
Combined Effects of Fludarabine and Interferon Alpha on Autophagy Regulation Define the Phase of Cell Survival and Promotes Responses in LLC-MK2 and K562 Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Chemicals and Antibodies
2.3. MTT Assay to Determine the Cytotoxic Effect of Fdb
2.4. Biochemical and Immunohistochemical Analysis for Autophagy and Immune-Signaling Response
2.4.1. Western Blot Analysis
2.4.2. Immunohistochemical Analysis
3. Results
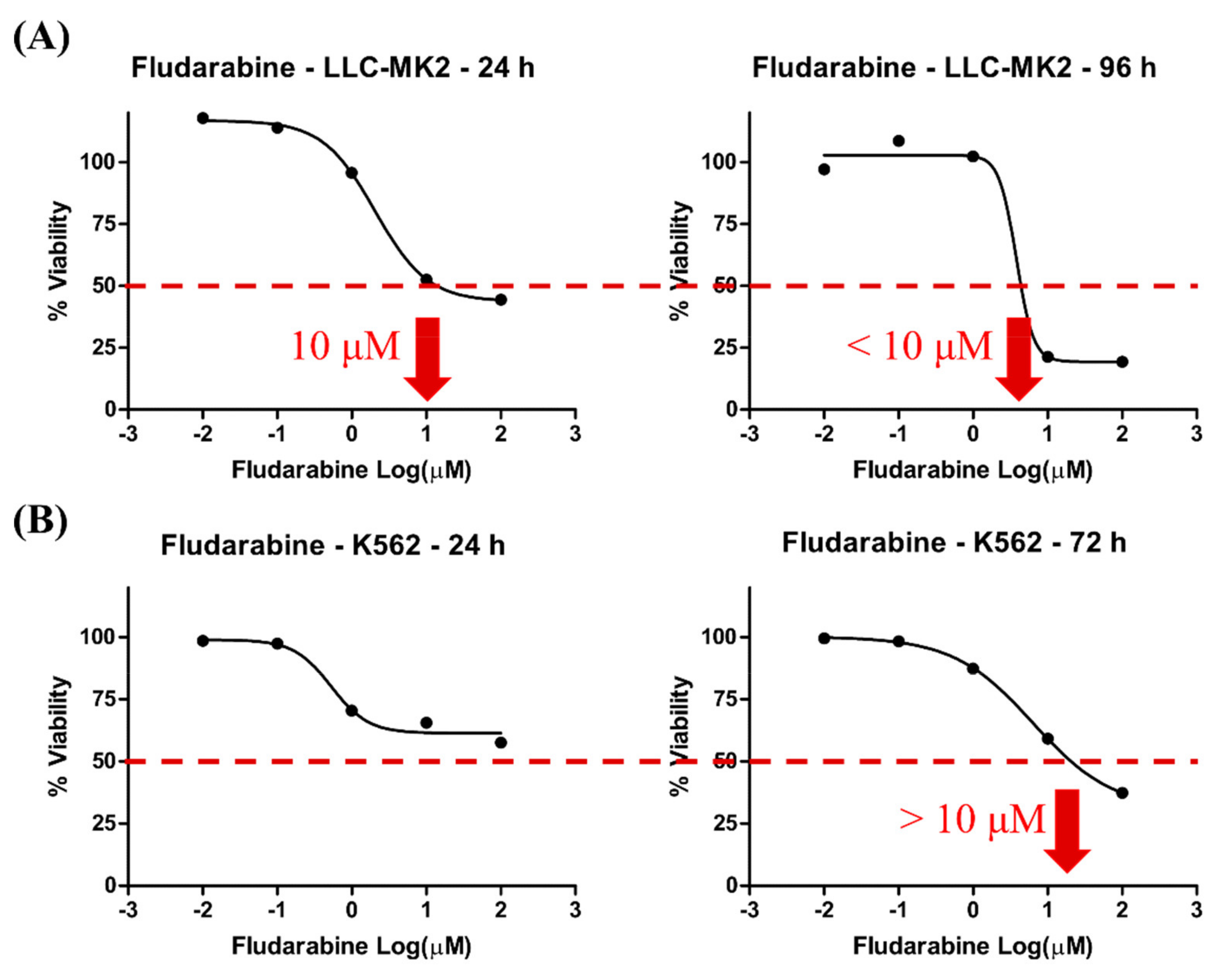
3.1. Cell Viability Induced by Fdb
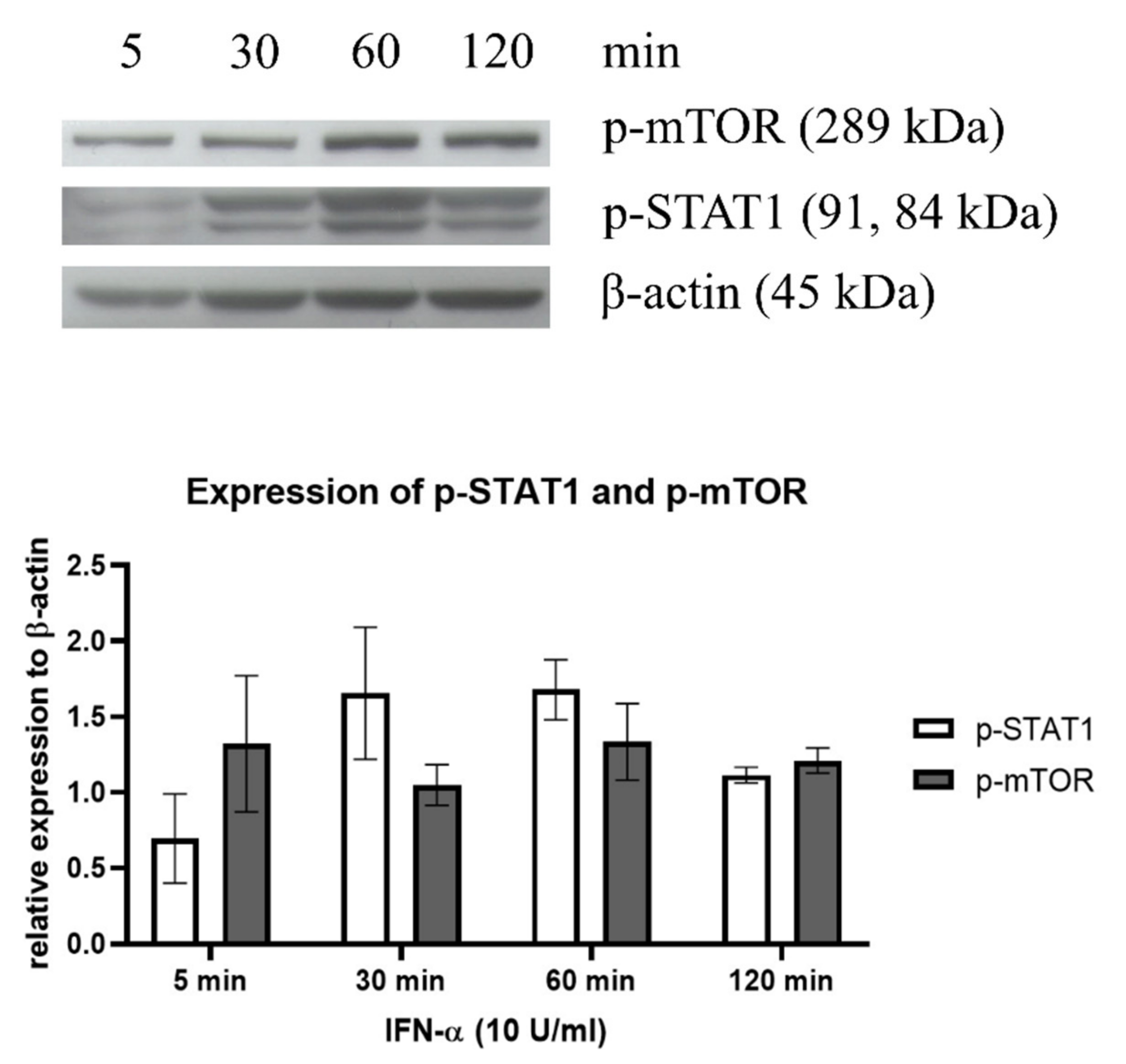
3.2. IFN-α Induced STAT1 Signaling in Both LLC-MK2 and K562 Cells That Rendered IFNR
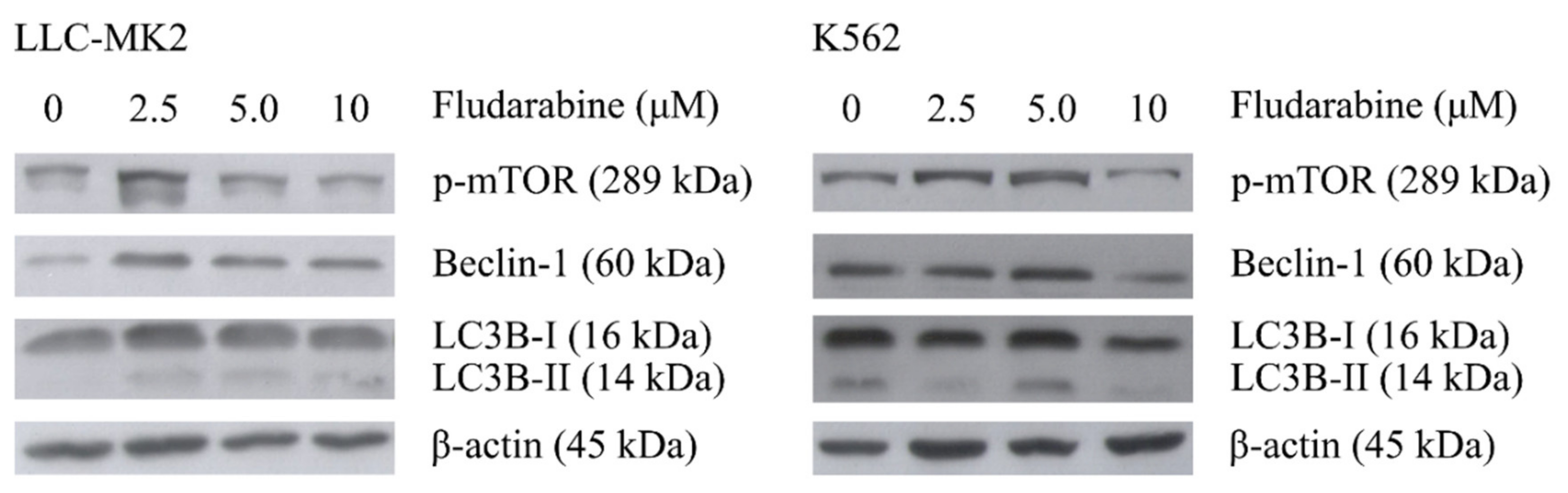
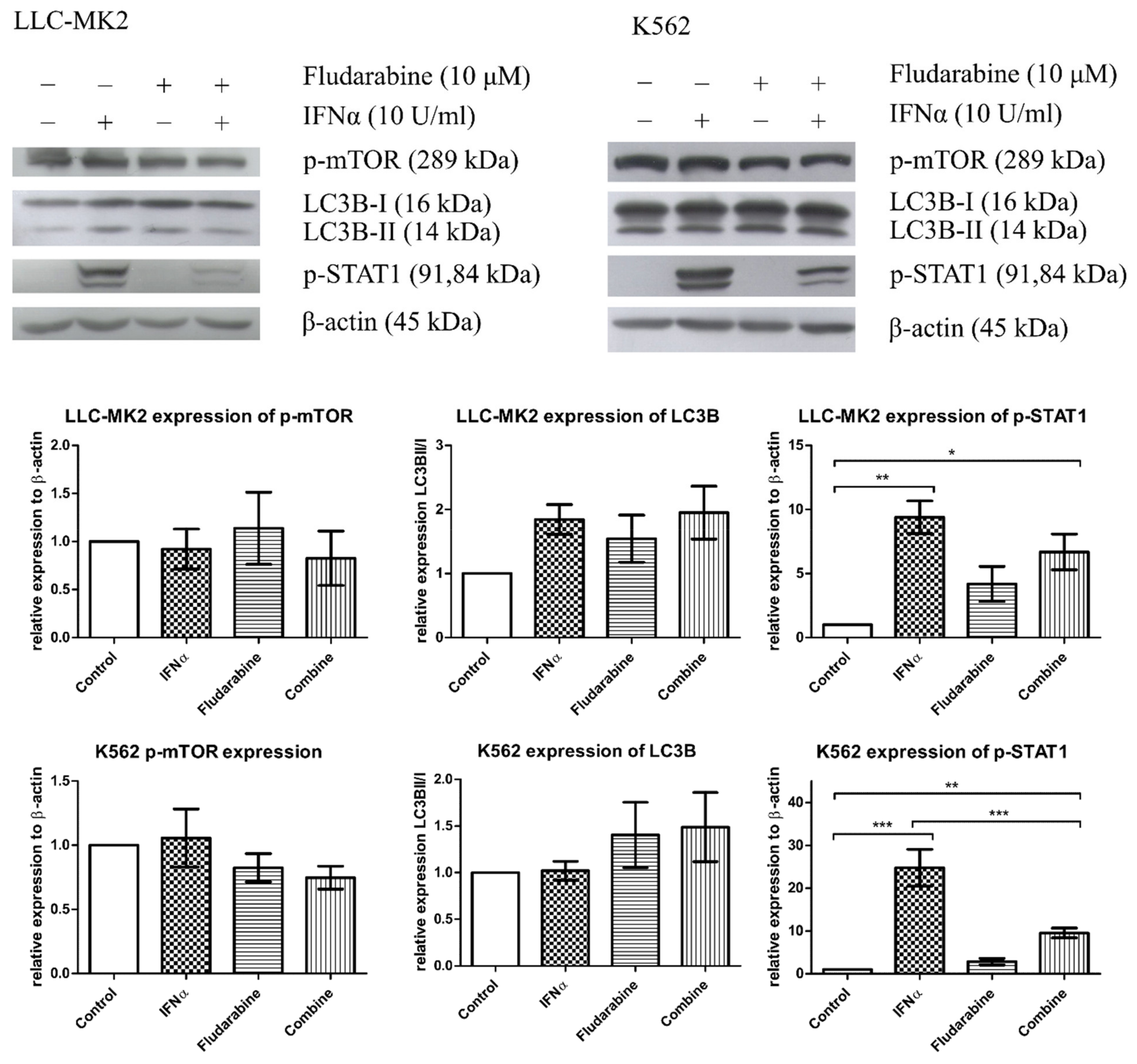
3.3. The Effect of Fdb Treatment on IFN-α-Exposed LLC-MK2 and K562 Cells
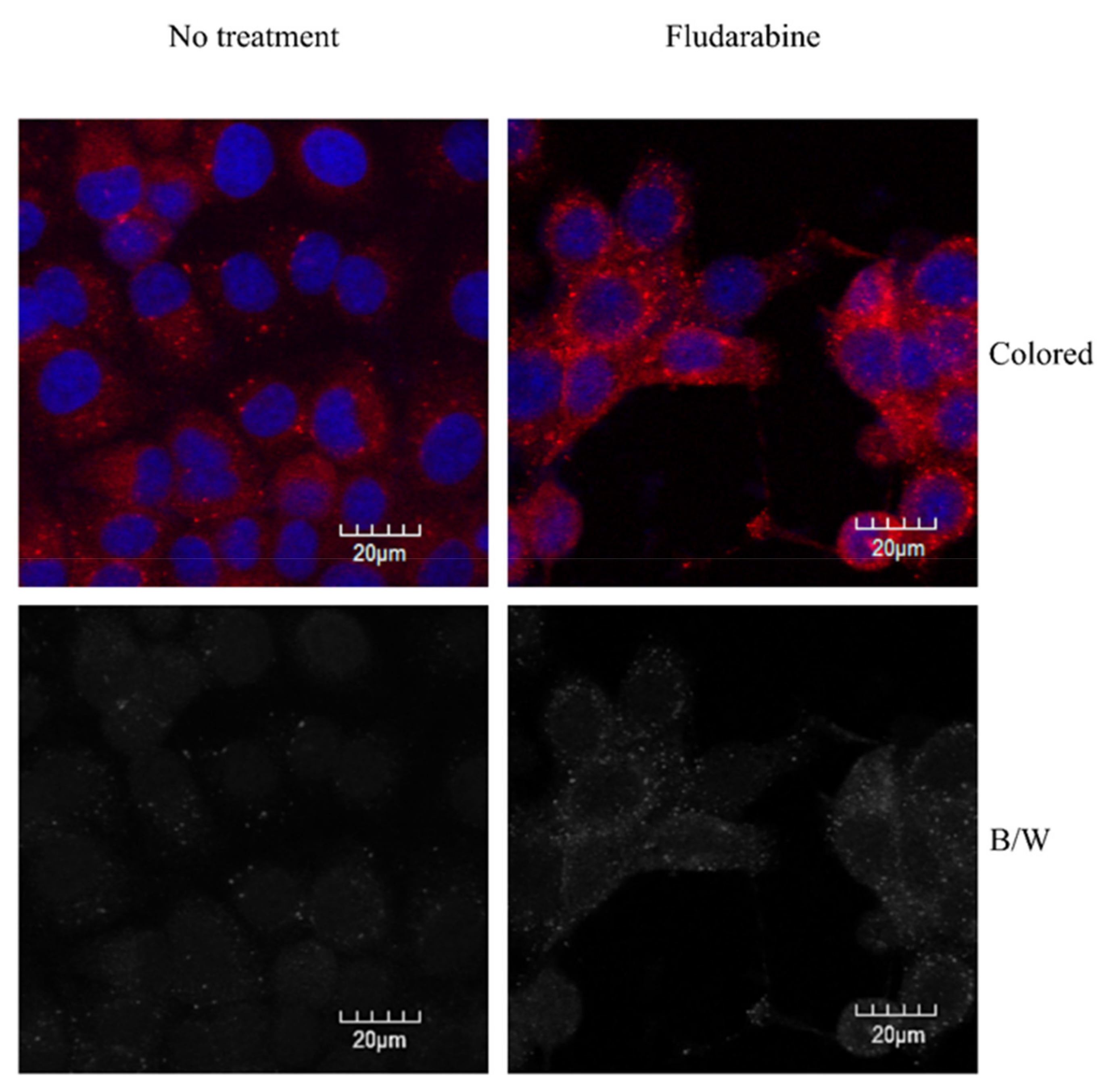
3.4. Autophagy Phenomena in Live Cells after Treatment with IFN-α, Fdb, and IFN-α + Fdb
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Yan, X.; Zhou, R.; Ma, Z. Autophagy—Cell Survival and Death. In Autophagy: Biology and Diseases: Basic Science; Qin, Z.-H., Ed.; Springer: Singapore, 2019; pp. 667–696. [Google Scholar]
- Qian, M.; Fang, X.; Wang, X. Autophagy and inflammation. Clin. Transl. Med. 2017, 6, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Wang, M.