
Improved Planktonic, Benthic Foraminiferal and Nannofossil Biostratigraphy Aids the Interpretation of the Evolution at Hole U1468A: IODP Expedition 359, the Maldives

 , and
, and
Abstract
:1. Introduction
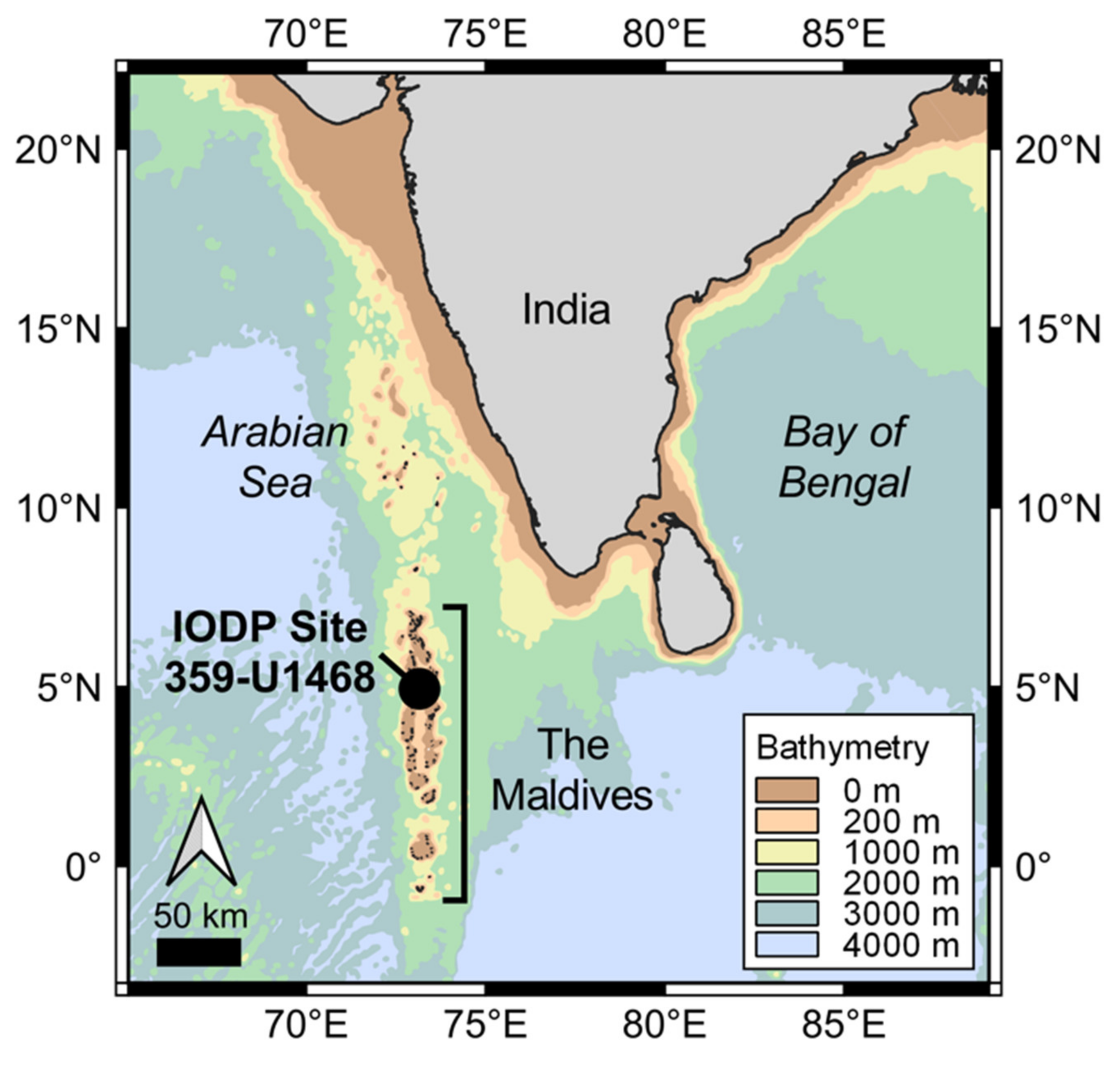
2. Geological Setting
3. Materials and Methods
4. Results (All Biostratigraphic Data Are Reported from Bottom to Top)
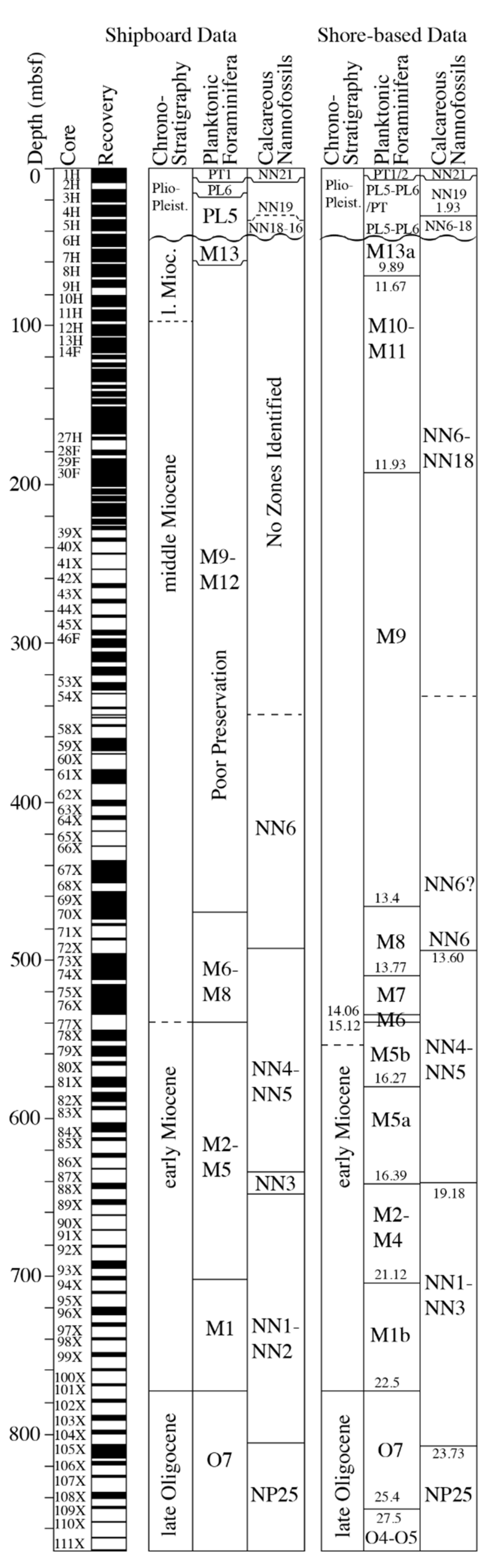
4.1. The Biostratigraphy of Hole U1468A
4.1.1. Planktonic Foraminifera


4.1.2. Benthic Foraminifera
4.1.3. Calcareous Nannoplankton
5. Discussion
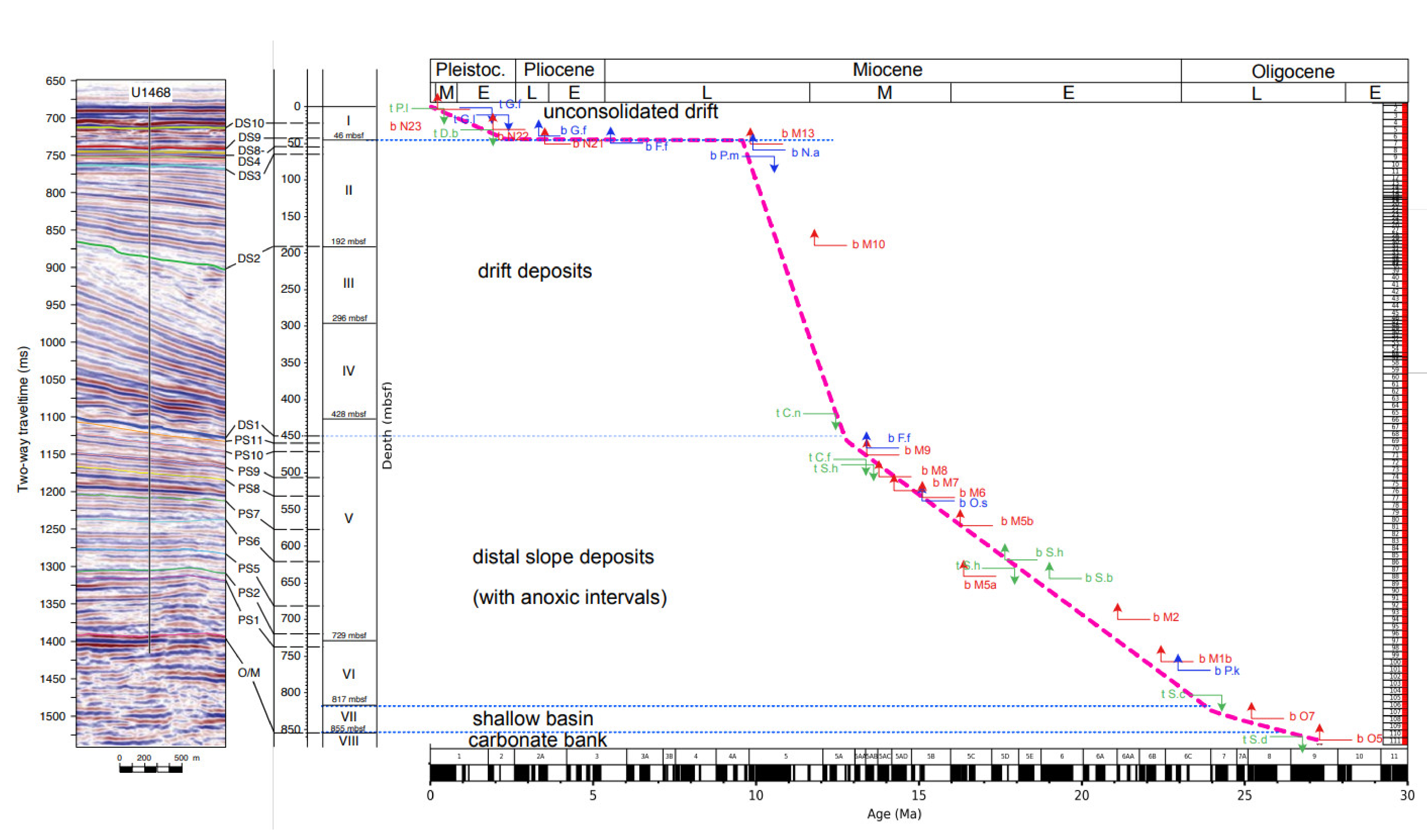
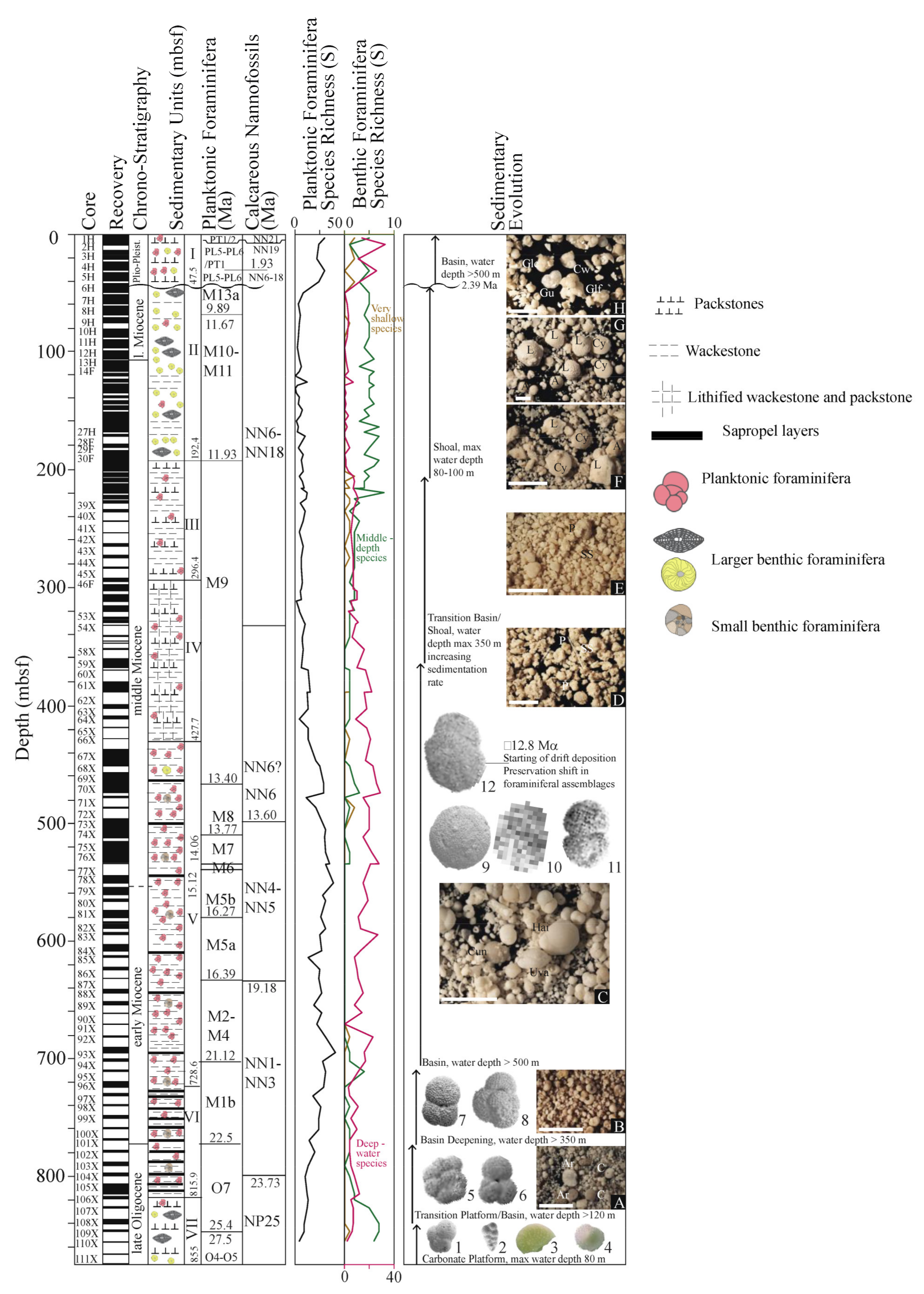
5.1. Biostratigraphy and Sedimentation Rates at Hole U1468A
5.2. Paleo-Bathymetric Evolution at Hole U1468A
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Betzler, C.; Fürstenau, J.; Lüdmann, T.; Hübscher, C.; Lindhorst, S.; Paul, A.; Reijmer, J.J.G.; Droxler, A.W. Sea-level and ocean-current control on carbonate-platform growth, Maldives, Indian Ocean. Basin Res. 2013, 25, 172–196. [Google Scholar] [CrossRef]
- Betzler, C.; Luedmann, T.; Reijmer, J.; Eberli, G.; Swart, P.; Droxler, A.; Tiwari, M.; Gischler, E.; Hübscher, C. Currents, Monsoon and Sea Level in the Indian Ocean: The Neogene of the Maldives; Full Proposal 820; IODP: College Station, TX, USA, 2013. [Google Scholar]
- Parker, J.H.; Gischler, E. Modern foraminiferal distribution and diversity in two atolls from the Maldives, Indian Ocean. Mar. Micropaleontol. 2011, 78, 30–49. [Google Scholar] [CrossRef]
- Storz, D.; Gischler, E.; Parker, J.; Klostermann, L. Changes in diversity and assemblages of foraminifera through the Holocene in an atoll from the Maldives, Indian Ocean. Mar. Micropaleontol. 2014, 106, 40–54. [Google Scholar] [CrossRef]
- Aubert, O.; Droxler, A.W. Seismic stratigraphy and depositional signatures of the Maldive carbonate system (Indian Ocean). Mar. Pet. Geol. 1996, 13, 503–536. [Google Scholar] [CrossRef]
- Belopolsky, A.V.; Droxler, A.W. Seismic expressions of prograding carbonate bank margins: Middle Miocene, Maldives, Indian Ocean. In Seismic Imaging of Carbonate Reservoirs and Systems; Eberli, G.P., Masaferro, J.L., Sarg, J.F., Eds.; American Association of Petroleum Geologists: Calgary, AB, Canada, 2004; pp. 267–290. [Google Scholar]
- Betzler, C.; Lüdmann, T.; Hübscher, C.; Fürstenau, J. Current and sea-level signals in periplatform ooze (Neogene, Maldives, Indian Ocean). Sediment. Geol. 2013, 290, 126–137. [Google Scholar] [CrossRef]
- Lüdmann, T.; Kalvelage, C.; Betzler, C.; Fürstenau, J.; Hübscher, C. The Maldives, a giant isolated carbonate platform dominated by bottom currents. Mar. Pet. Geol. 2013, 43, 326–340. [Google Scholar] [CrossRef]
- Premoli Silva, I.; Spezzaferri, S. Paleogene planktonic foraminifer biostratigraphy and paleoenvironmental remarks on Paleogene sediments from Indian Ocean sites, Leg 115. In Proceedings of the Ocean Drilling Program, Scientific Results; Duncan, R.A., Backmann, J., Peterson, L.C., Eds.; Ocean Drilling Program: College Station. TX, USA, 1990; pp. 277–314. [Google Scholar] [CrossRef]
