
Mercury Accumulation, and the Mercury-PCB-Sex Interaction, in Lake Whitefish (Coregonus clupeaformis)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Methods
2.2. Mercury Determinations
2.3. Data Analyses for Hg and MeHg Concentrations
2.4. Bioenergetics Modeling
2.5. Comparison of PCBs with Hg
3. Results
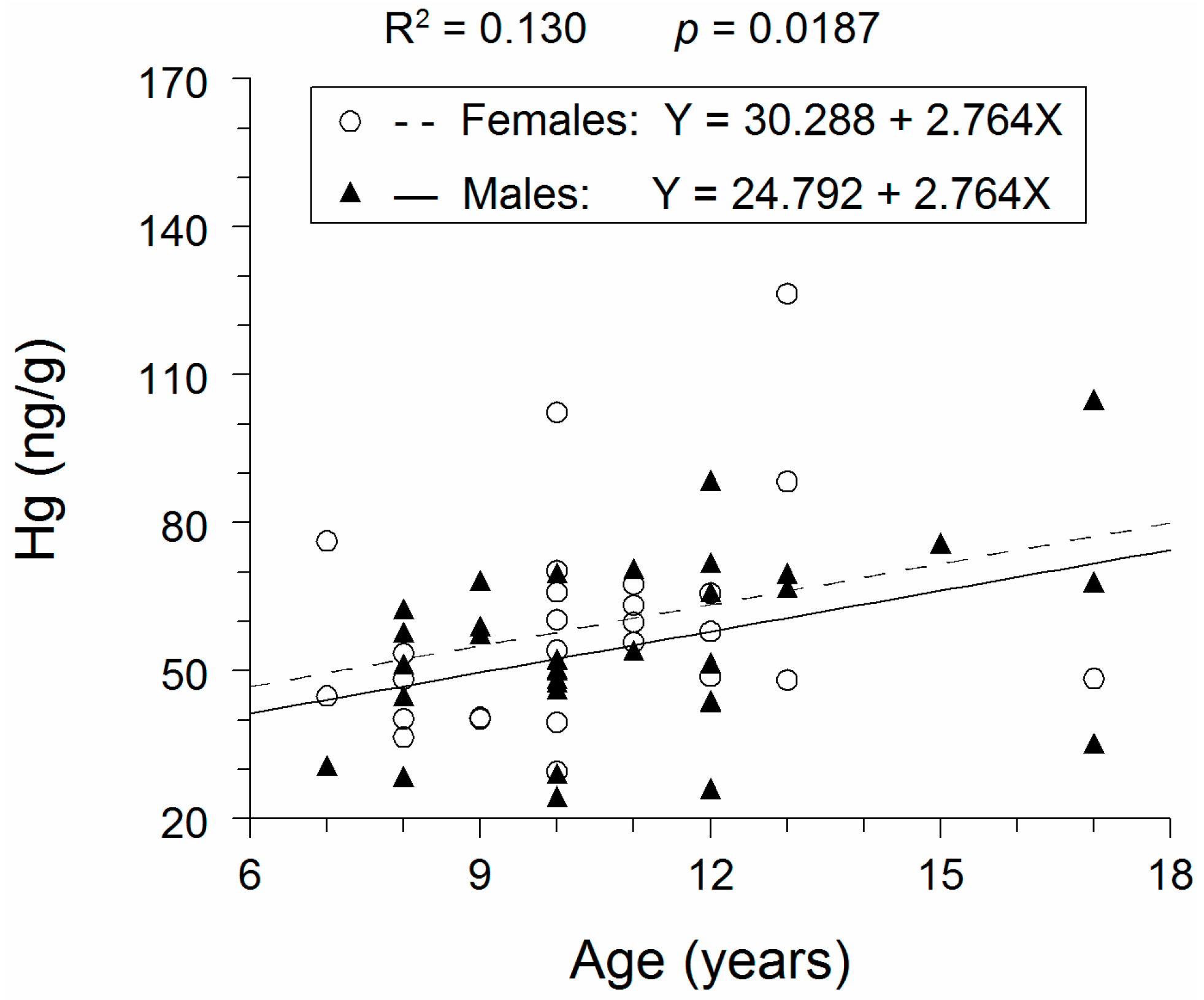
3.1. Hg Concentrations
3.2. MeHg Concentrations
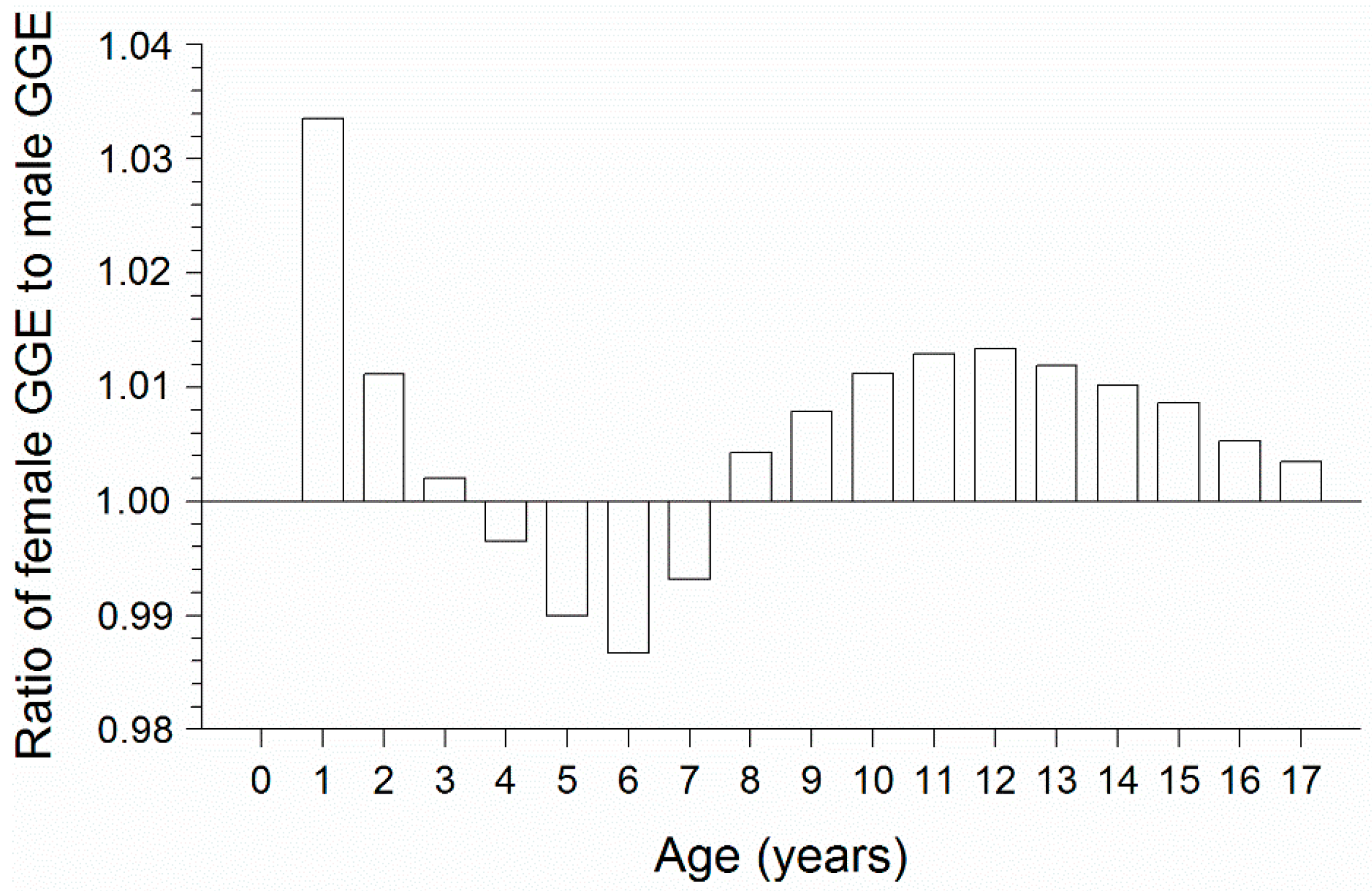
3.3. Bioenergetics Modeling
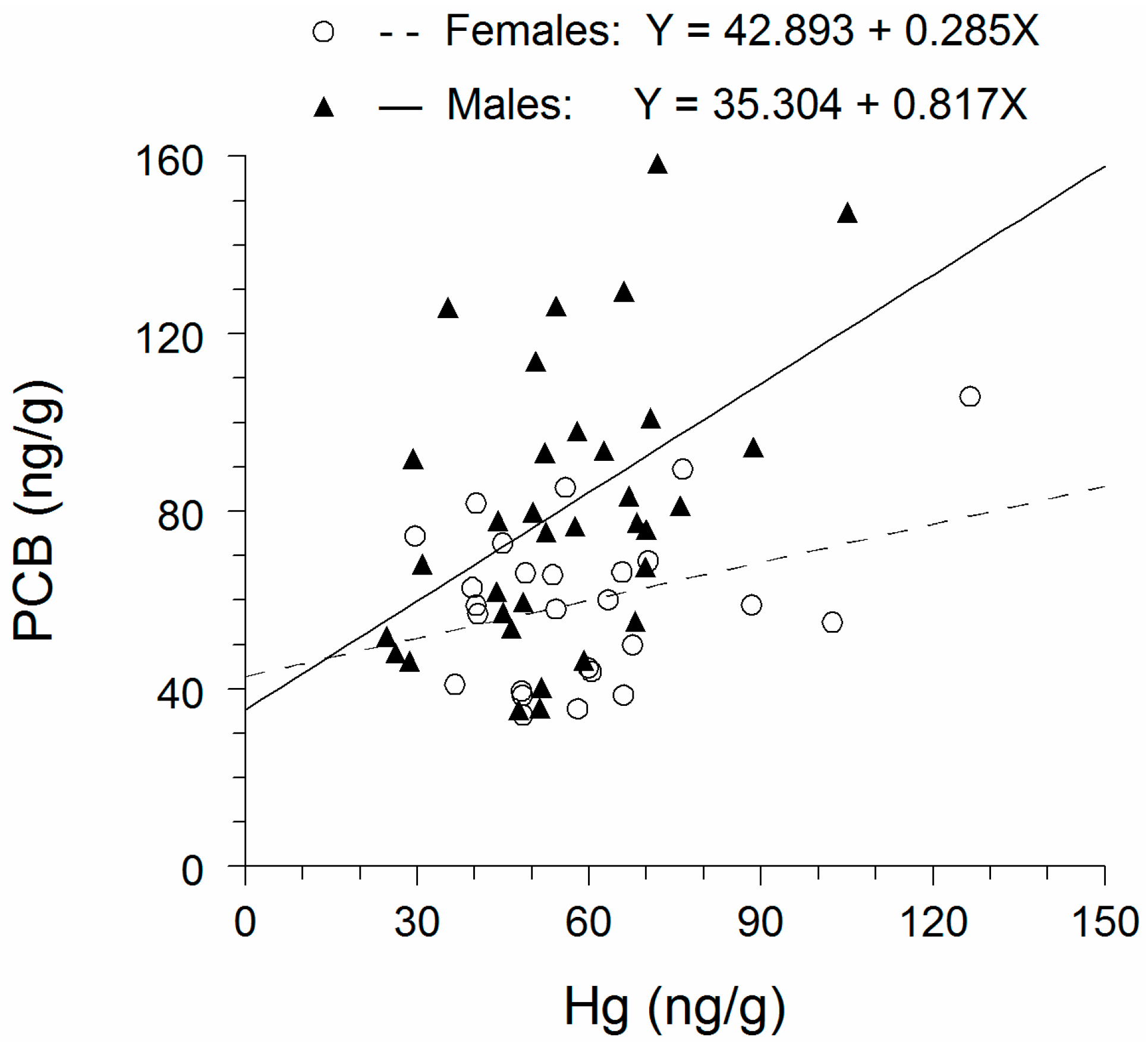
3.4. Comparison of PCBs with Hg
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brenden, T.O.; Brown, R.W.; Ebener, M.P.; Reid, K.; Newcomb, T.J. Great Lakes commercial fisheries: Historical overview and prognoses for the future. In Great Lakes Fisheries Policy and Management: A Binational Perspective, 2nd ed.; Taylor, W.W., Lynch, A.J., Leonard, N.J., Eds.; Michigan State University Press: East Lansing, MI, USA, 2013; pp. 339–397. [Google Scholar]
- Ebener, M.P.; Kinnunen, R.E.; Schneeberger, P.J.; Mohr, L.C.; Hoyle, J.A.; Peeters, P. Management of commercial fisheries for lake whitefish in the Laurentian Great Lakes of North America. In International Governance of Fisheries Ecosystems: Learning from the Past, Finding Solutions for the Future; Schechter, M.G., Taylor, W.W., Leonard, N.J., Eds.; American Fisheries Society: Bethesda, MD, USA, 2008; pp. 99–143. [Google Scholar]
- Kinietz, W.V. The Indians of the Western Great Lakes, 1615–1760; University of Michigan Press: Ann Arbor, MI, USA, 1965. [Google Scholar]