-L.; Zhao, J. Crosstalk between autophagy and type I interferon responses in innate antiviral immunity. Viruses 2019, 11, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, C.W.; Lee, S.H. The roles of autophagy in cancer. Int. J. Mol. Med. 2018, 19, 3466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keating, M.; Kantarjian, H.; Talpaz, M.; Redman, J.; Koller, C.; Barlogie, B.; Velasquez, W.; Plunkett, W.; Freireich, E.J.; McCredie, K.B. Fludarabine: A new agent with major activity against chronic lymphocytic leukemia. Blood 1989, 74, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Shevrin, D.H.; Lad, T.; Kilton, L.; Cobleigh, M.; Blough, R.; Weidner, L.; Vogelzang, N. Phase II trial of fludarabine phosphate in advanced renal cell carcinoma: An Illinois Cancer Council study. Investig. New Drugs 1989, 7, 251–253. [Google Scholar] [CrossRef]
- Von Hoff, D.D.; Green, S.; Surwit, E.A.; Hannigan, E.V.; Alberts, D.S. Phase II Study of Fludarabine Phosphate (NSC 312887) in Patients with Advanced Cervical Cancer: A Southwest Oncology Group Study. Am. J. Clin. Oncol. 1990, 13, 433–435. [Google Scholar] [CrossRef]
- Chun, H.G.; Leyland-Jones, B.; Cheson, B.D. Fludarabine phosphate: A synthetic purine antimetabolite with significant activity against lymphoid malignancies. J. Clin. Oncol. 1991, 9, 175–188. [Google Scholar] [CrossRef]
- Von Hoff, D.D.; Green, S.; Alberts, D.S.; Stock-Novack, D.L.; Surwit, E.A.; Miller, T.P.; Stephens, R.L. Phase II Study of Fludarabine Phosphate (NSC-312887) in Patients with Advanced Endometrial Cancer: Southwest Oncology Group Study. Am. J. Clin. Oncol. 1991, 14, 193–194. [Google Scholar] [CrossRef]
