Combined Effects of Trace Metals and Light on Photosynthetic Microorganisms in Aquatic Environment


Environmental Biogeochemistry and Ecotoxicology, Department F.-A. Forel for Environmental and Aquatic Sciences, School of Earth and Environmental Sciences, Faculty of Sciences, University of Geneva, Uni Carl Vogt, Bvd Carl-Vogt 66, CH-1211 Geneva 4, Switzerland
*
Author to whom correspondence should be addressed.
Environments 2018, 5(7), 81; https://0-doi-org.brum.beds.ac.uk/10.3390/environments5070081
Submission received: 14 June 2018
/
Revised: 6 July 2018
/
Accepted: 8 July 2018
/
Published: 12 July 2018
(This article belongs to the Special Issue Environmental Toxicology of Trace Metals)
Abstract
:In the present review, we critically examine the state-of-the-art of the research on combined effects of trace metals and light on photosynthetic microorganisms in aquatic environment. Light of different intensity and spectral composition affects the interactions between trace metals and photosynthetic microorganisms directly, by affecting vital cellular functions and metal toxicokinetics and toxicodynamics, and indirectly, by changing ambient medium characteristics. Light radiation and in particular, the ultraviolet radiation component (UVR) alters the structure and reactivity of dissolved organic matter in natural water, which in most of the cases decreases its metal binding capacity and enhances metal bioavailability. The increase of cellular metal concentrations is generally associated with increasing light intensity, however further studies are necessary to better understand the underlying mechanisms. Studies on the combined exposures of photosynthetic microorganisms to metals and UVR reveal antagonistic, additive or synergistic interactions depending on light intensity, spectral composition or light pre-exposure history. Among the light spectrum components, most of the research was performed with UVR, while the knowledge on the role of high-intensity visible light and environmentally relevant solar light radiation is still limited. The extent of combined effects also depends on the exposure sequence and duration, as well as the species-specific sensitivity of the tested microorganisms and the activation of stress defense responses.
1. Introduction
Studying the combined action of multiple stressors of different nature is currently one of the hot topics in ecotoxicology [1]. Hence, the role of environmental stressors on micropollutant effects, including trace metals, received considerable attention in the past 10 years. Various studies exploring the combined action of environmental factors (temperature, nutrients,) and micropollutants on model organisms [2,3,4,5] highlighted different type of stressor interactions. Combined action might occur when organisms are challenged to face simultaneous exposure to multiple stressors (Figure 1). In such case the induced cumulative stress response might correspond to the sum of the observed effects when the microorganism is exposed to each single stressor (Figure 1A). However, very often, the environmental stressor and the trace metal interact [2,3,6]. In such a case, the resulting effects might be lower (antagonism) or higher (synergism) than the sum of the effects induced by the individual stressors. Such synergistic or antagonistic interactions among stressors are generally associated with common modes of action of the two micropollutants. Hence, examining the underlining mechanisms of two (or more) stressor interactions is essential for the understanding of their combined effects [7]. Furthermore, in complex environmental settings, organisms often cope with one stressor when exposure to a second stressor occurs. Three possible scenarios should be further consider [4] (Figure 1B): (a) the microorganisms respond to the prolonged exposure to the stressor with acclimation, however the acclimation does not interfere with the sensitivity to the micropollutant; (b) acclimated microorganisms present a decreased sensitivity to the micropollutant, in this case co-tolerance between the environmental stressor and the micropollutant is observed; (c) the environmental factors cause a stress to the microorganism that is already at its tolerance limits, the addition of the micropollutant can only increase the experienced stress. The occurrence of one of these scenarios depends on the ability of the organism to acclimate to the stressor. Indeed, any increase or decrease in sensitivity is also associated with the modes of action of the combined stressors and the type of activated stress responses [5].
Nevertheless, the combined effects in the natural environment are difficult to quantify due to the complexity and non-linear relationships between the observed effects and the combination of environmental stressor(s) and micropollutant diversity [6]. Indeed, the combined action of two stressors might have unpredictable indirect effects through biota species interactions such as competition, presence of predators or pathogens [8]. The sensitivity to stressors could differ considerably among species and vary in time and in intensity [5]. Exposure sequence and duration could be also important in the sense that biological receptors might respond to the stressor activating cellular responses that might alter micropollutant toxicokinetics and toxicodynamics and thus result in increased or reduced sensitivity [4]. Moreover the boundary between “environmental factor” and “environmental stressor” is hardly defined since is related to the debated definition of stress. In present review we adopt the stress concept that “stress occur when an organism is forced outside its normal operating range” [9]. Among the various environmental stressors, the present review focusses on light irradiation and trace metals, which affect photosynthetic microorganisms in the aquatic environment (Figure 2). Photosynthetic (micro)organisms play a pivotal role in the element biogeochemical cycles, energy and matter fluxes [10].
Trace metals are ubiquitous and conservative micropollutants [11]. Following the interactions with different constituents in the aquatic environment, the trace metals are present under different chemical forms [12], which determine to a large extent their fate, bioavailability and toxicity [13,14,15].
Various environmental factors, including light, might influence metal speciation and bioavailability [2,4], as well as the microorganism “state” and consequently affect the metal uptake and the associated cellular effects [4]. Light radiation is one of the most variable environmental parameters. The light intensity and spectral composition change on a daily scale and depend on cloud coverage, air vapor or aerosol composition. Both photosynthetically active radiation (PAR) and ultraviolet radiation (UVR) have an essential role in the aquatic environment but might become stressors for photosynthetic microorganisms [19,20].
The objective of this review paper is to provide an overview of state-of-the-art of the knowledge and discuss the existing gaps concerning the combined interaction of trace metals and light with photosynthetic microorganisms in the aquatic environment. Given the rather large subject to be covered, we focus specifically on the light as an environmental stressor; and its role in (i) trace metal speciation and bioavailability; (ii) change of the oxidative activity in the aquatic systems; (iii) possible modifications of trace metal toxicokinetics and (iv) alteration of the vital cellular functions of photosynthetic microorganisms and toxicodynamics. The role of light pre-exposure history and acclimation will be also discussed.
2. Light Radiation from Environmental Factor to Stressor
Natural solar radiation is operationally divided in visible light and ultraviolet radiation, both playing crucial role in the aquatic environment. Solar light is essential to photosynthetic organisms converting light into chemical energy. However, the excess of absorbed light energy, which is neither used for photosynthetic processes nor dissipated by photoprotective mechanisms, could induce a stress to the cells. The visible light-induced stress directly correlates with the photosynthetic capacity of the cell, which in turn varies considerably among phytoplankton species and can be also affected by the environmental stressors, such as high salinity or nutrient deprivation [20]. The UVR-induced stress is associated with its high energy, which might transfer to biomolecules and dissipate via various photochemical reactions. UVR effects depend on its intensity and spectral composition with the shorter and energy-rich wavelength having stronger potential to degrade various types of biological molecules. Indeed, UVR was shown to causes DNA/RNA lesion and protein degradation [21,22,23]. In photosynthetic organisms, UVB could inactivate photosynthesis causing damages to photosystem I (PSI) and photosystem II (PSII), to oxidize and thereby bleach the photosynthetic pigments [23], to generate excessive levels of reactive oxygen species (ROS) [21] and to induce oxidative stress [24]. Photosynthetic microorganisms have developed several protective strategies against UVR, encompassing DNA repair and photoreactivation, increase in the antioxidant response activity, species specific UVR avoidance via EPS production and mat formation in some cyanobacteria or production of screening molecules [21,22,25,26,27]. Evidences exist that the increased solar radiation, in particular UVR-component of the spectrum, has an important deleterious influence not only on the human health [28], but also to the environment and affects different element biogeochemical cycles [29,30]. It is anticipated that future changes in the surface radiation and interactions with climate variability [30] will influence the aquatic ecosystems, thus increasing the uncertainty in predicting the environmental risks associated with chemical pollution.
3. Light as a Modifier of the Ambient Medium Characteristics
The interplay between light and trace metals can modify the chemical speciation (and thus the bioavailability) in the ambient medium by (i) altering the amount, the structure and reactivity of DOM [30,31,32] and (ii) enhancing the oxidative activity in the aquatic systems [29,33].
3.1. Photo-Transformations of Dissolved Organic Matter
Among the various components in the aquatic systems, the dissolved organic matter (DOM) is central in protecting the aquatic organisms from different kinds of stress. DOM is known to protect photosynthetic organisms by (i) binding different trace metals and thus decreasing the proportion of the free metal ions and their bioavailability and (ii) absorbing the large part of the solar radiation (in particular more detrimental UVR component), thus playing role of sunscreen (Figure 2). Therefore, the variation of the environmental factors that could affect DOM concentration, structure or reactivity will influence its protective capacity to photosynthetic organisms. Sunlight, and in particular UV-B component of increased intensity), could alter both DOM quality and quantity (as reviewed in Ref. [29,30]) via different transformation processes including mineralization, photo-degradation [34,35,36,37], loss of suspended particulate organic carbon and production of the dissolved organic carbon (e.g., by photo-dissolution) [38,39,40]. Photo-alteration of DOM structure was studied by using a combination of large array of the analytical tools, probing different characteristics of such complex and “ill-defined” compounds: UV-VIS spectrophotometry and excitation-emission matrixes, infrared spectroscopy, flow field-flow fractionation multi-detection and size exclusion chromatography, liquid chromatography coupled to UV and organic carbon detection [30,41,42]. For example, UVR altered DOM fluorescence properties and decreased the aromaticity and its molecular masses, and induced a formation of low-molecular-mass products (Figure 3). A decrease in the aromaticity and UV absorbing properties together with no changes in the DOC concentrations was hypothesized to correspond to conversion of the aromatics moieties into low-molecular-mass-organic compounds. The extent of photo-transformations depended thus on the type and origin of the DOM. For example, irradiation of the allochthonous DOM [43] produced low molecular mass biologically labile products, which was not the case for the algal derived DOM [44]. In contrast, the photo-bleaching appears independent on the sources, both allochthonous [36] and autochthonous [37] were bleached by 20–50% per unit of residual dissolved organic carbon. Similarly, photo-degradation on phytoplankton-derived DOM exposed to sunlight resulted in an overall decrease in DOM aromaticity. Exposure to UV radiation for six days induced a photo-mineralization of DOM and attached organic matter (AOM) to algal cells isolated form eutrophic lake Taihu, affecting tyrosine-like, humic-like and tryptophan-like substances, but the photo-degradation of AOM was lower [45]. Four days exposure to sun of the allochthonous DOM in sub-alpine Lake Tiancai, impacted by climate change resulted in a decrease of molecular mass, but an increase in the aromaticity of DOM, as well as photo-mineralization to CO2 of 32% of DOM [46].