- Pothoven, S.A.; Madenjian, C.P. Changes in consumption by alewives and lake whitefish after dreissenid mussel invasions in Lakes Michigan and Huron. N. Am. J. Fish. Manag. 2008, 28, 308–320. [Google Scholar]
- Pothoven, S.A.; Madenjian, C.P. Increased piscivory by lake whitefish in Lake Huron. N. Am. J. Fish. Manag. 2013, 33, 1194–1202. [Google Scholar] [CrossRef]
- Wiener, J.G.; Krabbenhoft, D.P.; Heinz, G.H.; Scheuhammer, A.M. Ecotoxicology of mercury. In Handbook of Ecotoxicology, 2nd ed.; Hoffman, D.J., Rattner, B.A., Burton, G.A., Jr., Cairns, J., Jr., Eds.; CRC Press: Boca Raton, FL, USA, 2003; pp. 407–461. [Google Scholar]
- WHO (World Health Organization). Ten Chemicals of Major Public Health Concern. 2010. Available online: http://www.who.int/ipcs/assessment/public_health/chemicals_phc/en/ (accessed on 3 November 2015).
- Clarkson, T.W.; Magos, L.; Myers, G.J. The toxicology of mercury—Current exposures and clinical manifestations. N. Engl. J. Med. 2003, 349, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Scheuhammer, A.M.; Meyer, M.W.; Sandheinrich, M.B.; Murray, M.W. Effects of environmental methylmercury on the health of wild birds, mammals, and fish. Ambio 2007, 36, 12–18. [Google Scholar] [CrossRef]
- Sandheinrich, M.B.; Bhavsar, S.P.; Bodaly, R.A.; Drevnick, P.E.; Paul, E.A. Ecological risk of methylmercury to piscivorous fish of the Great Lakes region. Ecotoxicology 2011, 20, 1577–1587. [Google Scholar] [CrossRef] [PubMed]