- Vincent, E.; Toumarkine, M. Neogene planktonic foraminifers from the western tropical Indian Ocean, Leg 115. In Proceedings of the Ocean Drilling Program, Scientific Results; Duncan, R.A., Backman, J., Peterson, L.C., Eds.; Ocean Drilling Program: College Station, TX, USA, 1990; Volume 115, pp. 795–836. [Google Scholar]
- Nicora, A.; Premoli Silva, I. Paleogene Shallow-Water Larger Foraminifers from Holes 714A and 715A, Leg 115, Indian Ocean. In Proceedings of the Ocean Drilling Program, Scientific Results; Ocean Drilling Program: College Station, TX, USA, 1990; Volume 115, pp. 381–393. [Google Scholar]
- Backman, J.; Duncan, R.A.; Peterson, L.C.; Baker, P.A.; Baxter, A.N.; Boersma, A.; Cullen, J.L.; Droxler, A.W.; Fisk, M.R.; Greenough, J.D.; et al. Proceedings of the Ocean Drilling Program, Initial Reports; Ocean Drilling Program: College Station, TX, USA, 1988; Volume 115. [Google Scholar]
- Aubert, O.; Droxler, A.W. General Cenozoic evolution of the Maldives carbonate system (equatorial Indian Ocean). Bull. Des Cent. Rech. Explor. Elf Aquitaine 1992, 16, 113–136. [Google Scholar]
- Purdy, E.G.; Bertram, G.T. Carbonate concepts from the Maldives, Indian Ocean; American Association of Petroleum Geologists: Tulsa, OK, USA, 1993. [Google Scholar]
- Betzler, C.; Eberli, G.P.; Alvarez Zarikian, C.A.; and the Expedition 359 Scientists. Maldives Monsoon and Sea Level, Proceedings of the International Ocean Discovery Program, 359; International Ocean Discovery Program: College Station, TX, USA, 2017. [Google Scholar] [CrossRef]
- Lüdmann, T.; Betzler, C.; Eberli, G.P.; Reolid, J.; Reijmer, J.J.G.; Sloss, C.R.; Bialik, O.M.; Alvarez-Zarikian, C.A.; Alonso-Garcia, M.; Blättler, C.L.; et al. Carbonate delta drift: A new sediment drift type. Mar. Geol. 2018, 401, 98–111. [Google Scholar] [CrossRef]
- Reolid, J.; Betzler, C.; Lüdmann, T. Facies and sedimentology of a carbonate delta drift (Miocene, Maldives). Sedimentology 2019, 66, 1243–1265. [Google Scholar] [CrossRef]
- Coletti, G.; Stainbank, S.; Fabbrini, A.; Spezzaferri, S.; Foubert, A.; Kroon, D.; Betzler, C. Biostratigraphy of large benthic foraminifera from Hole U1468A (Maldives): A CT-scan taxonomic approach. Swiss J. Geosci. 2018, 111, 495–508. [Google Scholar] [CrossRef] [Green Version]
- Blow, W.H. Late Middle Eocene to Recent planktonic foraminifera biostratigraphy. In Proceedings of the First International Conference on Planktonic Microfossils; Brönniman, P., Renz, H.H., Eds.; E.J. Brill: Leiden, The Netherlands, 1969; pp. 199–422. [Google Scholar]
- Blow, W.H. A study of the morphology, taxonomy, evolutionary relationships and the stratigraphical distribution of some Globigerinida (mainly Globigerinacea). In The Cainozoic Globigerinida; E.J. Brill: Leiden, The Netherlands, 1979; pp. 1–1413. [Google Scholar]
