316 | 0.302 |
| Sphaeroidinellopsis kochi | Top | 3.131 | 4.987 | 4.49 | 1.856 | 1.359 |
| Sphaeroidinellopsis seminulina | Top | 3.335 | 3.625 | 3.59 | 0.290 | 0.255 |
| Globoconella inflata | Base | 1.908 | 0.978 | - | 0.930 | - |
| Globoturborotalita nepenthes | Top | 4.200 | 4.276 | 4.38 | 0.076 | 0.180 |
| Globoturborotalita woodi | Top | 2.512 | 2.172 | 2.3 | 0.340 | 0.212 |
| Hirsutella margaritae | Top | 3.131 | 3.075 | 3.83 | 0.056 | 0.699 |
| Truncorotalia tosaensis | Base | 3.059 | 3.075 | 3.35 | 0.016 | 0.291 |
| Truncorotalia truncatulinoides | Base | 2.159 | 2.332 | 1.92 | 0.173 | 0.239 |
| Globorotalia plesiotumida | Top | 3.358 | 4.37 | 3.72 | 1.012 | 0.362 |
| Menardella limbata | Top | 1.837 | 1.126 | 2.37 | 0.711 | 0.533 |
| Dentoglobigerina altispira | Top | 2.697 | 2.603 | 3.47 | 0.094 | 0.773 |
| Average diachroneity | 0.498 | 0.398 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, A.R.; Crundwell, M.P.; Leckie, R.M.; Albanese, J.; Uzel, J.P. Diachroneity Rules the Mid-Latitudes: A Test Case Using Late Neogene Planktic Foraminifera across the Western Pacific. Geosciences 2022, 12, 190. https://0-doi-org.brum.beds.ac.uk/10.3390/geosciences12050190
Lam AR, Crundwell MP, Leckie RM, Albanese J, Uzel JP. Diachroneity Rules the Mid-Latitudes: A Test Case Using Late Neogene Planktic Foraminifera across the Western Pacific. Geosciences. 2022; 12(5):190. https://0-doi-org.brum.beds.ac.uk/10.3390/geosciences12050190
Chicago/Turabian StyleLam, Adriane R., Martin P. Crundwell, R. Mark Leckie, James Albanese, and Jacob P. Uzel. 2022. "Diachroneity Rules the Mid-Latitudes: A Test Case Using Late Neogene Planktic Foraminifera across the Western Pacific" Geosciences 12, no. 5: 190. https://0-doi-org.brum.beds.ac.uk/10.3390/geosciences12050190