- Wiener, J.G.; Evers, D.C.; Gay, D.A.; Morrison, H.A.; Williams, K.A. Mercury contamination in the Laurentian Great Lakes region: Introduction and overview. Environ. Pollut. 2012, 161, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Somers, E.C.; Ganser, M.A.; Warren, J.S.; Basu, N.; Wang, L.; Zick, S.M.; Park, S.K. Mercury exposure and antinuclear antibodies among females of reproductive age in the United States: NHANES. Environ. Health Perspect. 2015, 123, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Wren, C.D. A review of metal accumulation and toxicity in wild mammals. I. Mercury. Environ. Res. 1986, 40, 210–244. [Google Scholar] [CrossRef]
- Ginsberg, G.L.; Toal, B.E. Development of a single-meal fish consumption advisory for methyl mercury. Risk Anal. 2000, 20, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.; Gochfeld, M.; Powers, C.W.; Waishwell, L.; Warren, C.; Goldstein, B.D. Science, policy, stakeholders, and fish consumption advisories: Developing a fish fact sheet for the Savannah River. Environ. Manag. 2001, 27, 501–514. [Google Scholar] [CrossRef]
- Pastorok, R.A.; Bartell, S.M.; Ferson, S.; Ginzburg, L.R. Ecological Modeling in Risk Assessment: Chemical Effects on Populations, Ecosystems, and Landscapes; Lewis: Boca Raton, FL, USA, 2002. [Google Scholar]
- McClain, W.C.; Chumchal, M.M.; Drenner, R.W.; Newland, L.W. Mercury concentrations in fish from Lake Meredith, Texas: Implications for the issuance of fish consumption advisories. Environ. Monit. Assess. 2006, 123, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Rooker, J.R.; Gill, G.A.; Turner, J.P. Bioaccumulation of mercury in pelagic fishes from the northern Gulf of Mexico. Can. J. Fish. Aquat. Sci. 2007, 64, 458–469. [Google Scholar] [CrossRef]