- Berggren, W.A.; Kent, D.V.; Swisher, C.C., III; Aubry, M.-P. A revised Cenozoic geochronology and chronostratigraphy. In Geochronology, Time Scales and Global Stratigraphic Correlation: A Unified Temporal Framework for an Historical Geology; Berggren, W.A., Kent, D.V., Aubry, M.-P., Hardenbol, J., Eds.; SEPM Society for Sedimentary Geology: Broken Arrow, OK, USA, 1995; pp. 129–212. [Google Scholar]
- Wade, B.S.; Pearson, P.N.; Berggren, W.A.; Pälike, H. Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale. Earth-Sci. Rev. 2011, 104, 111–142. [Google Scholar] [CrossRef] [Green Version]
- Raffi, I.; Wade, B.S.; Pälike, H. Chapter 29—The Neogene Period. In Geologic Time Scale 2020; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1141–1215. [Google Scholar]
- Thompson, P.R.; Bé, A.W.H.; Duplessy, J.-C.; Shackleton, N.J. Disappearance of pink-pigmented Globigerinoides ruber at 120,000 yr BP in the Indian and Pacific Oceans. Nature 1979, 280, 554–558. [Google Scholar] [CrossRef]
- Kennett, J.P.; Srinivasan, M.S. Neogene Planktonic Foraminifera—A Phylogenetic Atlas; Hutchinson Ross Publishing Company: Stroudsburg, PA, USA, 1983. [Google Scholar]
- Bolli, H.M.; Saunders, J.B. Oligocene to Holocene low latitude planktic foraminifera. In Plankton Stratigraphy; Bolli, H.M., Saunders, J.B., Perch-Neilsen, K., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 155–262. [Google Scholar]
- Wade, B.S.; Olsson, R.K.; Pearson, P.N.; Huber, B.T.; Berggren, W.A. Atlas of Oligocene Planktonic Foraminifera; No. 46; Cushman Foundation for Foraminiferal Research Special Publication: Lawrence, KS, USA, 2018. [Google Scholar]
- Huber, B.T.; Petrizzo, M.R.; Young, J.R.; Falzoni, F.; Gilardoni, S.E.; Bown, P.R.; Wade, B.S. Pforams@microtax: A new online taxonomic database for planktonic foraminifera. Micropaleontology 2016, 62, 429–438. [Google Scholar] [CrossRef]
- Van Morkhoven, F.M.; Berggren, W.A.; Edwards, A.S. Cenozoic cosmopolitan deep-water benthic foraminifera. Bull. Des Cent. Rech. Explor. Elf-Aquitaine 1986, 11, 421. [Google Scholar]
- Sgarella, F.; Moncharmont Zei, M. Benthic foraminifera of the Gulf of Naples (Italy): Systematics and autecology. Boll. Della Soc. Paleontol. Ital. 1993, 32, 145–264. [Google Scholar]
- Sen Gupta, B.; Smith, L.E.; Machain-Castillo, M.L. Foraminifera of the Gulf of Mexico. In Gulf of Mexico–Origins, Waters, and Biota: Biodiversity; Texas A and M Press: College Station, TX, USA, 2009; pp. 87–129. [Google Scholar]
- Loeblich, A.; Tappan, H. Foraminifera of the Sahul Shelf and Timor Sea; Cushman Foundation for Foraminiferal Research, Dept. of Invertebrate Paleontology, Museum of Comparative Zoology: Cambridge, MA, USA, 1994; Volume 31, p. 661. [Google Scholar]
- Jones, R.W. The Challenger Foraminifera; Oxford University Press: Oxford, NY, USA, 1994. [Google Scholar]