- Rodriguez, G. Fludarabine phosphate. Investig. New Drugs 1994, 12, 75–92. [Google Scholar] [CrossRef]
- Gandhi, V.; Plunkett, W. Cellular and clinical pharmacology of fludarabine. Clin. Pharmacokinet. 2002, 41, 93–103. [Google Scholar] [CrossRef]
- Montillo, M.; Ricci, F.; Tedeschi, A. Fludarabine in the treatment of chronic lymphocytic leukemia: A review. Ther. Clin. Risk Manag. 2009, 5, 187–207. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, C.M.; Bussi, C.; Arroyo, D.S.; Sastre, D.; Heller, V.; Stanganelli, C.; Slavutsky, I.; Iribarren, P. Effects of rapamycin in combination with fludarabine on primary chronic lymphocytic leukemia cells. Leuk. Lymphoma 2019, 60, 1299–1303. [Google Scholar] [CrossRef]
- Wang, X.-X.; Gao, Y.; Jin, J.; Cao, J.N.; Feng, J.F.; Wang, H.Q.; Zhang, H.L.; Cai, Q.Q.; Li, Z.M.; Jiang, W.Q.; et al. Bortezomib in combination with fludarabine plus cyclophosphamide for patients with relapsed or refractory mantle-cell lymphoma: Results of the LYM-4003 study. Ann. Hematol. 2021, 100, 2961–2968. [Google Scholar] [CrossRef]
- Gao, C.; Wen, C.; Li, Z.; Lin, S.; Gao, S.; Ding, H.; Zou, P.; Xing, Z.; Yu, Y. Fludarabine Inhibits Infection of Zika Virus, SFTS Phlebovirus, and Enterovirus A71. Viruses 2021, 13, 774. [Google Scholar] [CrossRef]