- Mason, R.P.; Sullivan, K.A. Mercury in Lake Michigan. Environ. Sci. Technol. 1997, 31, 942–947. [Google Scholar] [CrossRef]
- Lepak, R.F.; Yin, R.; Krabbenhoft, D.P.; Ogorek, J.M.; DeWild, J.F.; Holsen, T.M.; Hurley, J.P. Use of stable isotope signatures to determine mercury sources in the Great Lakes. Environ. Sci. Technol. Lett. 2015, 2, 335–341. [Google Scholar] [CrossRef]
- Marvin, C.; Painter, S.; Rossmann, R. Spatial and temporal patterns in mercury contamination in sediments of the Laurentian Great Lakes. Environ. Res. 2004, 95, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Suchanek, T.H.; Mullen, L.H.; Lamphere, B.A.; Richerson, P.J.; Woodmansee, C.E.; Slotton, D.G.; Harner, E.J.; Woodward, L.A. Redistribution of mercury from contaminated lake sediments of Clear Lake, California. Water Air Soil Pollut. 1998, 104, 77–102. [Google Scholar] [CrossRef]
- Madenjian, C.P.; Stapanian, M.A.; Cott, P.A.; Krabbenhoft, D.P.; Edwards, W.H.; Ogilvie, L.M.; Mychek-Londer, J.G.; DeWild, J.F. Females exceed males in mercury concentrations of burbot Lota lota. Arch. Environ. Contam. Toxicol. 2015, 68, 678–688. [Google Scholar] [CrossRef] [PubMed]
- Trudel, M.; Tremblay, A.; Schetagne, R.; Rasmussen, J.B. Estimating food consumption rates of fish using a mercury mass balance model. Can. J. Fish. Aquat. Sci. 2000, 57, 414–428. [Google Scholar] [CrossRef]
- Madenjian, C.P.; O’Connor, D.V.; Nortrup, D.A. A new approach toward evaluation of fish bioenergetics models. Can. J. Fish. Aquat. Sci. 2000, 57, 1025–1032. [Google Scholar] [CrossRef]