- Hayward, B.W.; Carter, R.; Grenfell, H.R.; Hayward, J.J. Depth distribution of Recent deep-sea benthic foraminifera east of New Zealand, and their potential for improving paleobathymetric assessments of Neogene microfaunas. N. Z. J. Geol. Geophys. 2001, 44, 555–587. [Google Scholar] [CrossRef]
- Spezzaferri, S.; Rögl, F.; Coric, S.; Hohenegger, J. Paleoenvironmental changes and agglutinated foraminifera across the Karpatian/Badenian (Early/Middle Miocene) boundary in the Styrian Basin (Austria, Central Paratethys). In Proceedings of the Sixth International Workshop on Agglutinated Foraminifera, New York, NY, USA, 1–7 September 2001; Buík, M., Kaminski, M.A., Eds.; Grzybowski Foundation Special Publication: Krakow, Poland, 2004; Volume 8, pp. 423–459. [Google Scholar]
- Hohenegger, J. Estimation of environmental paleogradient values based on presence/absence data: A case study using benthic foraminifera for paleodepth estimation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 217, 115–130. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: New York, NY, USA, 2006. [Google Scholar]
- Holbourn, A.; Henderson, A.S.; Macleod, N. Atlas of Benthic Foraminifera; Blackwell: Hoboken, NJ, USA, 2013. [Google Scholar]
- Debenay, J.-P. A Guide to 1000 Foraminifera from Southwestern Pacific: New Caledonia; IRD Editions Marseille/Publication Scientifiques du Muséum: Paris, France, 2012. [Google Scholar]
- Horton, W.C. Foraminifera of the Cenozoic and Recent Genus Sphaerogypsina Galloway. Master’s Thesis, University of Missouri, Columbia, MO, USA, 1962. [Google Scholar]
- Renema, W. Terrestrial influence as a key driver of spatial variability in large benthic foraminiferal assemblage composition in the Central Indo-Pacific. Earth-Sci. Rev. 2018, 177, 514–544. [Google Scholar] [CrossRef]
- Chaproniere, G.C.H. Palaeoecology of Oligo-Miocene larger Foraminiferida, Australia. Alcheringa 1975, 1, 37–58. [Google Scholar] [CrossRef]
- Hottinger, L. Distribution of larger Peneroplidae, Borelis and Nummulitidae in the Gulf of Elat, Red Sea. Utr. Micropaleontol. Bull. 1977, 15, 35–109. [Google Scholar]
- Hallock, P.; Glenn, E.C. Larger foraminifera: A tool for paleoenvironmental analysis of Cenozoic carbonate depositional facies. Palaios 1986, 1, 55–64. [Google Scholar] [CrossRef]
- Geel, T. Recognition of stratigraphic sequences in carbonate platform and slope deposits: Empirical models based on microfacies analysis of Palaeogene deposits in southeastern Spain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 155, 211–238. [Google Scholar] [CrossRef]
- Hohenegger, J. Coenoclines of larger foraminifera. Micropaleontology 2000, 46, 127–151. [Google Scholar]
- Beavington-Penney, S.J.; Racey, A. Ecology of extant nummulitids and other larger benthic foraminifera: Applications in palaeoenvironmental analysis. Earth-Sci. Rev. 2004, 67, 219–265. [Google Scholar] [CrossRef]