Moreover, the observed photo-alteration of model DOM, such as Suwannee River fulvic acid (SRFA) and Suwannee River humic acid (SRHA), or Elliot humic acid (EHA) appeared at low radiation intensity and was not directly proportional to the received irradiance doses [41,42]. Photochemical behavior of total dissolved organic matter (<0.22 μm), high-molecular-weight (HMW, 1 kDa–0.22 μm) DOM and low-molecular-weight (LMW, <1 kDa) isolated form Yangtze River, its coastal area, and the Pacific Ocean was compared. Humic-like and tryptophan-like components in the riverine, coastal, and oceanic surface waters were photodegraded, but the degradation degree depended on both the DOM source and molecular weight [47]. Fifty eight-hour exposure to sunlight resulted in a full degradation of the humic-like component and induced a transformation of the EPS into transparent exopolymer particles (TEP) and unstable flocs of different size of microbial mat samples Bosten Lake [48]. The flock formation was shown to depend on the irradiation light spectral composition, with EPS isolated form cyanobacterium Chroococcus minutus forming unstable flocs under UVR, but stable flocs under solar light [49].
3.2. Consequences for Trace Metal Speciation
The photo-induced transformations of DOM could results in: (i) a decrease of DOM concentrations or alteration of its structure and thus increase in the free metal ion concentration and bioavailability [34,50]; (ii) no change in the free metal ion concentrations, because of the production of low molecular weight dissolved organic acids able to scavenge dissolved metal [51]; (iii) an increase of the metal complexation because of a net production of functional groups on humic-like fractions that outcompete for the complexation by low molecular weight dissolved organic acids [34].
Most of the available literature revealed a decrease of metal complexation by DOM and consequently an increase of the free metal ion concentrations as results of DOM photo-transformation [34,52,53,54,55,56]. Selected examples include, an increased free copper-ion concentrations, [Cu2+] in UV-B irradiated organic rich estuarine waters [52]; riverine HS [34], alpine lake water samples [53], organic-rich estuarine waters [52]. Similarly, an enhancement of free lead ion concentrations, [Pb2+] in the estuarine samples after UVR [56] and UVR irradiated DOM of different origin such as waste water treatment plant effluents ([41], Figure 4). However, no clear relationship between the radiation dose and the amount of the released [Pb2+] was found for SRFA, SRHA [41] and EHA [42], suggesting that low-molecular-mass products also exhibit lead binding properties. Few examples illustrating the increase of the metal complexation by the photo-altered DOM can be also found. Cu complexation by wetland HS was increased upon photo-transformation [34] and Pb binding by sun-altered OM from WWTP Hinwill (Figure 4). An increase of [Cd2+] via photolytic release of bound cadmium from Cd-contaminated fulvic acid was found, whereas no changes were observed in Cd-amended natural lake water HS [55]. The effects of the light on the metal speciation were DOM-nature dependent. For example, the concentration of the free lead ions increased in presence of sun-altered DOM following the order EHA > Negro River DOM > SRHA > SRFA > AHA (Figure 4). No effect was observed for organic matter isolates from waste water treatment plants (WWTP) in Duebendorf (Db) and a significant decline was found for [Pb2+] in the presence of sun-altered OM from WWTP Hinwill (Hw).
Very little information exists on the kinetics of the metal release from the photo-transformed DOM or on the size speciation currently. The progress in the development of different speciation techniques capable of providing specific metal species [59,60] gave further opportunities to explore the dynamic aspects of the metal speciation [61]. However, the capabilities of these techniques are rarely used to follow the effects of the light and photo-transformed DOM on the metal speciation. One of the few examples revealed diurnal changes in Cu, Cd and Pb speciation in the contaminated Riou Mort river (France), which coincided with changes in water quality parameters (pH, O2, temperature, DOM) associated with the photosynthetic activity of benthic phytoplankton and their EPS and exudates production [11,62].
3.3. Consequences for Trace Metal Bioavailability
Even more scarce and inconclusive is the literature about the metal bioavailability in the presence of photo-altered DOM. Cu, Zn, Co and Pb, but not Ni and Cd toxicity to the freshwater green alga Pseudokirchneriella subcapitata increased up to 78% after 10 days of UVR of high- and low-organic carbon water samples [63]. Pb, Cu, Ni, and Cd concentrations required to reduce algal growth by 50% was significantly decreased with both solar and UVR doses, however the changes in toxicity with UVR-dose were inconsistent among the metals tested [64]. Cu toxicity to larva Pimephales promelas increased in the presence of photo-oxidated DOM, following model predictions based on [Cu2+] [65]. Photolysis of DOM was associated with an increase in free copper ions [42,51] and to a subsequent increase in bioavailability and toxicity to eukaryotic algae [42,63].
3.4. Light-Induced Enhancement of the Oxidative Activity in Ambient Medium
Solar radiation is known to enhance the oxidative reactivity of the aquatic systems [29] and thus to promote the oxidative damage in the biota. Indeed the UV-induced photoreactions of inorganic nitrogen, DOM, and metals results in a production of the ROS [30,66] such as superoxide anions, hydrogen peroxide, singlet oxygen and hydroxyl radicals. Although it is well documented that the ROS produced inside the cell can adversely affect the vital cellular function, very little is known about the effect of externally generated ROS (i.e., species produced in the water surrounding the cell) on the aquatic organisms [66] and plants [67]. In is generally accepted that the impact of externally generated ROS on biota is inversely related to their properties and reactivity [66]: highly reactive and short-lived radicals, such as •OH radicals, are expected to have minimal impact. Enhancement of the oxidative activity in chemical medium may also affect microorganisms through the production of toxic trace metal species. For example, the photolysis of organic Cu-complexes and interactions with superoxide ion radical O2•− may be anticipated to increase the Cu bioavailability and hence Cu toxicity to phytoplankton [66]. The reduction of the Cu(II) to Cu(I) in oxic aquatic systems occurred to a large extend via UV-induced chemical reactions of Cu(II) complexed to strong ligands [51,68,69]. The photochemical properties of the organic exudate secreted by a toxic strain of cyanobacterium Microcystis aeruginosa were studied by measuring ROS generation and redox transformations of Fe in the presence of the organic exudate under acidic (pH 4) and alkaline (pH 8) conditions. Simulated sunlight-irradiation of organic exudate released by M. aeruginosa generates nanomolar concentrations of superoxide and hydrogen peroxide. The photo-generated superoxide was shown to plays an important role in Fe(III) reduction under alkaline conditions, leading to an increase of Fe availability in natural waters in the presence of these algal exudates [70]. This interaction with transition metals is likely to be one of the main processes through which photo-chemically produced or other charged externally generated ROS, can have an adverse effect on aquatic biota; but further studies are needed to ascertain the ecological impact of these types of reactions in natural waters. Indeed, high ROS levels were found during photo-oxidation of DOC for Amazon River, no oxidative stress in fish Hemigrammus levis was detected [71].
4. Light as a Modifier of the Trace Metal Effect to Photosynthetic Microorganisms
4.1. Toxicokinetics Aspects
Despite that trace metal toxicokinetics was extensively studied in photosynthetic microorganisms [16], very little is known about light-dependent changes in metal uptake and intracellular compartmentalization. Cd and Zn uptake rates in M. aeruginosa significantly increased with augmentation of visible light from 18 to 55 µmol photons m−2 s−1 [72]. Intracellular Zn distribution stayed unchanged during light irradiation, while enhanced Cd accumulation in granules were reported under higher light conditions (55 µmol photons m−2 s−1) [72]. These results shed some light on the role of visible light on metal toxicokinetics, however although environmentally relevant these low-light irradiation conditions do not represent a stress for photosynthetic organisms. Zn uptake was also determined in three cyanobacterial species acclimated to low (50 µmol photons m−2 s−1) and high-visible light (300 µmol photons m−2 s−1) [73,74]. For the two strains of M. aeruginosa a significant increase in the intracellular Zn was observed under high-light irradiation, whereas no significant differences were found in Synechocystis sp. suggesting that the influence of light on metal uptake is species specific [73]. A positive correlation between light radiation intensity and metal intracellular content was found for Cu in the green alga C. reinhardtii. In this study intracellular copper content was higher in cells exposed to solar simulated light as compared with cells incubated under low artificial light [75]. Significant increase of intracellular Cu was also found in the cyanobacterium Anabaena doliolum after short-term exposure to UVB radiation. However, under such conditions, it is possible that the intracellular increase induced by UVR was not associated with metal cellular uptake alteration but to membrane damages [76].
An increase in the intracellular metal quota is the prerequisite for toxicity to occur, however, only very few studies investigated intracellular metal content in parallel to cellular effects. A direct correlation was observed between light-associated variations of intracellular Cu, Cd and Zn concentration and growth, photosynthetic activity and oxidative stress [72,75,76]. Even less is known about sub-cellular metal distribution and other detoxification strategies, therefore further studies are highly sought to better elucidate the possible role of light irradiation on trace metal toxicokinetics. Overall, the scarce information available on trace metal toxicokintetics under different light conditions is obtained in studies with three “classical” trace metals and very few model cyanobacteria and green algae, thus further studies are necessary to see to what extent the current findings can extend to other trace metals and microorganisms.