- Madenjian, C.P.; Ebener, M.P.; Sepúlvede, M.S. PCB concentrations of lake whitefish (Coregonus clupeaformis) vary by sex. J. Great Lakes Res. 2015, 41, 1185–1190. [Google Scholar] [CrossRef]
- Madenjian, C.P.; Keir, M.J.; Whittle, D.M. Sexual difference in mercury concentrations of lake trout (Salvelinus namaycush) from Lake Ontario. Chemosphere 2011, 83, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Madenjian, C.P.; Blanchfield, P.J.; Hrenchuk, L.E.; Van Walleghem, J.L.A. Mercury elimination rates for adult northern pike Esox lucius: Evidence for a sex effect. Bull. Environ. Contam. Toxicol. 2014, 93, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Gerstenberger, S.L.; Dellinger, J.A. PCBs, mercury, and organochlorine concentrations in lake trout, walleye, and whitefish from selected tribal fisheries in the upper Great Lakes region. Environ. Toxicol. 2002, 17, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Gewurtz, S.B.; Bhavsar, S.P.; Fletcher, R. Influence of fish size and sex on mercury/PCB concentration: Importance for fish consumption advisories. Environ. Int. 2011, 37, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Madenjian, C.P. Sex effect on polychlorinated biphenyl concentrations in fish: A synthesis. Fish Fish. 2011, 12, 451–460. [Google Scholar] [CrossRef]
- Raymond, B.; Rossmann, R. Total and methyl mercury accumulation in 1994–1995 Lake Michigan lake trout and forage fish. J. Great Lakes Res. 2009, 35, 438–446. [Google Scholar]