- Drobne, K.; Ćosović, V.; Čretnik, J.; Turk, J.; Briguglio, A.; Rögl, F.; Praprotnik, A. Differences and similarities of the Eocene to recent Sphaerogypsina tests collected from the Pannonian basin to the Adriatic Sea. In Proceedings of the EGU General Assembly Conference Abstracts, Vienna, Austria, 23–28 April 2017; p. 17042. [Google Scholar]
- Novak, V.; Renema, W. Ecological tolerances of Miocene larger benthic foraminifera from Indonesia. J. Asian Earth Sci. 2018, 151, 301–323. [Google Scholar] [CrossRef]
- Bown, P.R.; Young, J.R. Techniques. In Calcareous Nannofossil Biostratigraphy (British Micropalaeontological Society Publications Series); Bown, P.R., Ed.; Chapman and Kluwer Academic: London, UK, 1988; pp. 16–28. [Google Scholar]
- Young, J.R. Neogene Nannofossils, 225th ed.; Kluwer Academic: Dordrecht, The Netherlands, 1998. [Google Scholar]
- Hine, N.; Weaver, P.P.E. Quaternary. In Calcareous Nannofossil Biostratigraphy; Bown, P.R., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 266–283. [Google Scholar]
- Young, J.; Geisen, M.; Cros, L.; Kleijne, A.; Sprengel, C.; Probert, I.; Østergaard, J. A guide to extant coccolithophore taxonomy. J. Nannoplankt. Res. Spec. Issue 2003, 1, 1–132. [Google Scholar]
- Young, J.R.; Bown, P.R.; Lees, J.A. Nannotax3 Website; International Nannoplankton Association: London, UK, 2021. [Google Scholar]
- Martini, E. Standard Tertiary and Quaternary calcareous nannoplankton zonation. In Proceedings of the Second Planktonic Conference, Roma, 1970; Farinacci, A., Ed.; Tecnoscienza: Roma, Italy, 1971; pp. 739–785. [Google Scholar]
- Backman, J.; Raffi, I.; Rio, D.; Fornaciari, E.; Pälike, H. Biozonation and biochronology of Miocene through Pleistocene calcareous nannofossils from low and middle latitudes. Newsl. Stratigr. 2012, 45, 221–244. [Google Scholar] [CrossRef]
- Raffi, I.; Backman, J.; Fornaciari, E.; Pälike, H.; Rio, D.; Lourens, L.; Hilgen, F. A review of calcareous nannofossil astrobiochronology encompassing the past 25 million years. Quat. Sci. Rev. 2006, 25, 3113–3137. [Google Scholar] [CrossRef]
- Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D.; Ogg, G.M. The Geologic Time Scale 2012; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Leckie, R.M.; Farnham, C.; Schmidt, M.G. Oligocene planktonic foraminifer biostratigraphy of Hole 803D (Ontong Java Plateau) and Hole 628A (Little Bahama Bank), and comparison with the southern high latitudes. In Proceedings of the Ocean Drilling Program, Scientific Results; Berger, W.H., Kroenke, L.W., Janecek, T.R., Backman, J., Bassinot, F., Corfield, R.M., Delaney, M.L., Hagen, R., Jansen, E., Krissk, L.A., et al., Eds.; Ocean Drilling Program: College Station, TX, USA, 1993; Volume 130, pp. 113–136. [Google Scholar]
- Piller, W.E.; Harzhauser, M.; Mandic, O. Miocene Central Paratethys stratigraphy-current status and future directions. Stratigraphy 2007, 4, 151–168. [Google Scholar]