4.2. Mode-of-Action and Multistressor Interactions
Trace metal toxicity is generally associated with three main modes-of-action: (i) blocking of the functional groups of essential biomolecules; (ii) displacement of essential metal ions in biomolecules; (iii) modification of the biomolecule conformations [77]. The resulting effects depend on the affected cellular compartment and on the biological target. Among the multiple effects of metals on photosynthetic microorganisms, those on the photosynthetic machinery are of particular relevance for understanding the interplay with light. Toxic and excessive concentrations of essential metals were reported to bind and destabilize different proteins of both photosystem II (PS II) and I (PS I) [78] and to inhibit chlorophyll biosynthesis [79]. As a result, a reduction of photosynthetic yield [80,81,82] and chlorophyll bleaching [83,84,85] are commonly observed endpoint in metal toxicity tests with algae. Another major mode-of-action of metals in phytoplankton is the excessive cellular ROS generation. ROS are normal by-products of the oxidative metabolism and are important signal molecules with a central role in the activation of cellular stress responses [19]. In photosynthetic microorganisms ROS are continuously produced via oxidative phosphorylation and at the level of photosystem I and II (PS I and PS II) [86]. When cells are exposed to metals the amount of cellular ROS might exceeds their antioxidant capacity, oxidative stress occurs [87]. Oxidative stress can act at the level of different physiological processes and target different kind of cellular components. It might cause lipid peroxidation and membrane alteration, protein degradation and enzyme inactivation, DNA and RNA damages [88]. Given the potential of metals to induce oxidative stress in phytoplankton, increase in ROS and activation of the antioxidant response are common endpoints reported in algal toxicity tests with metals [89,90,91,92].
Light exhibits overlapping mode-of-action with those of trace metals. When light irradiation exceeds the photosynthetic capacity of the microorganism significant levels of ROS are generated and damages at the photosynthetic apparatus observed [19]. Singlet oxygen (1O2) formation occur mainly in PS II, while PS I contribute to the formation of superoxide anions (O2•−) by univalent reduction of O2. In fact, when light intensity is high, oxygen can be used as electron acceptor on the donor side of PS I (Mahler reaction) [93]. Phytoplankton cope with excess of light and associated ROS generation in different ways. They can use avoidance strategies, (e.g., phototaxis in C. reinhardtii) [94], regulate the light harvesting domain, increase the amount of energy dissipated as heat via non-photochemical quenching (NPQ) and/or protect the cell from oxidative stress with antioxidant molecules and enzymes [19,93]. Oxidative stress and pigment bleaching are the most relevant effects observed in phytoplankton cells exposed to excess of visible light [95] and to high UVB doses [24]. Given similar mode-of-action in photosynthetic microorganisms, exposure to metals can enhance the ROS formation due to the alteration of the photosynthetic electron chain [87].
Considering the overlapping mode-of-action and thus the possible interaction, it can be expected that combined exposure to the light and trace metals might results either in synergistic effects (enhanced sensitivity) or in antagonistic ones (reduced sensitivity) (See Section 4.3). There is thus a high uncertainty in the prediction of the combined action of these stressors. What is more, some metals, such as Cu and Fe, are involved in Haber Weiss cycle and Fenton reactions that lead to the production of highly reactive hydroxyl radical (●OH) from the less reactive superoxide radical (O2●−) and hydrogen peroxide (H2O2) respectively [87], which makes even more complex the understanding of their interactions with light. On the other hand cellular stress response pathways activated to face excess of visible light and UVB might be effective to cope with excess in intracellular metal quota, thus reducing toxic effects induced by metal. A deeper mechanistic understanding of the cellular events involved in trace metal-light interactions is required. Gaining such knowledge could contribute to a possible extension of the adverse outcome pathways (AOP) concept including the changes in environmental factors [96], which is currently underdeveloped.
4.3. Toxicodynamics Aspects
A significant number of studies explored the combined effects of metals and light on different eukaryotic algae and cyanobacteria [7,72,73,74,75,76,97,98,99,100,101], as well as periphytic communities [102,103] (Table 1). The research focused on different metals including Zn [72,73,74,103], Cd [7,64,72,98,100,102], Hg [100], Cu [62,75,76,97], Ni [64], Pb [64] and on different light radiation components: visible [72,73,74,75,99,102] and UV radiation [7,75,76,97,98,100,101]. Most of the work explored these combined effects on model organisms, mainly cyanobacterial species [72,73,74,99], but the results are rather contrasting. Only one work focused on periphytic communities [103]. Most of the studies performed with isolated species revealed an enhancement of the metal effects on photosynthetic yield and growth inhibition under increased visible light radiation [72,73,74,99]. Similarly the intracellular ROS generation, lipid peroxidation and enzymatic antioxidant response increased upon exposure of cyanobacterium Nostoc muscorum to Hg with increase of the visible light radiation from 10 to 130 µmol photons m−2 s−1 [99].
Visible light-induced effects are receiving attention only recently in environmental toxicology while more consideration was given to UVR radiation, in particularly UVB component. The effects of UVB radiation on phytoplankton was thoroughly investigated and consequently combined action of UVB and other environmental stressors such as metals was also considered [20,104]. Most of the available works focused on the combined effects of UVR and Cd and revealed contrasting results [7,98,100]. The combination of UVB and Cd yielded synergistic effect on growth inhibition, photosynthetic yield and lipid peroxidation in cyanobacterium Plectonema boryanum [98], but induced antagonistic effect on the same endpoints in another cyanobacterium Anabaena sp. [100], demonstrating species-specificity of the responses. Cd effects to the green alga C. reinhardtii were investigated under a combination of Cd and UVA and UVB radiations [7], resulted in synergistic effects on several endpoints, notably photosynthetic yield and lipid peroxidation., while no significant difference on growth inhibition was found. The later research suggest that the type of interaction synergistic/additive or antagonistic can also differ according to measured biological effects. Short-term combined exposure to Cu and UVR induced a synergistic effects in growth, photosynthetic yield and lipid peroxidation of cyanobacterium A. doliolum, which were associated with an increase of the intracellular Cu content [76]. No significant changes in EC50 (growth inhibition) and decrease of photosynthetic yield were observed when the green alga Selenastrum capricornutum was exposed to Cu under light with different visible and UVR spectral composition and in presence of natural DOM [101], despite significant increase in Cu2+ ions were measured upon exposure of the natural water with high DOC content.
Most of the available studies addressed the effect of variable levels of visible light, or the effect of UV radiation (Table 1). However, in the natural environment, photosynthetic organisms are simultaneously exposed to visible and UVR light components, therefore the environmentally relevant knowledge about the interplay of solar light and trace metals is missing. The only study addressing the combined effects of Cu and simulated natural solar light on C. reinhardtii showed synergistic effects under solar simulated light with enhanced UVB radiation and antagonistic effects under solar simulated light with natural UVR intensity [75]. Overall, both increase and decrease of the trace metal adverse effects to photosynthetic microorganisms were reported for their combined exposure with light radiation (visible or UVR), depending on the microorganism and light intensity and spectral composition. However, most of the above-mentioned studies merely described the observed combined effects and the type of interaction, no information is available on the cellular mechanisms driving these interactions.
Another aspect that might contribute to the reported confounding trends is that light radiation sources and experimental set-ups differ considerably among the studies. The provided information about the exposure conditions is often incomplete, rendering very difficult inter-study comparability. Real light radiation intensity and spectrum are either not measured, or measured at the surface of exposed cultures, where it might differ considerably from those received by the cells within the culture. The UVR penetration in the exposed cell suspensions can also change among the treatments if cell densities differ significantly. To improve the comparability among experimental setting, UVA and UVB extinction coefficient for cyanobacterial and eukaryotic algal cultures were used to determine the average UVA and UVB intensities received by the microorganisms in the exposure vessel [105]. The implementation of the principles of sound ecotoxicology [106] will improve the consistency and reliability in the evaluation of the effects of multiple stressors to biota.
4.4. Role of the Light Pre-Exposure History and Acclimation
The available literature suggests a significant role of light pre-exposure history on metal toxicity. In a multiple stressor environment, acclimation to one stressor might increase (co-tolerance) or decrease the tolerance to other stressors [4]. Indeed, Zn effects on photosynthetic yield of periphytic communities increased with enhanced light radiation for low light-acclimated biofilms (25 µmol photons m−2 s−1). However, small increase in Zn toxicity was observed in periphyton acclimated to medium light (100 µmol photons m−2 s−1) and high light (500 µmol photons m−2 s−1) with respect to those acclimated to low light [103]. The concentration of Cd able to induce the half-maximal decrease (EC50) of photosynthetic yield resulted to be lower in periphytic communities grown under low UVR radiations as compared with those grown under high UVR radiation, indicating Cd co-tolerance of communities with higher tolerance to UVR. Community tolerance to the two stressors was associated with a shift in community composition (periphyton grown under high UVR were dominated by cyanobacteria) and a reduction of periphyton biomass, suggesting that the acclimation to UVR is associated with elevated metabolic costs [102]. For the cyanobacterium Synechocystis sp. acclimated to high visible light (300 µmol photons m−2 s−1) no increase in Zn toxicity was found suggesting that the combined effects of metals and visible light are species specific [73].
To shed light on the role of light acclimation in metal effects to phytoplankton, the green alga C. reinhardtii was exposed to Cu prior and after acclimation to high visible light (395 µmol phot m−2 s−1). The acclimation of C. reinhardtii to high light resulted in a significant decrease of the EC50s for growth inhibition, oxidative stress, membrane damages and chlorophyll fluorescence (Figure 5), indicating an increased sensitivity of C. reinhardtii to Cu. High intensity visible light per se represents a stressor for C. reinhardtii and the addition of Cu exacerbated the experienced stress e.g., by disturbing a cellular homeostasis that was already at its operating limit. The current results agree with what previously observed for the cyanobacterium M. aeruginosa [72,73,74], suggesting that similar interaction between metals and light irradiation occur in prokaryotic and eukaryotic phytoplankton. Interestingly different results were obtained when C. reinhardtii cells acclimated to low-intensity light (45 µmol phot m−2 s−1) were simultaneously exposed to a combination of high-intensity light (395 µmol phot m−2 s−1) and copper. Under these conditions, a significant increase of the EC50s for growth inhibition, oxidative stress, membrane damages and chlorophyll fluorescence was observed.