- Xiu, H.; Gong, J.; Huang, T.; Peng, Y.; Bai, S.; Xiong, G.; Zhang, S.; Huang, H.; Cai, Z.; Zhang, G. Fludarabine inhibits type I interferon-induced expression of the SARS-CoV-2 receptor angiotensin-converting enzyme 2. Cell Mol. Immunol. 2021, 18, 1829–1831. [Google Scholar] [CrossRef]
- Kirkwood, J. Cancer immunotherapy: The interferon-α experience. In Seminars in Oncology; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Lesinski, G.B.; Anghelina, M.; Zimmerer, J.; Bakalakos, T.; Badgwell, B.; Parihar, R.; Hu, Y.; Becknell, B.; Abood, G.; Chaudhury, A.R.; et al. The antitumor effects of IFN-α are abrogated in a STAT1-deficient mouse. J. Clin. Investig. 2003, 112, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Baron, S.; Tyring, S.K.; Fleischmann, W.R.; Coppenhaver, D.H.; Niesel, D.W.; Klimpel, G.R.; Stanton, G.J.; Hughes, T.K. The interferons: Mechanisms of action and clinical applications. JAMA 1991, 266, 1375–1383. [Google Scholar] [CrossRef]
- Chang, E.; Boyd, A.; Nelson, C.C.; Crowley, D.; Law, T.; Keough, K.M.; Folkman, J.; Ezekowitz, R.A.B.; Castle, V.P. Successful treatment of infantile hemangiomas with interferon-alpha-2b. J. Pediatr. Hematol. Oncol. 1997, 19, 237–244. [Google Scholar] [CrossRef]
- Renault, P.F.; Hoofnagle, J.H. Side effects of alpha interferon. In Seminars in Liver Disease; Thieme Medical Publishers, Inc.: Stuttgart, Germany, 1989. [Google Scholar]