- Spezzaferri, S. Planktonic foraminiferal biostratigraphy of the Oligocene and lower Miocene in the oceanic record. An overview. Palaeontogr. Ital. 1994, 81, 1–187. [Google Scholar]
- Young, J.R.; Archontikis, O.A.; Su, X.; Pratiwi, S.D. Nannofossil palaeoecology of Lower Miocene sapropels from IODP Expedition 359, the Maldives. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 571, 110325. [Google Scholar] [CrossRef]
- Spezzaferri, S.; Pearson, P. Distribution and ecology of Catapsydrax indianus, a new planktonic foraminifer index species for the late Oligocene-early Miocene. J. Foraminferal Res. 2009, 39, 112–119. [Google Scholar] [CrossRef]
- Spezzaferri, S. The Oligocene/Miocene boundary in the Lemme Section (Piedmont Basin, Northern Italy): Paleoclimatic evidence based on planktonic foraminifera. G. Geol. 1996, 58, 119–139. [Google Scholar]
- Iaccarino, S.; Borsetti, A.M.; Rögl, F. Planktonic foraminifera of the Neogene Lemme-Carrosio GSSP Section (Piedmont, Northern Italy). G. Geol. 1996, 58, 35–49. [Google Scholar]
- Groeneveld, J.; De Vleeschouwer, D.; McCaffrey, J.C.; Gallagher, S.J. Dating the Northwest Shelf of Australia since the Pliocene. Geochem. Geophys. Geosystems 2021, 22, e2020GC009418. [Google Scholar] [CrossRef]
- Lam, A.R.; Crundwell, M.P.; Leckie, R.M.; Albanese, J.; Uzel, J.P. Diachroneity rules the mid-latitudes: A test case using Late Neogene planktic foraminifera across the Western Pacific. Geosciences 2022, 12, 190. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Palaeoecology of Benthic Foraminifera; Longman Scientific and Technical: London, UK, 1991. [Google Scholar]
- Murray, J.W. Comparative studies of living and dead benthic foraminiferal distributions. In Foraminifera; Hedley, R.H., Adamas, C.G., Eds.; Academic Press Inc.: Cambridge, MA, USA, 1976; pp. 45–109. [Google Scholar]
- Coxall, H.K.; Spezzaferri, S. Taxonomy, biostratigraphy and phylogeny of Oligocene Catapsydrax, Globorotaloides and Protentelloides. Cushman Found. Foraminifer. Res. Spec. Publ. 2018, 46, 79–124. [Google Scholar]
- Stainbank, S.; Spezzaferri, S.; Kroon, D.; de Leau, E.S.; Rüggeberg, A. The Planktonic foraminifera Globigerinoides eoconglobatus n. sp. in a glacial–interglacial context: IODP359 Sites U1467 and U1468. Swiss J. Geosci. 2018, 111, 483–494. [Google Scholar] [CrossRef]
- Stainbank, S.; Spezzaferri, S.; Rüggeberg, A.; Raddatz, J.; de Leau, E.S.; Yu, S.M.; Zhang, M.; Kroon, D. Monsoon and tropical climate forcing on the physicochemical and thermocline characteristics of the Maldives Inner Sea: Insights from Marine Isotope Stages 1–2 and 10–13. Paleoceanogr. Paleoclimatol. 2021, 36, e2020PA004105. [Google Scholar] [CrossRef]
- Swart, P.K.; Blättler, C.L.; Nakakuni, M.; Mackenzie, G.J.; Betzler, C.; Eberli, G.P.; Reolid, J.; Alonso-García, M.; Slagle, A.L.; Wright, J.D.; et al. Cyclic anoxia and organic rich carbonate sediments within a drowned carbonate platform linked to Antarctic ice volume changes: Late Oligocene-early Miocene Maldives. Earth Planet. Sci. Lett. 2019, 521, 1–13. [Google Scholar] [CrossRef]