The common cellular responses activated to face oxidative stress induced by the exposure to high visible light and Cu, might be one of the reasons for the reduced sensitivity. Such results agree with the antagonistic effect observed for oxidative stress under combined exposure of C. reinhardtii to Cu and solar simulated light [73]. The antagonistic effect of these two stressors was associated with higher level of expression of genes encoding for antioxidant response enzymes. Overall, the extent of toxic action of trace metals and light radiation depended both on the sensitivity of the organism and on the activated stress responses. The study of multiple organisms, potentially with variable sensitivities to light and trace metals, is thus of primary importance in order to extrapolate the potential impact of these pollutants in natural environments where multiple interacting species are present.
5. Conclusions
Important advances were achieved with respect to the fundamental understanding of the role of the light on the trace metal speciation, bioavailabity and effects to photosynthetic microorganisms in aquatic environment. There is a consensus that light, of different intensity and spectral composition, could affect the directly or indirectly the interactions between trace metals and photosynthetic microorganism. Several interconnected mechanisms, relevant for surface waters, were identified: (i) alteration of the structure and reactivity of DOM, resulting in decrease of metal binding capacity and increase of metal bioavailability (and effects); (ii) effect on chemical speciation of the redox sensitive metals, such as Fe and Cu; (iii) enhancement of the oxidative activity in the aquatic systems; (iv) alteration of trace metal toxicokinetics, resulting generally in increase of the cellular metal with light intensity; (v) influence on the vital cellular functions of photosynthetic microorganisms and trace metal toxicodynamics. Light might either increase or decrease the trace metal effects depending on light intensity, spectral composition or light pre-exposure history. Among the light spectrum components, most of the studies focused on UVR, while the understanding of the role of the PAR and solar light in trace metal effects is overlooked. The results on the combined exposure of phytoplankton to metals and UVR revealed both positive or negative interactions between UVR and metal exposure, however available data comparability need to improve. The combined effects also depend on the exposure sequence and duration, as well as specific sensitivity of the tested species and the activation of stress responses.
Despite the increasing number of the studies dealing with combined effects of light and trace metals on photosynthetic microorganisms, numerous knowledge gaps sill persist. Future studies should focus on the improved understanding of the influence of light radiation on the chemodynamics of the processes underlying metal bioavailability in natural environment. The lack of knowledge on the dynamics of light induced trace metal transformations in the ambient medium and their interconnection with cellular concentrations and effects in our view is one of the important gap in the current knowledge. An improved knowledge of the combined action of light irradiation and metals at the molecular and cellular level, such as cellular stress responses and pathways activated to maintain cell homeostasis, would help better defining the mechanisms of interaction of these two stressors. What is more, considering the high biological diversity that characterizes photosynthetic microorganisms, a further research on species-specific light sensitivity and species-specific stress response is highly sought to evaluate the combined effects of trace metals and light in natural environments, where multiple species are present. Future studies might focus on those phytoplankton species that present different light response strategies, such as light stress avoidance or production of antioxidant and light screening pigments. Better understanding of the mechanisms driving trace metal effects in the complex environment settings will reduce the uncertainties associated with laboratory-to-field extrapolation of the toxicity data used in environmental risk assessment. Further experimentation under more environmentally relevant light irradiation conditions will enable relevant lab-to-field extrapolation of toxicity data. Examples include, light intensity and spectral composition comparable with the natural environment, sudden changes in light radiation, different pre-exposure history. In addition, in the natural environment, photosynthetic microorganisms might have to face stress pulses or chronic contamination. Pulsed and chronic exposure to metals and light irradiation might be very informative and could give an insight on possible phytoplankton stress responses and adaptation to the combination of the two stressors. What is more, in natural environments changes in light irradiation might indirectly affect photosynthetic organisms also by influencing other parameters such as water temperature, microcosm tests and field experimentation with planktonic or benthic community would be of key importance to gain fundamental knowledge on the role of light irradiation on the species–species interactions and consequences for trace metals effects in the complex environment settings.
Author Contributions
Both authors contributed to the paper conceptualization and review writing.
Funding
V.I.S. and G.C. gratefully acknowledge the financial support provided by Swiss National Science Foundation (SNSF) project 200021-134627, G.C. gratefully acknowledge the financial support provided by Swiss National Science Foundation project 164428_MHV fellowship.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Artigas, J.; Arts, G.; Babut, M.; Caracciolo, A.B.; Charles, S.; Chaumot, A.; Combourieu, B.; Dahllof, I.; Despreaux, D.; Ferrari, B.; et al. Towards a renewed research agenda in ecotoxicology. Environ. Pollut. 2012, 160, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Heugens, E.H.W.; Hendriks, A.J.; Dekker, T.; Straalen, N.M.V.; Admiraal, W. A review of the effects of multiple stressors on aquatic organisms and analysis of uncertainty factors for use in risk assessment. Crit. Rev. Toxicol. 2002, 31, 247–284. [Google Scholar] [CrossRef]
- Holmstrup, M.; Bindesbol, A.M.; Oostingh, G.J.; Duschl, A.; Scheil, V.; Kohler, H.R.; Loureiro, S.; Soares, A.M.; Ferreira, A.L.; Kienle, C.; et al. Interactions between effects of environmental chemicals and natural stressors: A review. Sci. Total Environ. 2010, 408, 3746–3762. [Google Scholar] [CrossRef] [PubMed]
- Fischer, B.B.; Pomati, F.; Eggen, R.I. The toxicity of chemical pollutants in dynamic natural systems: The challenge of integrating environmental factors and biological complexity. Sci. Total Environ. 2013, 449, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Segner, H.; Schmitt-Jansen, M.; Sabater, S. Assessing the impact of multiple stressors on aquatic biota: The receptor’s side matters. Environ. Sci. Technol. 2014, 48, 7690–7696. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.; Bednarska, A.J.; Kramarz, P.E.; Loureiro, S.; Scheil, V.; Kudlek, J.; Holmstrup, M. Interactions between toxic chemicals and natural environmental factors a meta-analysis and case studies. Sci. Total Environ. 2010, 408, 3763–3774. [Google Scholar] [CrossRef] [PubMed]
- Korkaric, M.; Behra, R.; Fischer, B.B.; Junghans, M.; Eggen, R.I. Multiple stressor effects in chlamydomonas reinhardtii toward understanding mechanisms of interaction between effects of ultraviolet radiation and chemical pollutants. Aquat. Toxicol. 2015, 162, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Clements, W.H.; Rohr, J.R. Community responses to contaminants: Using basic ecological principles to predict ecotoxicological effects. Environ. Toxicol. Chem. 2009, 28, 1789–1800. [Google Scholar] [CrossRef] [PubMed]
- Van Straalen, N.M. Ecotoxicology becomes stress ecology. Environ. Sci. Technol. 2003, 37, 324–330. [Google Scholar] [CrossRef]
- Lyons, T.W.; Reinhard, C.T.; Planavsky, N.J. The rise of oxygen in earth’s early ocean and atmosphere. Nature 2014, 506, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Tercier-Waeber, M.-L.; Stoll, S.; Slaveykova, V.I. Trace metal behavior in surface waters: Emphasis on dynamic speciation, sorption processes and bioavailability. Arch. Sci. 2012, 65, 1–24. [Google Scholar]
- Templeton, D.M.; Ariese, F.; Cornelis, R.; Danielsson, L.-G.; Muntau, H.; van Leeuwen, H.P.; Lobinski, R. IUPAC guidelines for terms related to speciation of trace elements. Pure Appl. Chem. 2000, 72, 1453–1470. [Google Scholar]
- Campbell, P.G.C. Interactions between trace metals and aquatic organisms: A critique of the free-ion activity model. Met. Speciat. Bioavailab. 1994, 2, 45–102. [Google Scholar]
- Wilkinson, K.J.; Buffle, J. Critical evaluation of physicochemical parameters and processes for modelling the biological uptake of trace metals in environmental (aquatic) systems. Physicochem. Kinet. Trans. Biointerfaces 2004, 9, 447–533. [Google Scholar]
- Buffle, J.; Wilkinson, K.J.; van Leeuwen, H.P. Chemodynamics and bioavailability in natural waters. Environ. Sci. Technol. 2009, 43, 7170–7174. [Google Scholar] [CrossRef] [PubMed]
- Worms, I.; Simon, D.F.; Hassler, C.S.; Wilkinson, K.J. Bioavailability of trace metals to aquatic microorganisms: Importance of chemical, biological and physical processes on biouptake. Biochimie 2006, 88, 1721–1731. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, M.; Le Faucheur, S.; Fortin, C.; Campbell, P.G. Cadmium detoxification strategies in two phytoplankton species: Metal binding by newly synthesized thiolated peptides and metal sequestration in granules. Aquat. Toxicol. 2009, 92, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Slaveykova, V.I.; Guignard, C.; Eybe, T.; Migeon, H.N.; Hoffmann, L. Dynamic nanosims ion imaging of unicellular freshwater algae exposed to copper. Anal. Bioanal. Chem. 2009, 393, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wakao, S.; Fischer, B.B.; Niyogi, K.K. Sensing and responding to excess light. Annu. Rev. Plant Biol. 2009, 60, 239–260. [Google Scholar] [CrossRef] [PubMed]