- Weis, P.; Ashley, J.T.F. Contaminants in fish of the Hackensack Meadowlands, New Jersey: Size, sex, and seasonal relationships as related to health risks. Arch. Environ. Contam. Toxicol. 2007, 52, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Ebener, M.P.; Brenden, T.O.; Wright, G.M.; Jones, M.L.; Faisal, M. Spatial and temporal distributions of lake whitefish spawning stocks in Northern lakes Michigan and Huron, 2003–2008. J. Great Lakes Res. 2010, 36 (Suppl. 1), 38–51. [Google Scholar] [CrossRef]
- Christensen, J.M. Burning of otoliths, a technique for age determination of soles and other fish. J. Cons. Perm. Int. Exp. Mer 1964, 29, 73–81. [Google Scholar] [CrossRef]
- USEPA (United States Environmental Protection Agency). Method 7473: Mercury in Solids and Solutions by Thermal Decomposition, Amalgamation, and Atomic Absorption; Office of Water: Washington, DC, USA, 2007.
- USEPA (United States Environmental Protection Agency). Guidelines Establishing Test Procedures for the Analysis of Pollutants; Office of Water: Washington, DC, USA, 1990.
- USEPA (United States Environmental Protection Agency). Method 1630: Methyl Mercury in Water by Distillation, Aqueous Ethylation, Purge and Trap, and Cold Vapor Atomic Fluorescence Spectrometry; Office of Water: Washington, DC, USA, 1998.
- Niimi, A.J. Biological and toxicological effects of environmental contaminants in fish and their eggs. Can. J. Fish. Aquat. Sci. 1983, 40, 306–312. [Google Scholar] [CrossRef]
- Claramunt, R.M.; Michigan Department of Natural Resources, Charlevoix, MI, USA. Personal communication, 2015.