- Zhu, S.; Cao, L.; Yu, Y.; Yang, L.; Yang, M.; Liu, K.; Huang, J.; Kang, R.; Livesey, K.M.; Tang, D. Inhibiting autophagy potentiates the anticancer activity of IFN1@/IFNα in chronic myeloid leukemia cells. Autophagy 2013, 9, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Aryan, M.; Qian, S.; Cabrera, R.; Liu, X. A Focused Review on Recent Advances in the Diagnosis and Treatment of Viral Hepatitis. Gastroenterol. Res. 2021, 14, 139–156. [Google Scholar] [CrossRef]
- Van der Sluis, R.M.; Zerbato, J.M.; Rhodes, J.W.; Pascoe, R.D.; Solomon, A.; Kumar, N.A.; Dantanarayana, A.I.; Tennakoon, S.; Dufloo, J.; McMahon, J.; et al. Diverse effects of interferon alpha on the establishment and reversal of HIV latency. PLoS Pathog. 2020, 16, e1008151. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.M.; Lennox, J.L.; Mulligan, M.M.; Loring, D.; Zetterberg, H.; Blennow, K.; Kessing, C.; Koneru, R.; Easley, K.; Tyor, W.R. Cerebrospinal fluid interferon alpha levels correlate with neurocognitive impairment in ambulatory HIV-Infected individuals. J. Neurovirol. 2017, 23, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grubczak, K.; Grzeszczuk, A.; Groth, M.; Hryniewicz, A.; Kretowska-Grunwald, A.; Flisiak, R.; Moniuszko, M. Effects of Pegylated Interferon Alpha and Ribavirin (pegIFN-α/RBV) Therapeutic Approach on Regulatory T Cells in HCV-Monoinfected and HCV/HIV-Coinfected Patients. Viruses 2021, 13, 1448. [Google Scholar] [CrossRef] [PubMed]