- Hader, D.P.; Williamson, C.E.; Wangberg, S.A.; Rautio, M.; Rose, K.C.; Gao, K.; Helbling, E.W.; Sinha, R.P.; Worrest, R. Effects of UV radiation on aquatic ecosystems and interactions with other environmental factors. Photochem. Photobiol. Sci. 2015, 14, 108–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.-Y.; Hader, D.-P. Reactive oxygen species and UV-B: Effect on cyanobacteria. Photochem. Photobiol. Sci. 2002, 1, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Häder, D.-P. UV-induced DNA damage and repair: A review. Photochem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Warwick, F.V.; Roy, S. Solar ultraviolet-b radiation and aquatic primary production: Damage, protection, and recovery. Environ. Rev. 1993, 1, 1–12. [Google Scholar]
- Cheloni, G.; Slaveykova, V. Photo-oxidative stress in green algae and cyanobacteria. React. Oxyg. Species 2018, 5, 126–133. [Google Scholar] [CrossRef]
- Singh, S.P.; Hader, D.P.; Sinha, R.P. Cyanobacteria and ultraviolet radiation (UVR) stress: Mitigation strategies. Ageing Res. Rev. 2010, 9, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Ambasht, N.K.; Sinha, J.P.; Klisch, M.; Häder, D.-P. UV-B-induced synthesis of mycosporine-like amino acids in three strains of nodularia (cyanobacteria). J. Photochem. Photobiol. B 2003, 71, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Laurion, I.; Lami, A.; Sommaruga, R. Distribution of mycosporine-like amino acids and photoprotective carotenoids among freshwater phytoplankton assemblages. Aquat. Microb. Ecol. 2002, 26, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Norval, M.; Cullen, A.P.; de Gruijl, F.R.; Longstreth, J.; Takizawa, Y.; Lucas, R.M.; Noonan, F.P.; van der Leun, J.C. The effects on human health from stratospheric ozone depletion and its interactions with limate change. Photochem. Photobiol. Sci. 2007, 6, 232–251. [Google Scholar] [CrossRef] [PubMed]
- Zepp, R.G.; Erickson, D.J., III; Paul, N.D.; Sulzberger, B. Interactive effects of solar uv and climate change on biogeochemical cycles. Photochem. Photobiol. Sci. 2007, 6, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Sulzberger, B.; Durisch-Kaiser, E. Chemical characterization of dissolved organic matter (DOM): A prerequisite for understanding UV-induced changes of DOM absorption properties and bioavailability. Aquat. Sci. 2009, 71, 104–126. [Google Scholar] [CrossRef]
- Gonsior, M.; Peake, B.M.; Cooper, W.T.; Podgorski, D.; Andrilli, J.; Cooper, W.J. Photochemically induced changes in dissolved organic matter identified by ultrahigh resolution fourier transform ion cyclotron resonance mass spectrometry. Environ. Sci. Technol. 2009, 43, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Porcal, P.; Amirbahman, A.; Kopacek, J.; Novak, F.; Norton, S.A. Photochemical release of humic and fulvic acid-bound metals from simulated soil and streamwater. J. Environl. Monit. 2009, 11, 1064–1071. [Google Scholar] [CrossRef] [PubMed]
- Zepp, R.G.; Callaghan, T.V.; Erickson, D.J., III. Interactive effects of ozone depletion and climate change on biogeochemical cycles. Photochem. Photobiol. Sci. 2003, 2, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.L.; Meyer, J.S.; McKnight, D.M. Photooxidation of wetland and riverine dissolved organic matter: Altered copper complexation and organic composition. Hydrobiologia 2007, 579, 95–113. [Google Scholar] [CrossRef]
- Moran, M.A.; Sheldon, W.M.; Zepp, R.G. Carbon loss and optical property changes dring long-term photochemical and biological degradation of estuarine dissolved organic matter. Limnol. Oceangr. 2000, 45, 1254–1264. [Google Scholar] [CrossRef]
- Del Vecchio, R.; Blough, N.V. Photobleaching of chromophoric dissolved organic matter in natural waters: Kinetics and modeling. Mar. Chem. 2002, 78, 231–253. [Google Scholar] [CrossRef]
- Obernosterer, I.; Benner, R. Competition between biological and photochemical processes in the mineralization of dissolved organic carbon. Limnol. Oceangr. 2004, 2004, 117–124. [Google Scholar] [CrossRef]
- Mayer, L.M.; Schick, L.L.; Skorko, K.; Boss, E. Photdissolution of particulate organic matter from sediments. Limnol. Oceangr. 2006, 51, 1064–1071. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, M.; Qin, B.; Feng, S. Photochemical degradation of chromophoric-dissolved organic matter exposed to simulated UV-B and natural solar radiation. Hydrobiologia 2009, 627, 159–168. [Google Scholar] [CrossRef]
- Dalzell, B.J.; Minor, E.C.; Mopper, K.M. Photodegradation of estuarine dissolved organic matter: A multi-method assessment of DOM transformation. Org. Geochem. 2009, 40, 243–257. [Google Scholar] [CrossRef]
- Spierings, J.; Worms, I.A.M.; Mieville, P.; Slaveykova, V.I. Effect of humic substance photoalteration on lead bioavailability to freshwater microalgae. Environ. Sci. Technol. 2011, 45, 345–3458. [Google Scholar] [CrossRef] [PubMed]
- Worms, I.A.; Adenmatten, D.; Mieville, P.; Traber, J.; Slaveykova, V.I. Photo-transformation of pedogenic humic acid and consequences for Cd(ii), Cu(ii) and Pb(ii) speciation and bioavailability to green microalga. Chemosphere 2015, 138, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L.; Moran, M.A.; Sheldon, W.M.; Zepp, R.G.; Opsahl, S. Determination of apparent quantum yeald spectra for the formation of biologically labile photoproducts. Limnol. Oceangr. 2002, 47, 343–352. [Google Scholar] [CrossRef]
- Obernosterer, I.; Sempere, R.; Herndl, G.J. Ultraviolet radiation induces reversal of the bioavailability of dom to marine bacterioplankton. Aquat. Microb. Ecol. 2001, 24, 61–68. [Google Scholar] [CrossRef]
- Xu, H.; Jiang, H. UV-induced photochemical heterogeneity of dissolved and attached organic matter associated with cyanobacterial blooms in a eutrophic freshwater lake. Water Res. 2013, 47, 6506–6515. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.X.; Zhang, Y.Y.; Chen, F.Z.; Chang, Y.G.; Liu, Z.W. Photochemical reactivities of dissolved organic matter (DOM) in a sub-alpine lake revealed by EEM-PARAFAC: An insight into the fate of allochthonous DOM in alpine lakes affected by climate change. Sci. Total Environ. 2016, 568, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.-Z.; Yang, G.-P.; Zhang, H.-H. Photochemical behavior of dissolved and colloidal organic matter in estuarine and oceanic waters. Sci. Total Environ. 2017, 607–608, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Shammi, M.; Pan, X.; Mostofa, K.M.G.; Zhang, D.; Liu, C.-Q. Photo-flocculation of microbial mat extracellular polymeric substances and their transformation into transparent exopolymer particles: Chemical and spectroscopic evidences. Sci. Rep. 2017, 7, 9074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.; Zhao, C.; Mu, S.; Pan, X.; Zhang, D.; Al-Misned, F.A.; Mortuza, M.G. Effects of irradiation and pH on fluorescence properties and flocculation of extracellular polymeric substances from the cyanobacterium Chroococcus minutus. Coll. Surf. B Biointerfaces 2015, 128, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Tonietto, A.E.; Lombardi, A.T.; Vieira, A.A.H. The effects of solar irradiation on copper speciation and organic complexation. J. Braz. Chem. Soc. 2011, 22, 1695–1700. [Google Scholar] [CrossRef] [Green Version]
- Vidali, R.; Remoundaki, E.; Tsezos, M. Humic acids copper binding following their photochemical alteration by simulated solar light. Aquat. Geochem. 2010, 16, 207–218. [Google Scholar] [CrossRef]
- Shank, G.C.; Whitehead, R.F.; Smith, M.L.; Skrabal, S.A.; Kieber, R.J. Photodegradation of strong copper complexing ligands in organic-rich estuarine waters. Limnol. Oceangr. 2006, 51, 884–892. [Google Scholar] [CrossRef]
- Sander, S.; Kim, J.P.; Anderson, B.; Hinter, K.A. Effect of UVB irradiation on Cu2+-binding organic ligands and Cu2+ speciation in alpine lake water of new zealand. Environ. Chem. 2005, 2, 56–62. [Google Scholar] [CrossRef]
- Lindim, C.; Mota, A.M.; Goncalves, M.L.S. Influence of UV-B irradiation in lead speciation from an estuarine sample. Water Res. 2000, 34, 3325. [Google Scholar] [CrossRef]
- Neron, R.; Auclair, J.C.; Fortin, C. Rate of Cd2+ release from dissolved fulvic acid and natural dissolved organic carbon as a function of UVB dose. Environ. Chem. 2006, 3, 433–438. [Google Scholar] [CrossRef]
- Worms, I.A.M.; Szigeti, Z.A.G.; Dubascoux, S.; Lespes, G.; Traber, J.; Sigg, L.; Slaveykova, V.I. Colloidal organic matter from wastewater treatment plant effluents: Characterization and role in metal distribution. Water Res. 2010, 44, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Worms, I.A.M.; Traber, J.; Kistler, D.; Sigg, L.; Slaveykova, V.I. Uptake of Cd(ii) and Pb(ii) by microalgae in presence of colloidal organic matter from wastewater treatment plant effluents. Environ. Pollut. 2010, 158, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Alasonati, E.; Slaveykova, V.I.; Gallard, H.; Croué, J.-P.; Benedetti, M.F. Characterization of the colloidal organic matter from the amazonian basin by asymmetrical flow field-flow fractionation and size exclusion chromatography. Water Res. 2010, 44, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Batley, G.E.; Apte, S.C.; Stauber, J.L. Speciation and bioavailability of trace metals in water: Progress since 1982. Aust. J. Chem. 2004, 57, 903–919. [Google Scholar] [CrossRef]
- Sigg, L.; Black, F.; Buffle, J.; Cao, J.; Cleven, R.; Davison, W.; Galceran, J.; Gunkel, P.; Kalis, E.; Kistler, D.; et al. Comparison of analytical techniques for dynamic trace metal speciation in natural freshwaters. Environ. Sci. Technol. 2006, 40, 1934–1941. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, H.P.; Buffle, J. Chemodynamics of aquatic metal complexes: From small ligands to colloids. Environ. Sci. Technol. 2009, 43, 7175–7183. [Google Scholar] [CrossRef] [PubMed]
- Tercier-Waeber, M.-L.; Hezard, T.; Masson, M.; Schäfer, J.R. In situ monitoring of the diurnal cycling of dynamic metal species in a stream under contrasting photobenthic biofilm activity and hydrological conditions. Environ. Sci. Technol. 2009, 43, 7237–7244. [Google Scholar] [CrossRef] [PubMed]