- Rudstam, L.G.; Binkowski, F.P.; Miller, M.A. A bioenergetics model for analysis of food consumption patterns of bloater in Lake Michigan. Trans. Am. Fish. Soc. 1994, 123, 344–357. [Google Scholar] [CrossRef]
- Madenjian, C.P.; Pothoven, S.A.; Kao, Y.-C. Reevaluation of lake trout and lake whitefish bioenergetics models. J. Great Lakes Res. 2013, 39, 358–364. [Google Scholar] [CrossRef]
- Madenjian, C.P.; O’Connor, D.V.; Pothoven, S.A.; Schneeberger, P.J.; Rediske, R.R.; O’Keefe, J.P.; Bergstedt, R.A.; Argyle, R.L.; Brandt, S.B. Evaluation of a lake whitefish bioenergetics model. Trans. Am. Fish. Soc. 2006, 135, 61–75. [Google Scholar] [CrossRef]
- Mohr, L.C.; Ebener, M.P. The coregonine community. In The State of Lake Huron in 1999; Ebener, M.P., Ed.; Great Lakes Fishery Commission Special Publication: Ann Arbor, MI, USA, 2005; pp. 69–76. [Google Scholar]
- Hirayama, K.; Yasutake, A. Sex and age differences in mercury distribution and excretion in methylmercury-administered mice. J. Toxicol. Environ. Health 1986, 18, 49–60. [Google Scholar] [CrossRef] [PubMed]
- National Academy of Sciences. Toxicological Effects of Methylmercury; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- Yasutake, A.; Hirayama, K.; Inoue, M. Mechanism of urinary excretion of methylmercury in mice. Arch. Toxicol. 1989, 63, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Naganuma, A.; Kobayashi, K.; Imura, N. An explanation for strain and sex differences in renal uptake of methylmercury in mice. Toxicology 1991, 69, 317–329. [Google Scholar] [CrossRef]
- Tanaka, T.; Naganuma, A.; Miura, N.; Imura, N. Role of testosterone in γ-glutamyltranspeptidase-dependent renal methymercury uptake in mice. Toxicol. Appl. Pharmacol. 1992, 112, 58–63. [Google Scholar] [CrossRef]
- Bryan, M.B.; Scott, A.P.; Li, W. Sex steroids and their receptors in lampreys. Steroids 2008, 73, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Madenjian, C.P.; Johnson, N.S.; Siefkes, M.J.; Dettmers, J.M.; Blum, J.D.; Johnson, M.W. Mercury accumulation in sea lamprey (Petromyzon marinus) from Lake Huron. Sci. Total Environ. 2014, 470–471, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Madenjian, C.P.; O’Connor, D.V. Trophic transfer efficiency of mercury to lake whitefish Coregonus clupeaformis from its prey. Bull. Environ. Contam. Toxicol. 2008, 81, 566–570. [Google Scholar] [CrossRef] [PubMed]
- Madenjian, C.P.; David, S.R.; Krabbenhoft, D.P. Trophic transfer efficiency of methylmercury and inorganic mercury to lake trout Salvelinus namaycush from its prey. Arch. Environ. Contam. Toxicol. 2012, 63, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Madenjian, C.P.; Francis, J.T.; Braunscheidel, J.J.; Bohr, J.R.; Geiger, M.J.; Knottnerus, G.M. Mercury concentrations of bluegill (Lepomis macrochirus) vary by sex. Environments 2015, 2, 546–564. [Google Scholar] [CrossRef]
- Monteiro, L.R.; Isidro, E.J.; Lopes, H.D. Mercury content in relation to sex, size, age and growth in two scorpionfish (Helicolenus dactylopterus and Pontinus kuhlii) from Azorean waters. Water Air Soil Pollut. 1991, 56, 359–367. [Google Scholar] [CrossRef]
- Stafford, C.P.; Hansen, B.; Stanford, J.A. Mercury in fishes and their diet items from Flathead Lake, Montana. Trans. Am. Fish. Soc. 2004, 133, 349–357. [Google Scholar] [CrossRef]
- OMOE (Ontario Ministry of the Environment). Guide to Eating Ontario Sport Fish 2013–2014; Queen’s Printer for Ontario: Toronto, ON, Canada, 2013. [Google Scholar]
- Becker, D.S.; Bigham, G.N. Distribution of mercury in the aquatic food web of Onondaga Lake, New York. Water Air Soil Pollut. 1995, 80, 563–571. [Google Scholar] [CrossRef]
- USFDA (United States Food and Drug Administration). Health and Human Services; Public Health Service: Rockville, MD, USA, 2015.