- Kwaa, A.K.; Talana, C.A.; Blankson, J.N. Interferon alpha enhances NK cell function and the suppressive capacity of HIV-specific CD8+ T cells. J. Virol. 2019, 93, e01541-18. [Google Scholar] [CrossRef] [Green Version]
- Nakhlband, A.; Fakhari, A.; Azizi, H. Interferon-alpha position in combating with COVID-19: A systematic review. J. Med. Virol. 2021, 93, 5277–5284. [Google Scholar] [CrossRef]
- Levine, B.; Mizushima, N.; Virgin, H.W. Autophagy in immunity and inflammation. Nature 2011, 469, 323–335. [Google Scholar] [CrossRef]
- Levine, B.; Deretic, V. Unveiling the roles of autophagy in innate and adaptive immunity. Nat. Rev. Immunol. 2007, 7, 767–777. [Google Scholar] [CrossRef]
- Münz, C. Enhancing immunity through autophagy. Annu. Rev. Immunol. 2009, 27, 423–449. [Google Scholar] [CrossRef]
- Sharma, A.; Singh, K.; Mazumder, S.; Hill, B.T.; Kalaycio, M.; Almasan, A. BECN1 and BIM interactions with MCL-1 determine fludarabine resistance in leukemic B cells. Cell Death Dis. 2013, 4, e628. [Google Scholar] [CrossRef] [Green Version]
- Mahoney, E.; Byrd, J.C.; Johnson, A.J. Autophagy and ER stress play an essential role in the mechanism of action and drug resistance of the cyclin-dependent kinase inhibitor flavopiridol. Autophagy 2013, 9, 434–435. [Google Scholar] [CrossRef]