- Winch, S.; Ridal, J.; Lean, D. Increased metal bioavailability following alteration of freshwater dissolved organic carbon by ultraviolet B radiation exposure. Environ. Toxicol. 2002, 17, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Winch, S.; Lean, D. Comparison of changes in metal toxicity following exposure of water with high dissolved organic carbon content to solar, UV-B and UV-A radiation. Photochem. Photobiol. 2005, 81, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.L.; Meyer, J.S.; Boese, C.J. Toxicity of copper to larval pimephales promelas in the presence of photodegraded natural dissolved organic matter. Can. J. Fish. Aquat. Sci. 2007, 64, 391–401. [Google Scholar] [CrossRef]
- Helbling, E.W.; Zagarese, H.; Kieber, D.J.; Peake, B.M.; Scully, N.M. Reactive oxygen species in aquatic ecosystems. UV effects in aquatic organisms and ecosystems. R. Soc. Chem. 2003, 1, 251–288. [Google Scholar]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a Swiss army knife. Plant. Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Kieber, R.J.; Skrabal, S.A.; Smith, C.; Willey, J.D. Redox speciation of copper in rainwater: Temporal variability and atmospheric deposition. Environ. Sci. Technol. 2004, 38, 3587–3594. [Google Scholar] [CrossRef] [PubMed]
- Deguillaume, L.; Leriche, M.; Desboeufs, K.; Mailhot, G.; George, C.; Chaumerliac, N. Transition metals in atmospheric liquid phases: Sources, reactivity, and sensitive parameters. Chem. Rev. 2005, 105, 3388–3431. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Garg, S.; Waite, T.D. Light-mediated reactive oxygen species generation and iron redox transformations in the presence of exudate from the cyanobacterium Microcystis aeruginosa. Environ. Sci. Technol. 2017, 51, 8384–8395. [Google Scholar] [CrossRef] [PubMed]
- Johannsson, O.E.; Smith, D.S.; Sadauskas-Henrique, H.; Cimprich, G.; Wood, C.M.; Val, A.L. Photo-oxidation processes, properties of doc, reactive oxygen species (ROS), and their potential impacts on native biota and carbon cycling in the Rio Negro (Amazonia, Brazil). Hydrobiologia 2017, 789, 7–29. [Google Scholar] [CrossRef]
- Zeng, J.; Wang, W.X. Temperature and irradiance influences on cadmium and zinc uptake and toxicity in a freshwater cyanobacterium, Microcystis aeruginosa. J. Hazard. Mater. 2011, 190, 922–929. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Juneau, P. Different physiological and photosynthetic responses of three cyanobacterial strains to light and zinc. Aquat. Toxicol. 2016, 170, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Li, Z.-K.; Qiu, B.-S.; Juneau, P. Different responses to high light stress of toxic and non-toxic Microcystis aeruginosa acclimated under two light intensities and zinc concentrations. Toxicol. Environ. Chem. 2013, 95, 1145–1156. [Google Scholar] [CrossRef]
- Cheloni, G.; Cosio, C.; Slaveykova, V.I. Antagonistic and synergistic effects of light irradiation on the effects of copper on Chlamydomonas reinhardtii. Aquat. Toxicol. 2014, 155, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Rai, L.C.; Tyagi, B.; Mallick, N.; Rai, P.K. Interactive effects of UV-B and copper on photosynthetic activity of the cyanobacterium Anabaena doliolum. Environ. Exp. Bot. 1995, 35, 177–185. [Google Scholar] [CrossRef]
- Manson, A.Z.; Jenkins, K.D. Metal detoxification in aquatic organisms. In Metal Speciation and Bioavailability in Aquatic Systems; Tessier, A., Turner, D.R., Eds.; Wiley: Hoboken, NJ, USA, 1992. [Google Scholar]
- Barón, M.; Arellano, J.B.; López Gorgé, J. Copper and photosystem II: A controversial relationship. Physiol. Plant. 1995, 94, 174–180. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.C.; Henriques, F.S. Biochemical, physiological, and structural effects of excess copper in plants. Bot. Rev. 1991, 57, 246–273. [Google Scholar] [CrossRef]
- Juneau, A.; El Berdey, P.; Popovic, R. Pam fluorometry in the determination of the sensitivity of Chlorella vulgaris, Selenastrum capricornutum, and Chlamydomonas reinhardtii to copper. Arch. Environ. Contam. Toxicol. 2002, 42, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Perales-Vela, H.V.; Gonzalez-Moreno, S.; Montes-Horcasitas, C.; Canizares-Villanueva, R.O. Growth, photosynthetic and respiratory responses to sub-lethal copper concentrations in Scenedesmus incrassatulus (chlorophyceae). Chemosphere 2007, 67, 2274–2281. [Google Scholar] [CrossRef] [PubMed]
- Mallick, N.; Mohn, F. Use of chlorophyll fluorescence in metal-stress research: A case study with the green microalga Scenedesmus. Ecotoxicol. Environ. Saf. 2003, 55, 64–69. [Google Scholar] [CrossRef]
- Franklin, N.M.; Stauber, J.L.; Lim, R.P. Development of flow cytometry-based algal bioassays for assessing toxicity of copper in natural waters. Environ. Toxicol. Chem. 2001, 20, 160–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjoudja, S.; Vignoles, C.; Deluchat, V.; Lenain, J.F.; Le Jeune, A.H.; Baudu, M. Short term copper toxicity on Microcystis aeruginosa and Chlorella vulgaris using flow cytometry. Aquat. Toxicol. 2009, 94, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Jamers, A.; Blust, R.; De Coen, W.; Griffin, J.L.; Jones, O.A. Copper toxicity in the microalga Chlamydomonas reinhardtii: An integrated approach. Biometals 2013, 26, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Cirulis, J.T.; Scott, J.A.; Ross, G.M. Management of oxidative stress by microalgae. Can. J. Physiol. Pharmacol. 2013, 91, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.; Sigaud-Kunter, T.C.S.; Leitao, M.A.S.; Okamoto, O.K.; Morse, D.; Colepicolo, P. Heavy metal-induced oxidative stress in algae. J. Phycol. 2003, 39, 1008–1018. [Google Scholar] [CrossRef]
- Von Moos, N.; Slaveykova, V.I. Oxidative stress induced by inorganic nanoparticles in bacteria and aquatic microalgae—State of the art and knowledge gaps. Nanotoxicology 2014, 8, 605–630. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, B.N.; Mehta, S.K.; Amar, A.; Gaur, J.P. Oxidative stress in Scenedesmus sp. During short- and long-term exposure to Cu and Zn. Chemosphere 2006, 62, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Stoiber, T.L.; Shafer, M.M.; Karner Perkins, D.A.; Hemming, J.D.C.; Armstrong, E.D. Analysis of glutathione endpoints for measuring copper stress in Chlamydomonas reinhardtii. Environ. Toxicol. Chem. 2007, 26, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Stoiber, T.L.; Shafer, M.M.; Armstrong, D.E. Induction of reactive oxygen species in Chlamydomonas reinhardtii in response to contrasting trace metal exposures. Environ. Toxicol. 2013, 28, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Szivák, I.; Behra, R.; Sigg, L. Metal-induced reactive oxygen species production in Chlamydomonas reinhardtii (chlorophyceae). J. Phycol. 2009, 45, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Latifi, A.; Ruiz, M.; Zhang, C.C. Oxidative stress in cyanobacteria. FEMS Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witman, G.B. Chlamydomonas phototaxis. Trends Cell Biol. 1993, 3, 403–408. [Google Scholar] [CrossRef]
- Krause, G.H. Photoinhibition of photosynthesis. An evaluation of damaging and protective mechanisms. Physiol. Plant. 1988, 74, 566–574. [Google Scholar] [CrossRef]
- Hooper, M.J.; Ankley, G.T.; Cristol, D.A.; Maryoung, L.A.; Noyes, P.D.; Pinkerton, K.E. Interactions between chemical and climate stressors: A role for mechanistic toxicology in assessing climate change risks. Environ. Toxicol. Chem. 2013, 32, 32–48. [Google Scholar] [CrossRef] [PubMed]
- West, L.J.A.; Li, K.; Greenberg, B.M.; Mierle, G.; Smith, R.E.H. Combined effects of copper and ultraviolet radiation on a microscopic green alga in natural soft lake waters of varying dissolved organic carbon content. Aquat. Toxicol. 2003, 64, 39–52. [Google Scholar] [CrossRef]
- Prasad, S.M.; Zeeshan, M. UV-B radiation and cadmium induced changes in growth, photosynthesis, and antioxidant enzymes of cyanobacterium Plectonema boryanum. Biol. Plant. 2005, 49, 229–236. [Google Scholar] [CrossRef]
- Singh, R.; Srivastava, P.K.; Singh, V.P.; Dubey, G.; Prasad, S.M. Light intensity determines the extent of mercury toxicity in the cyanobacterium Nostoc muscorum. Acta Physiol. Plant. 2011, 34, 1119–1131. [Google Scholar] [CrossRef]
- Singh, P.K.; Rai, S.; Pandey, S.; Agrawal, C.; Shrivastava, A.K.; Kumar, S.; Rai, L.C. Cadmium and UV-B induced changes in proteome and some biochemical attributes of Anabaena sp. Pcc7120. Phykos 2012, 42, 39–50. [Google Scholar]
- Lee, W.M.; An, Y.J. Effects of zinc oxide and titanium dioxide nanoparticles on green algae under visible, UVA, and UVB irradiations: No evidence of enhanced algal toxicity under UV pre-irradiation. Chemosphere 2013, 91, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Navarro, E.; Robinson, C.T.; Behra, R. Increased tolerance to ultraviolet radiation (UVR) and cotolerance to cadmium in UVR-acclimatized freshwater periphyton. Limnol. Oceanogr. 2008, 53, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Corcoll, N.; Bonet, B.; Leira, M.; Montuelle, B.; Tlili, A.; Guasch, H. Light history influences the response of fluvial biofilms to Zn exposure. J. Phycol. 2012, 48, 1411–1423. [Google Scholar] [CrossRef] [PubMed]
- Hader, D.P.; Kumar, H.D.; Smith, R.C.; Worrest, R.C. Effects of solar UV radiation on aquatic ecosystems and interactions with climate change. Photochem. Photobiol. Sci. 2007, 6, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Navarro, E.; Muniz, S.; Korkaric, M.; Wagner, B.; de Caceres, M.; Behra, R. Ultraviolet radiation dose calculation for algal suspensions using UVA and UVB extinction coefficients. J. Photochem. Photobiol. B 2014, 132, 94–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, C.A.; Scott, A.P.; Johnson, A.C.; Panter, G.H.; Sheahan, D.; Roberts, M.; Sumpter, J.P. Principles of sound ecotoxicology. Environ. Sci. Technol. 2014, 48, 3100–3111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Combined effects of multiple stressors in the environment. (A) Illustration of the concepts of additivity, antagonism and synergism; (B) Illustration of combined effects that might occur when the microorganism is pre-exposed to environmental stressor and then to micropollutant, for example toxic trace metal. Three cases can be distinguished: (a) organism acclimates to the environmental stressor, but the acclimation does not interfere with the sensitivity to the trace metal; (b) acclimation confers decreased sensitivity to metal; (c) pre-exposure to the environmental stressor increase the sensitivity to the metal.