- Amrhein, J.F.; Stow, C.A.; Wible, C. Whole-fish versus filet polychlorinated-biphenyl concentrations: An analysis using classification and regression tree models. Environ. Toxicol. Chem. 1999, 18, 1817–1823. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Attribute | Females (n = 26) | Males (n = 34) |
|---|---|---|
| Total length (mm) | 515 (5) {468–548} | 501 (4) {451–549} |
| Weight (g) | 1192 (40) {825–1670} | 1012 (27) {740–1360} |
| Age (years) | 10.4 (0.4) {7–17} | 10.9 (0.4) {7–17} |
| Mean asymptotic length (mm) | 556 | 542 |
| Brody growth coefficient (year−1) | 0.165 | 0.138 |
| Characteristic | Mean |
|---|---|
| Hg concentration in somatic tissue (ng/g) | 68.3 (6.5) {44.9–84.7} |
| Hg concentration in ovaries (ng/g) | 4.9 (0.6) {3.3–6.9} |
| Expected percent change in whole-fish Hg concentration | |
| immediately after spawning due to release of eggs (%) | +17.9 (2.8) {+11.2–+25.4} |
| GSI (%) | 16.1 (2.0) {11.1–21.3} |
| Age (years) | Females | Males | ||||
|---|---|---|---|---|---|---|
| Weight at Age (g) | Cumulative Consumption (g) | Cumulative GGE | Weight at Age (g) | Cumulative Consumption (g) | Cumulative GGE | |
| 0 | 30 | 240 | 0.125 | 30 | 240 | 0.125 |
| 1 | 168 | 900 | 0.187 | 149 | 825 | 0.181 |
| 2 | 305 | 2188 | 0.139 | 268 | 1944 | 0.138 |
| 3 | 443 | 4124 | 0.107 | 387 | 3610 | 0.107 |
| 4 | 580 | 6733 | 0.086 | 505 | 5842 | 0.086 |
| 5 | 718 | 12,277 | 0.058 | 624 | 10,563 | 0.059 |
| 6 | 855 | 19,023 | 0.045 | 743 | 16,311 | 0.046 |
| 7 | 993 | 26,770 | 0.037 | 862 | 23,080 | 0.037 |
| 8 | 1060 | 34,979 | 0.030 | 908 | 30,092 | 0.030 |
| 9 | 1119 | 43,827 | 0.026 | 948 | 37,422 | 0.025 |
| 10 | 1171 | 48,963 | 0.024 | 985 | 41,649 | 0.024 |
| 11 | 1217 | 54,311 | 0.022 | 1018 | 46,018 | 0.022 |
| 12 | 1256 | 59,841 | 0.021 | 1046 | 50,503 | 0.021 |
| 13 | 1290 | 65,531 | 0.020 | 1072 | 55,105 | 0.019 |
| 14 | 1320 | 71,366 | 0.018 | 1095 | 59,807 | 0.018 |
| 15 | 1345 | 77,323 | 0.017 | 1114 | 64,594 | 0.017 |
| 16 | 1366 | 83,381 | 0.016 | 1132 | 69,464 | 0.016 |
| 17 | 1385 | 89,533 | 0.015 | 1147 | 74,406 | 0.015 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madenjian, C.P.; Ebener, M.P.; Krabbenhoft, D.P. Mercury Accumulation, and the Mercury-PCB-Sex Interaction, in Lake Whitefish (Coregonus clupeaformis). Environments 2016, 3, 7. https://0-doi-org.brum.beds.ac.uk/10.3390/environments3010007
Madenjian CP, Ebener MP, Krabbenhoft DP. Mercury Accumulation, and the Mercury-PCB-Sex Interaction, in Lake Whitefish (Coregonus clupeaformis). Environments. 2016; 3(1):7. https://0-doi-org.brum.beds.ac.uk/10.3390/environments3010007
Chicago/Turabian StyleMadenjian, Charles P., Mark P. Ebener, and David P. Krabbenhoft. 2016. "Mercury Accumulation, and the Mercury-PCB-Sex Interaction, in Lake Whitefish (Coregonus clupeaformis)" Environments 3, no. 1: 7. https://0-doi-org.brum.beds.ac.uk/10.3390/environments3010007