- Emeis, K.-C.; Robertson, A.H.F.; Richter, C.; Blanc-Valleron, M.-M.; Bouloubassi, I.; Brumsack, H.-J.; Cramp, A.; De Lange, G.J.; Stefano, E.D.; Flecker, R.; et al. Proceedings of the Ocean Drilling Program, Initial Reports; Ocean Drilling Program: College Station, TX, USA, 1998; Volume 160. [Google Scholar]
- Spezzaferri, S. Planktonic foraminiferal paleoclimatic implications across the Oligocene-Miocene transition in the oceanic record (Atlantic, Indian and South Pacific). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1995, 114, 43–74. [Google Scholar] [CrossRef]
- Haq, B.U.; Hardenbol, J.; Vail, P.R. Chronology of fluctuating sea levels since the Triassic. Science 1987, 235, 1156–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.G.; Mountain, G.S. Global sea-level change and the New Jersey margin. In Proceedings of the Ocean Drilling Program, Initial Reports; Ocean Drilling Program: College Station, TX, USA, 1994; Volume 150, pp. 11–20. [Google Scholar]
- Taylor, B.; Huchon, P.; Klaus, A.; Awadallah, S.A.M.; Brooks, C.K.; Célérier, B.; DeCarlo, E.H.; Floyd, J.; Frost, G.M.; Gardien, V.; et al. Proceedings of the Ocean Drilling Program, Initial Reports; Ocean Drilling Program: College Station, TX, USA, 2000; Volume 180. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
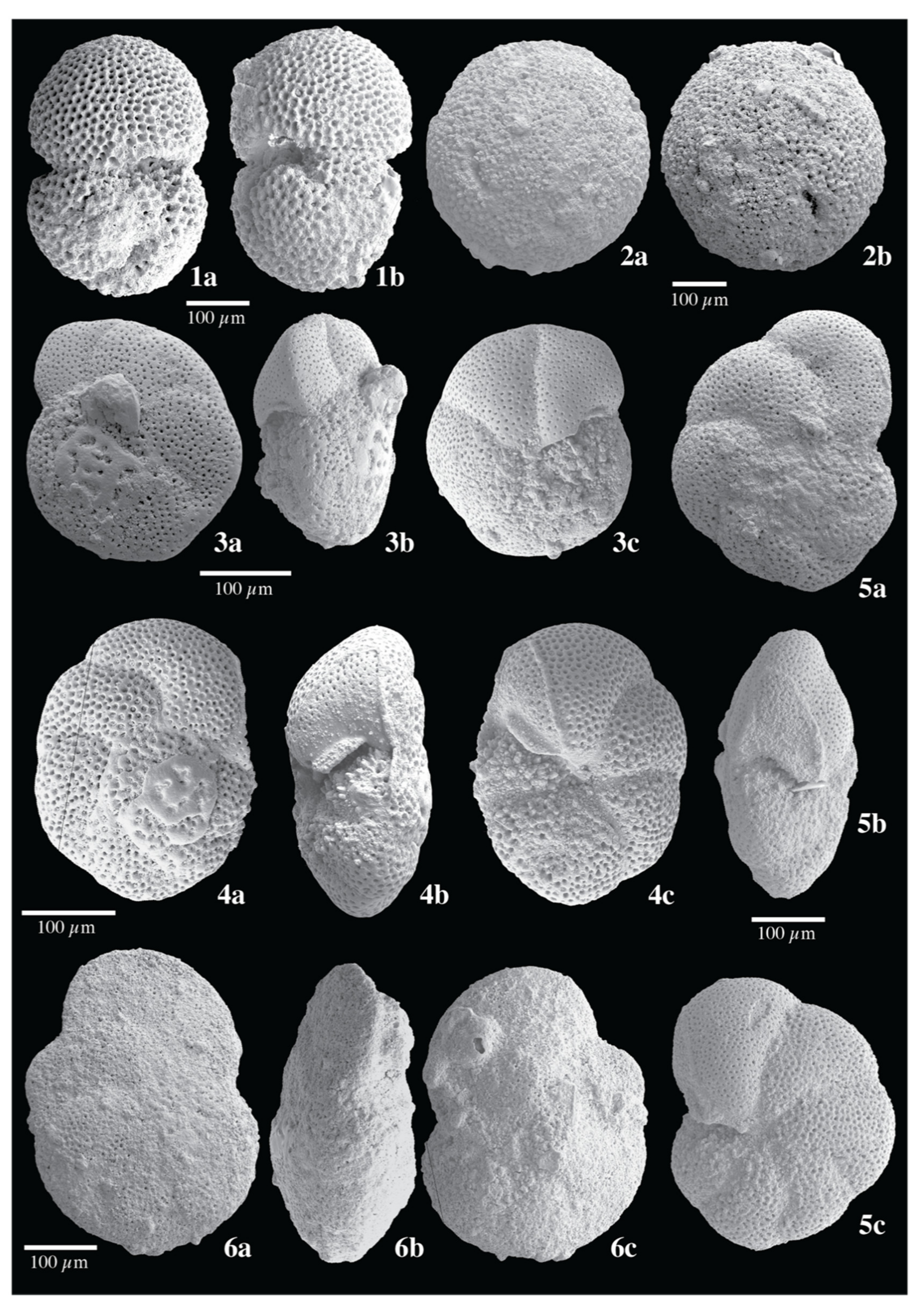{kind=link}
| Name | Plotcodes | Age | Top Depth | Bottom Depth | Data Origin |
|---|---|---|---|---|---|
| FO Emiliania huxleyi | bE_h | 0.29 | 3.4 | 8.8 | Shipboard / Postcruise data (this article) |
| LO Pseudoemiliania lacunosa | tP_l | 0.43 | 3.4 | 8.8 | Shipboard / Postcruise data (this article) |
| LO Discoaster brouweri | tD_b | 1.93 | 31.2 | 40.5 | Shipboard / Postcruise data (this article) |
| LO Coronocyclus nitescens | tC_n | 12.45 | 419.2 | 429 | Shipboard / Postcruise data (this article) |
| LCO Cyclicargolithus floridanus | tC_f | 13.33 | 473.3 | 497.9 | Shipboard / Postcruise data (this article) |
| LO Sphenolithus heteromorphus | tS_h | 13.6 | 486.9 | 497.9 | Shipboard / Postcruise data (this article) |
| FO Sphenolithus heteromorphus | bS_h | 17.65 | 624 | 624.7 | Shipboard / Postcruise data (this article) |
| LO Sphenolithus belemnos | tS_b | 17.94 | 624.7 | 643.7 | Shipboard / Postcruise data (this article) |
| FO Sphenolithus belemnos | bS_b | 19.01 | 644.7 | 653.7 | Shipboard / Postcruise data (this article) |
| LO Sphenolithus ciperoensis | tS_c | 24.3 | 806.4 | 807.3 | Shipboard / Postcruise data (this article) |
| LO Sphenolithus distentus | tS_d | 26.8 | 846.7 | 880 | Shipboard / Postcruise data (this article) |
| LO Globigerinoides ruber pink | tG_rp | 0.12 | 0.05 | 3.35 | Shipboard |
| LO Globigerinoidesella fistulosa | tG_f | 1.88 | 3.4 | 8.8 | Shipboard |