- Mahoney, E.; Johnson, A.J.; Wagner, A.J.; Hessler, J.; Gupta, S.V.; Herman, S.E.; Andritsos, L.; Jones, J.A.; Flynn, J.M.; Lucas, D.M.; et al. Autophagy Is a Relevant Cellular Process In CLL and Contributes to Drug Resistance of Flavopiridol. Blood 2010, 116, 691. [Google Scholar] [CrossRef]
- O’Shea, J.J.; Schwartz, D.M.; Villarino, A.V.; Gadina, M.; McInnes, I.B.; Laurence, A. The JAK-STAT pathway: Impact on human disease and therapeutic intervention. Annu. Rev. Med. 2015, 66, 311–328. [Google Scholar] [CrossRef] [Green Version]
- Hervas-Stubbs, S.; Perez-Gracia, J.L.; Rouzaut, A.; Sanmamed, M.F.; Le Bon, A.; Melero, I. Direct effects of type I interferons on cells of the immune system. Clin. Cancer Res. 2011, 17, 2619–2627. [Google Scholar] [CrossRef] [Green Version]
- Sadler, A.J.; Williams, B.R. Interferon-inducible antiviral effectors. Nat. Rev. Immunol. 2008, 8, 559–568. [Google Scholar] [CrossRef]
- Schmeisser, H.; Bekisz, J.; Zoon, K.C. New function of type I IFN: Induction of autophagy. J. Interf. Cytokine Res. 2014, 34, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Datan, E.; Roy, S.G.; Germain, G.; Zali, N.; McLean, J.E.; Golshan, G.; Harbajan, S.; Lockshin, R.A.; Zakeri, Z. Dengue-induced autophagy, virus replication and protection from cell death require ER stress (PERK) pathway activation. Cell Death Dis. 2016, 7, e2127. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Rocha, H.; Gomez-Gutierrez, J.G.; Garcia-Garcia, A.; Rao, X.-M.; Chen, L.; McMasters, K.M.; Zhou, H.S. Adenoviruses induce autophagy to promote virus replication and oncolysis. Virology 2011, 416, 9–15. [Google Scholar] [CrossRef] [Green Version]