Figure 1.
Combined effects of multiple stressors in the environment. (A) Illustration of the concepts of additivity, antagonism and synergism; (B) Illustration of combined effects that might occur when the microorganism is pre-exposed to environmental stressor and then to micropollutant, for example toxic trace metal. Three cases can be distinguished: (a) organism acclimates to the environmental stressor, but the acclimation does not interfere with the sensitivity to the trace metal; (b) acclimation confers decreased sensitivity to metal; (c) pre-exposure to the environmental stressor increase the sensitivity to the metal.

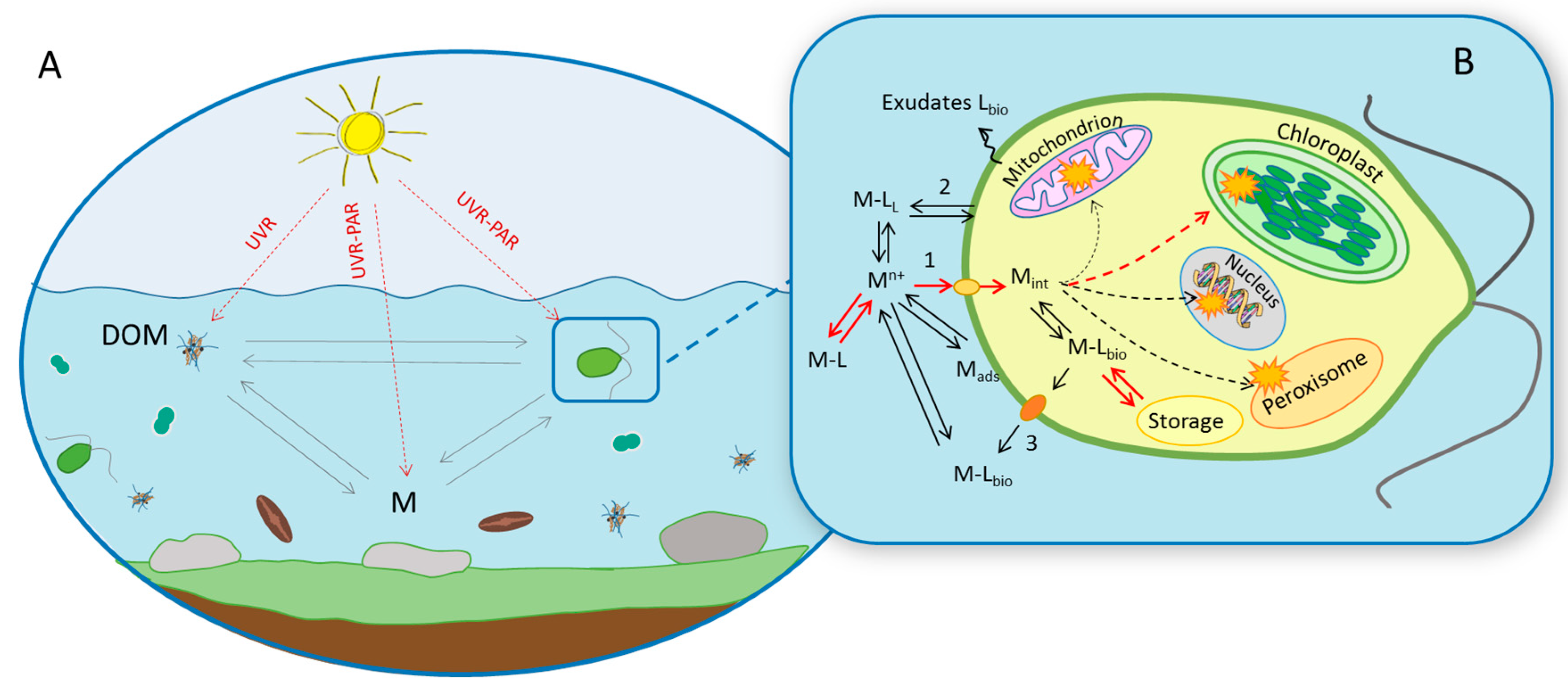
Figure 2.
Effect of the light irradiation on the interactions of trace metal (M) with dissolved organic matter (DOM) and photosynthetic microorganisms. (A) Photo-synthetically active radiation (PAR) and ultraviolet radiation (UVR) influence directly photosynthetic microorganism and their interaction with trace metals and exudates production. UVR might transform DOM and alter chemical speciation and bioavailability of metals. Light might also affect metal speciation directly via photoreactions and changing metal oxidation state. The bioavailable metals might be toxic to photosynthetic organisms and affect their photosynthetic activity. (B) Processes at cell-ambient medium interface determining metal bioavailability and intracellular fate in eukaryotic algae: (1) active transport; (2) passive transport; (3) efflux; Mn+ metal ion; M-L metal complexes with ligands; Mads metal adsorbed to the cell surface; Mint intracellular metal; Lbio biological ligand. To exert an effect both vital and toxic metals have first to interact with the microorganism surface and enter the cells by different mechanisms. Metal ions that reached the intracellular environment might or might not have a toxic action to the microorganism. Cells have evolved several response mechanisms confering protection in case of excess of intracellular metal quota [16]. Metals can be excreted or remain in the intracellular environment where they can be either sequestrated by metal complexing ligands or transported to specific subcellular compartments and stocked in Ca-phosphate granules [17,18]. When the intracellular content exceeds the ability of the cells to maintain homeostasis, toxic effects are observed. Changes in light radiation can affect different toxicokinetics and toxicodynamics processes. The processes known to be affected by light are given in red. No information available concerning the possible effects of light on the other processes.
Figure 2.
Effect of the light irradiation on the interactions of trace metal (M) with dissolved organic matter (DOM) and photosynthetic microorganisms. (A) Photo-synthetically active radiation (PAR) and ultraviolet radiation (UVR) influence directly photosynthetic microorganism and their interaction with trace metals and exudates production. UVR might transform DOM and alter chemical speciation and bioavailability of metals. Light might also affect metal speciation directly via photoreactions and changing metal oxidation state. The bioavailable metals might be toxic to photosynthetic organisms and affect their photosynthetic activity. (B) Processes at cell-ambient medium interface determining metal bioavailability and intracellular fate in eukaryotic algae: (1) active transport; (2) passive transport; (3) efflux; Mn+ metal ion; M-L metal complexes with ligands; Mads metal adsorbed to the cell surface; Mint intracellular metal; Lbio biological ligand. To exert an effect both vital and toxic metals have first to interact with the microorganism surface and enter the cells by different mechanisms. Metal ions that reached the intracellular environment might or might not have a toxic action to the microorganism. Cells have evolved several response mechanisms confering protection in case of excess of intracellular metal quota [16]. Metals can be excreted or remain in the intracellular environment where they can be either sequestrated by metal complexing ligands or transported to specific subcellular compartments and stocked in Ca-phosphate granules [17,18]. When the intracellular content exceeds the ability of the cells to maintain homeostasis, toxic effects are observed. Changes in light radiation can affect different toxicokinetics and toxicodynamics processes. The processes known to be affected by light are given in red. No information available concerning the possible effects of light on the other processes.

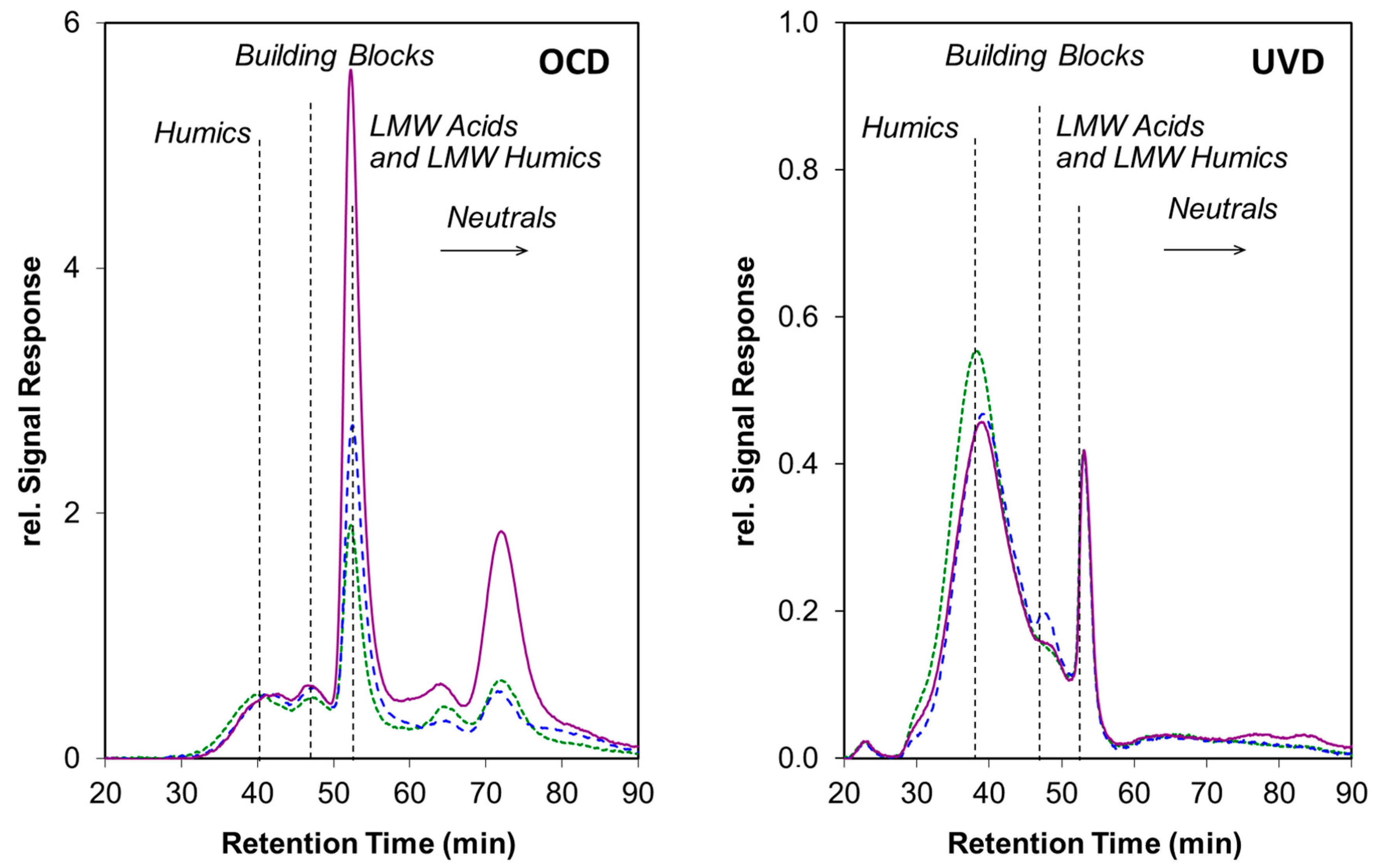
Figure 3.