| LO Globorotalia limbata | tG_l | 2.39 | 8.8 | 21.6 | Shipboard |
| FO Globigerinoidesella fistulosa | bG_f | 3.33 | 40.6 | 50.1 | Shipboard |
| FO Sphaeroidinella dehiscens | bS_d | 5.53 | 50.1 | 59.4 | Shipboard |
| FO Neogloboquadrina acostaensis | bN_a | 9.89 | 59.4 | 69 | Shipboard |
| LO Paragloborotalia mayeri | tP_m | 10.54 | 69 | 75.7 | Shipboard |
| FO Fohsella fohsi | bF_f | 13.4 | 466.6 | 474.8 | Shipboard |
| FO Orbulina suturalis | bO_s | 15.1 | 534.6 | 551.2 | Shipboard |
| LO Paragloborotalia kugleri | tP_k | 21.12 | 694.7 | 711.1 | Shipboard |
| FO Paragloborotalia kugleri | bP_k | 22.96 | 769.7 | 779.4 | Shipboard |
| base Zone N23 | bN23 | 0.2 | 3.4 | 12.9 | Postcruise data (this article) |
| base Zone PT1 | bPt1 | 1.88 | 31.9 | 41.4 | Postcruise data (this article) |
| base Zone Pl5-6 | bPL5-6 | 3.47 | 50.9 | 60.4 | Postcruise data (this article) |
| base Zone M13 | tM13 | 9.89 | 50.9 | 60.4 | Postcruise data (this article) |
| base Zone M10 | bM10 | 11.93 | 192.4 | 197.1 | Postcruise data (this article) |
| base Zone M9 | bM9 | 13.4 | 476.3 | 486 | Postcruise data (this article) |
| base Zone M8 | bM8 | 13.77 | 505.5 | 515.2 | Postcruise data (this article) |
| base Zone M7 | bM7 | 14.06 | 524.9 | 534.6 | Postcruise data (this article) |
| base Zone M6 | bM6 | 15.12 | 534.6 | 544.3 | Postcruise data (this article) |
| base Zone M5b | bM5b | 16.27 | 573.4 | 583.1 | Postcruise data (this article) |
| base Zone M5a | bM5a | 16.39 | 641.3 | 651 | Postcruise data (this article) |
| base Zone M2 | bM2 | 21.12 | 699.5 | 709.2 | Postcruise data (this article) |
| base Zone M1 | bM1b | 22.5 | 757.7 | 767.4 | Postcruise data (this article) |
| base Zone O7 | bO7 | 25.4 | 835.3 | 845 | Postcruise data (this article) |
| top Zone O5 | bO5 | 27.5 | 865 | 874.7 | Postcruise data (this article) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spezzaferri, S.; Young, J.; Stainbank, S.; Coletti, G.; Kroon, D. Improved Planktonic, Benthic Foraminiferal and Nannofossil Biostratigraphy Aids the Interpretation of the Evolution at Hole U1468A: IODP Expedition 359, the Maldives. Geosciences 2022, 12, 239. https://0-doi-org.brum.beds.ac.uk/10.3390/geosciences12060239
Spezzaferri S, Young J, Stainbank S, Coletti G, Kroon D. Improved Planktonic, Benthic Foraminiferal and Nannofossil Biostratigraphy Aids the Interpretation of the Evolution at Hole U1468A: IODP Expedition 359, the Maldives. Geosciences. 2022; 12(6):239. https://0-doi-org.brum.beds.ac.uk/10.3390/geosciences12060239
Chicago/Turabian StyleSpezzaferri, Silvia, Jeremy Young, Stephanie Stainbank, Giovanni Coletti, and Dick Kroon. 2022. "Improved Planktonic, Benthic Foraminiferal and Nannofossil Biostratigraphy Aids the Interpretation of the Evolution at Hole U1468A: IODP Expedition 359, the Maldives" Geosciences 12, no. 6: 239. https://0-doi-org.brum.beds.ac.uk/10.3390/geosciences12060239