- McLean, J.E.; Wudzinska, A.; Datan, E.; Quaglino, D.; Zakeri, Z. Flavivirus NS4A-induced autophagy protects cells against death and enhances virus replication. J. Biol. Chem. 2011, 286, 22147–22159. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowornruangrit, P.; Kumkate, S.; Sirigulpanit, W.; Leardkamolkarn, V. Combined Effects of Fludarabine and Interferon Alpha on Autophagy Regulation Define the Phase of Cell Survival and Promotes Responses in LLC-MK2 and K562 Cells. Med. Sci. 2022, 10, 20. https://0-doi-org.brum.beds.ac.uk/10.3390/medsci10010020
Bowornruangrit P, Kumkate S, Sirigulpanit W, Leardkamolkarn V. Combined Effects of Fludarabine and Interferon Alpha on Autophagy Regulation Define the Phase of Cell Survival and Promotes Responses in LLC-MK2 and K562 Cells. Medical Sciences. 2022; 10(1):20. https://0-doi-org.brum.beds.ac.uk/10.3390/medsci10010020
Chicago/Turabian StyleBowornruangrit, Pathompong, Supeecha Kumkate, Wipawan Sirigulpanit, and Vijittra Leardkamolkarn. 2022. "Combined Effects of Fludarabine and Interferon Alpha on Autophagy Regulation Define the Phase of Cell Survival and Promotes Responses in LLC-MK2 and K562 Cells" Medical Sciences 10, no. 1: 20. https://0-doi-org.brum.beds.ac.uk/10.3390/medsci10010020