Liquid chromatography coupled to organic carbon detection (OCD) and UV detection (UVD) of Elliot humic acid irradiated at 8.6 (blue line) and 21 × 106 J m−2 (violet line), illustrating the changes in the chemical composition upon irradiation. Non-irradiated control is given in green. The relative proportion of the building blocks and low molar mass neutral molecules increased significantly with radiation intensity. Further details about the light exposure conditions and DOM transformations can be found in Reference [42].
Figure 3.
Liquid chromatography coupled to organic carbon detection (OCD) and UV detection (UVD) of Elliot humic acid irradiated at 8.6 (blue line) and 21 × 106 J m−2 (violet line), illustrating the changes in the chemical composition upon irradiation. Non-irradiated control is given in green. The relative proportion of the building blocks and low molar mass neutral molecules increased significantly with radiation intensity. Further details about the light exposure conditions and DOM transformations can be found in Reference [42].

Figure 4.
Effect of the DOM nature of Pb speciation in the presence of 10 mg L−1 and irradiated at dose 3.2 × 105 J/m2. Error bars correspond to standard deviations of three replicates. Ion exchange speciation technique (IET) at “equilibrium mode” was used to determine the concentrations of the free lead ions, [Pb2+] in the presence of DOM. The values were normalized to IET measured Pb concentration in non-irradiated DOM [Pb]ITE,prior. AHA: Aldrich humic acid, SRFA: Suwannee River fulvic acid and SRHA: Suwannee River humic acid [41]; Db: organic matter from Duebendorf WWTP effluent, Hw: organic matter from Hinwil WWTP effluent [56,57], Elliot HA: Elliot humic acid, Negro River: Organic matter isolated form Negro River [58].
Figure 4.
Effect of the DOM nature of Pb speciation in the presence of 10 mg L−1 and irradiated at dose 3.2 × 105 J/m2. Error bars correspond to standard deviations of three replicates. Ion exchange speciation technique (IET) at “equilibrium mode” was used to determine the concentrations of the free lead ions, [Pb2+] in the presence of DOM. The values were normalized to IET measured Pb concentration in non-irradiated DOM [Pb]ITE,prior. AHA: Aldrich humic acid, SRFA: Suwannee River fulvic acid and SRHA: Suwannee River humic acid [41]; Db: organic matter from Duebendorf WWTP effluent, Hw: organic matter from Hinwil WWTP effluent [56,57], Elliot HA: Elliot humic acid, Negro River: Organic matter isolated form Negro River [58].

Figure 5.
(A) Comparison of Cu EC50s for C. reinhardtii acclimated to PAR45 (yellow bars) and to PAR395 (red bars). (B) Concentration-response curves for growth inhibition (GI), oxidative stress (OS), membrane damages (MD) and chlorophyll bleaching (CB) of C. reinhardtii exposed to copper. Data points are average values ± standard deviation.
Figure 5.
(A) Comparison of Cu EC50s for C. reinhardtii acclimated to PAR45 (yellow bars) and to PAR395 (red bars). (B) Concentration-response curves for growth inhibition (GI), oxidative stress (OS), membrane damages (MD) and chlorophyll bleaching (CB) of C. reinhardtii exposed to copper. Data points are average values ± standard deviation.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Example of studies investigating the combined effects of metal and light irradiation (Visible and UVR) on phytoplankton species and communities (effects on photosynthesis and/or cell growth).
Table 1.
Example of studies investigating the combined effects of metal and light irradiation (Visible and UVR) on phytoplankton species and communities (effects on photosynthesis and/or cell growth).
| Light | Metal | Organisms | Test Conditions | Effect | Reference |
|---|---|---|---|---|---|
| PAR | Zn Cd | M. aeruginosa | Light (18, 32, 55 µmol photons m−2 s−1) and temperature (15, 24, 30 °C) acclimated cultures exposed to Zn (4.4 × 10−11 M to 1.4 × 10−7 M) and Cd (5.3 × 10−11 M to 3.3 × 10−7 M) for 4 h and 72 h | ↑ | [72] |
| PAR | Hg | Nostoc muscorum | 72 h exposure to Hg under different levels of visible light (10, 70, 130 µmol photons m−2 s−1) | ↑ | [98] |
| PAR | Zn | M. aeruginosa CPCC299 M. aeruginosa CPCC632 | Light (50 and 500 µmol photons m−2 s−1) and Zn (7.8 × 10−7 M and 7.8 × 10−6 M) acclimated cultures exposed to 2000 µmol photons m−2 s−1 for 20 min | ↑ | [74] |
| PAR | Zn | M. aeruginosa CPCC299 M. aeruginosa CPCC632 Synechocystis sp. | Light (50 and 300 µmol photons m−2 s−1) acclimated cultures exposed for 4 h 30 min to Zn (0.78, 7.8 and 39 µM) | ↑ M = Sy | [73] |
| PAR | Zn | Periphytic communities | Light acclimated periphyton (25, 100, 500 µmol photons m−2 s−1) exposed for 6 h to 10, 25, 50, 100, 250, and 500 µmol photons m−2 s−1 and to Zn 1.5 mg L−1 | (LL) ↑↑ (ML,HL) ↑ | [103] |
| PAR, UVA, UVB | Cu | C. reinhardtii | Cultures exposed to light (artificial light: PAR 13.3 W m−2, UVAR 0.29 W m−2 and UVBR 0.08 W m−2; solar simulated light: PAR 230 W m−2, UVAR 9.75 W m−2 and UVBR 0.73 W m−2; solar simulated light with enhanced UVB: PAR 232.2 W m−2, UVAR 10.01 W m−2 and UVBR 2.01 W m−2) and Cu2+ 0.06, 0.6 and 6 µM | (HL) A (UVB) S | [75] |
| UVB | Cd | Plectonema boryanum | Cultures pre-incubated for 2 h with Cd (2 and 8 µM) and exposed to 20 µmol m−2 s−1 and 0.4 W m−2 UVB (0.4 Wm−2) for 30 and 90 min | S | [98] |
| UVB | Cd | Anabaena sp. | Combined exposure to 10 µM Cd and 30 min UVB (9.6 kJ m−2) | A | [100] |
| UVA+ UVB | Cd | C. reinhadtii | 48 h exposure to 100 ± 10 µmol m−2 s−1 with UVR (13 Wm−2 UVA; 0.5 Wm−2 UVB) or without UVR. | S | [7] |
| UVA+ UVB | Cd | Periphytic communities | After 38, 52, and 66 days acclimation to high UVR (686 ± 40 µmol photons s−1 m−2 PAR, 10.88 ± 1.9 mW cm−2 UVA, and 1.03 ± 0.11 mW cm−2 UVB) and low UVR (633 ± 31 µmol photons s−1 m−2 PAR, 3.61 ± 0.48 mWcm−2 UVA, 0.07 ± 0.007 mW cm−2 UVB) periphyton were exposed to 2, 10, 20, 40, and 60 µM Cd for 2 h | Co-T | [102] |
| UV-B | Cu | A. doliolum | Combined exposure to UV-B (12.9 mW m −2 nm −1) and Cu (0.3 and 0.5 mg L−1) for 12 and 25 min | S | [76] |
| PAR UVA UVB | Cu | Pseudokirchneriella subcapitata | 7 days exposure to copper in high DOM or low DOM natural water under PAR; PAR + UVA, PAR + UVA + UVB. 14:10 light dark cycle | = | [97] |
| UVA UVB | Pb Cu Ni Cd | P. subcapitata | Natural fresh water pre irradiated with UVA (16.14 W m−2) and UVB (3.4 W m−2) for up to 20 days and then used to perform toxicity tests | ↑ | [64] |
= No differences observed; ↑ increased sensitivity; S synergism; A antagonism; Co-T co-tolerance; M Microcystis aeruginosa; Sy Synechocystis; LL low light; ML medium light; HL high light.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cheloni, G.; Slaveykova, V.I. Combined Effects of Trace Metals and Light on Photosynthetic Microorganisms in Aquatic Environment. Environments 2018, 5, 81. https://0-doi-org.brum.beds.ac.uk/10.3390/environments5070081
AMA Style
Cheloni G, Slaveykova VI. Combined Effects of Trace Metals and Light on Photosynthetic Microorganisms in Aquatic Environment. Environments. 2018; 5(7):81. https://0-doi-org.brum.beds.ac.uk/10.3390/environments5070081
Chicago/Turabian StyleCheloni, Giulia, and Vera I. Slaveykova. 2018. "Combined Effects of Trace Metals and Light on Photosynthetic Microorganisms in Aquatic Environment" Environments 5, no. 7: 81. https://0-doi-org.brum.beds.ac.uk/10.3390/environments5070081
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.