Fire as a Selection Agent for the Dissemination of Invasive Species: Case Study on the Evolution of Forest Coverage

 , , , and
, , , and
Abstract
:1. Introduction
2. Geophysical and Biophysical Framework of the Area under Study
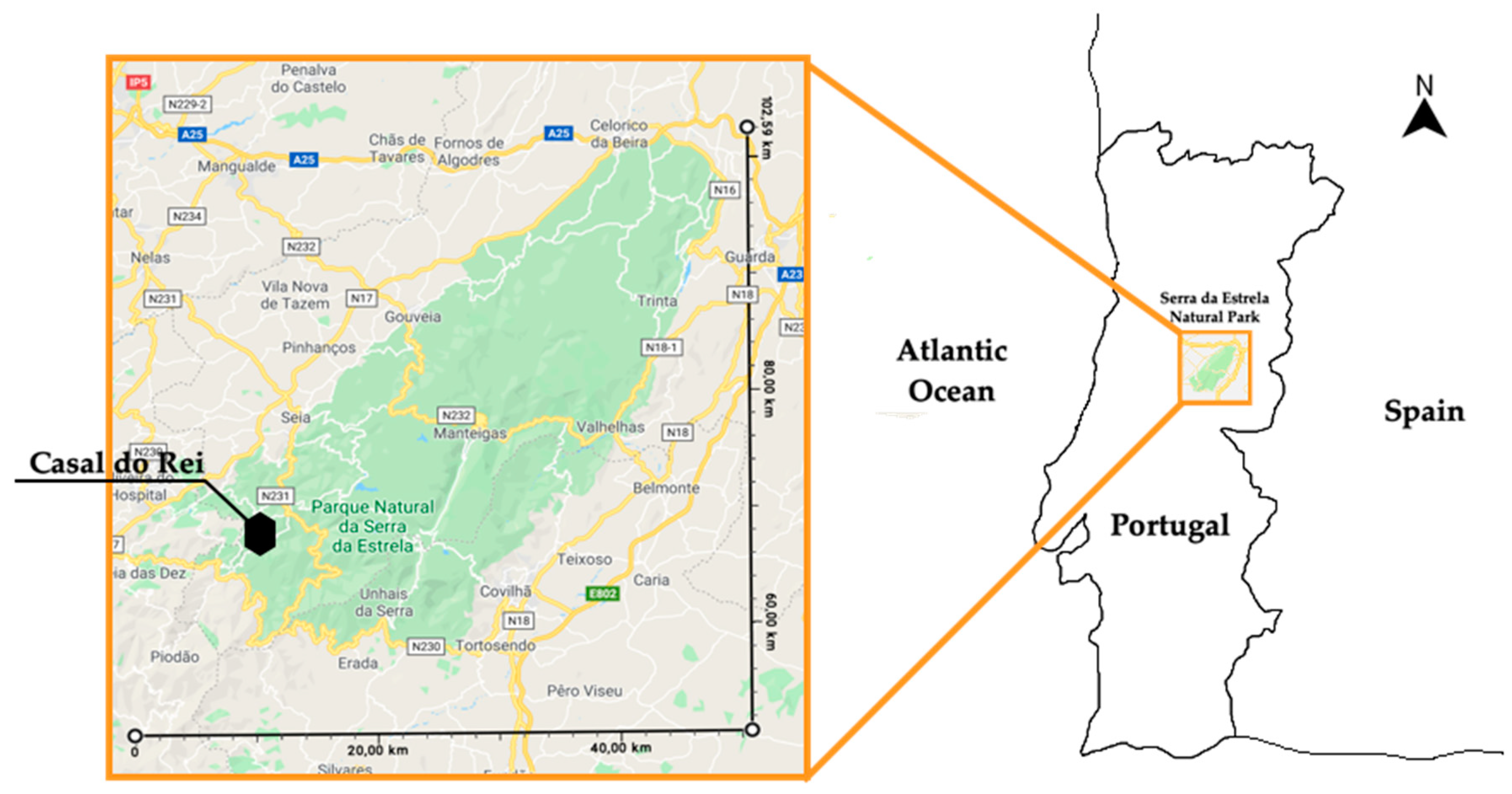
2.1. Location and Limits
2.2. Hydrography
2.3. Geomorphology
2.4. Geology
2.5. Pedology
2.6. Climatology
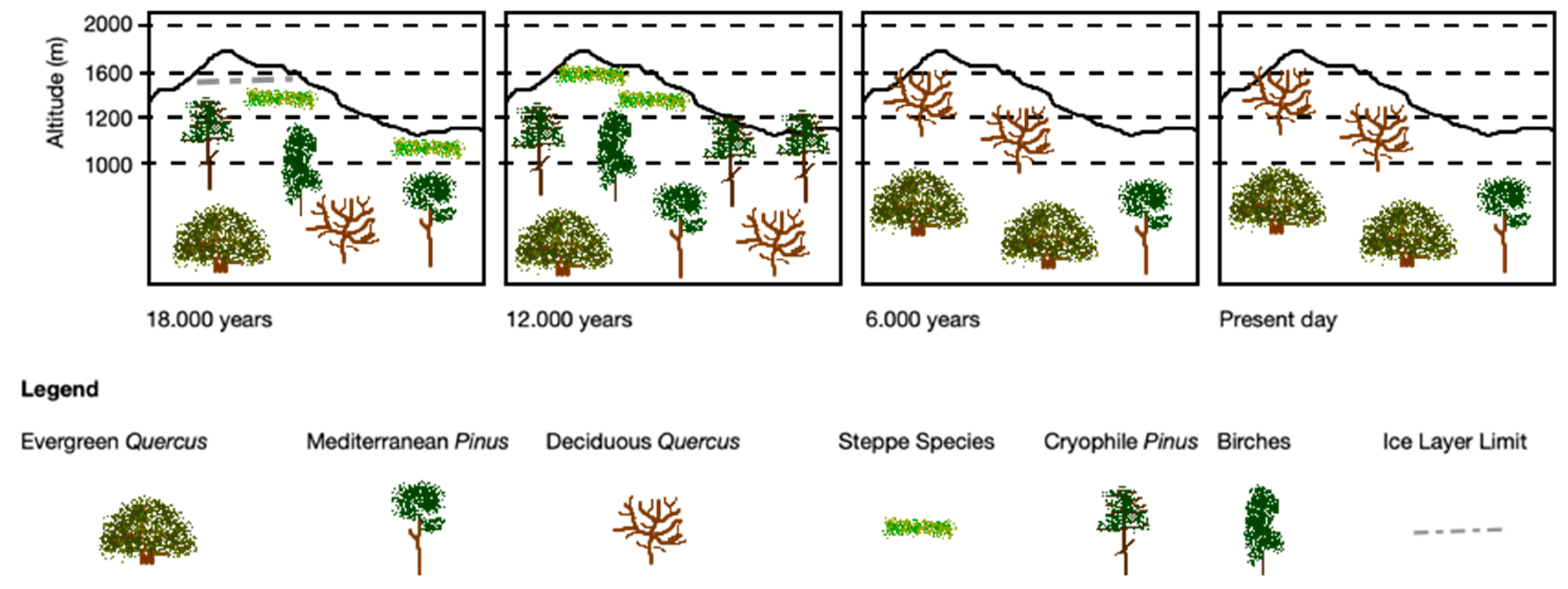
3. Evolution of Vegetation Cover
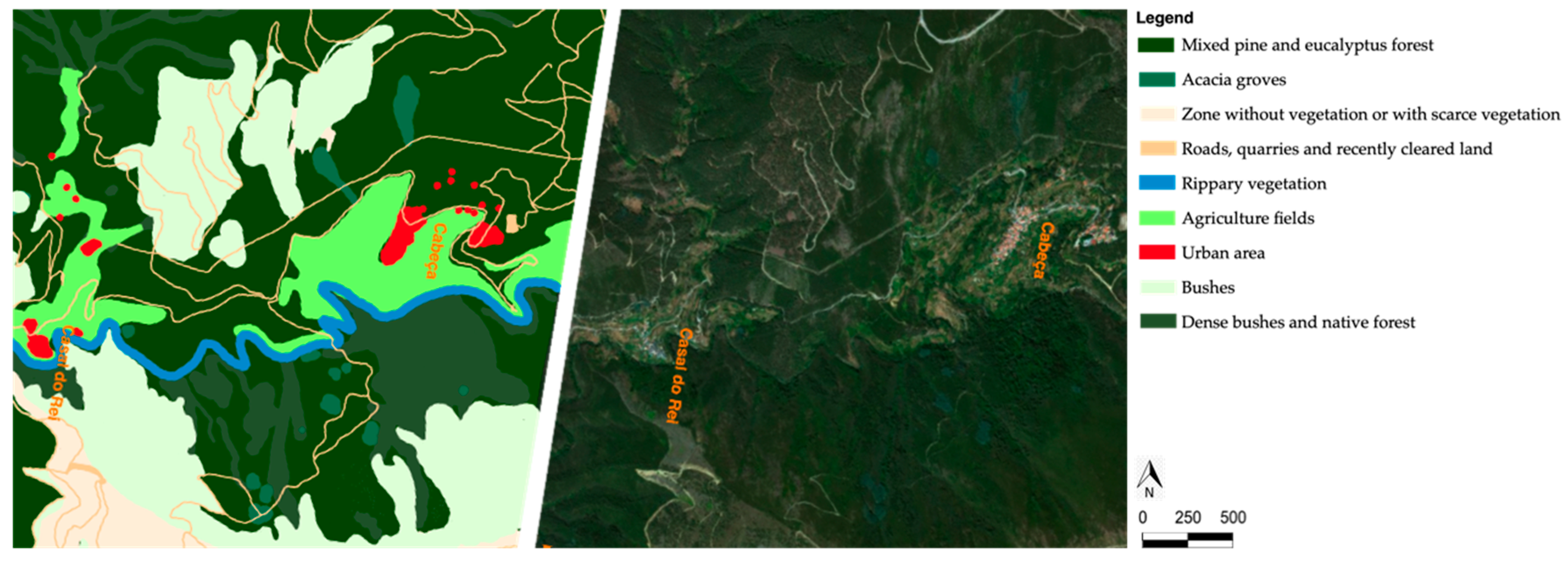
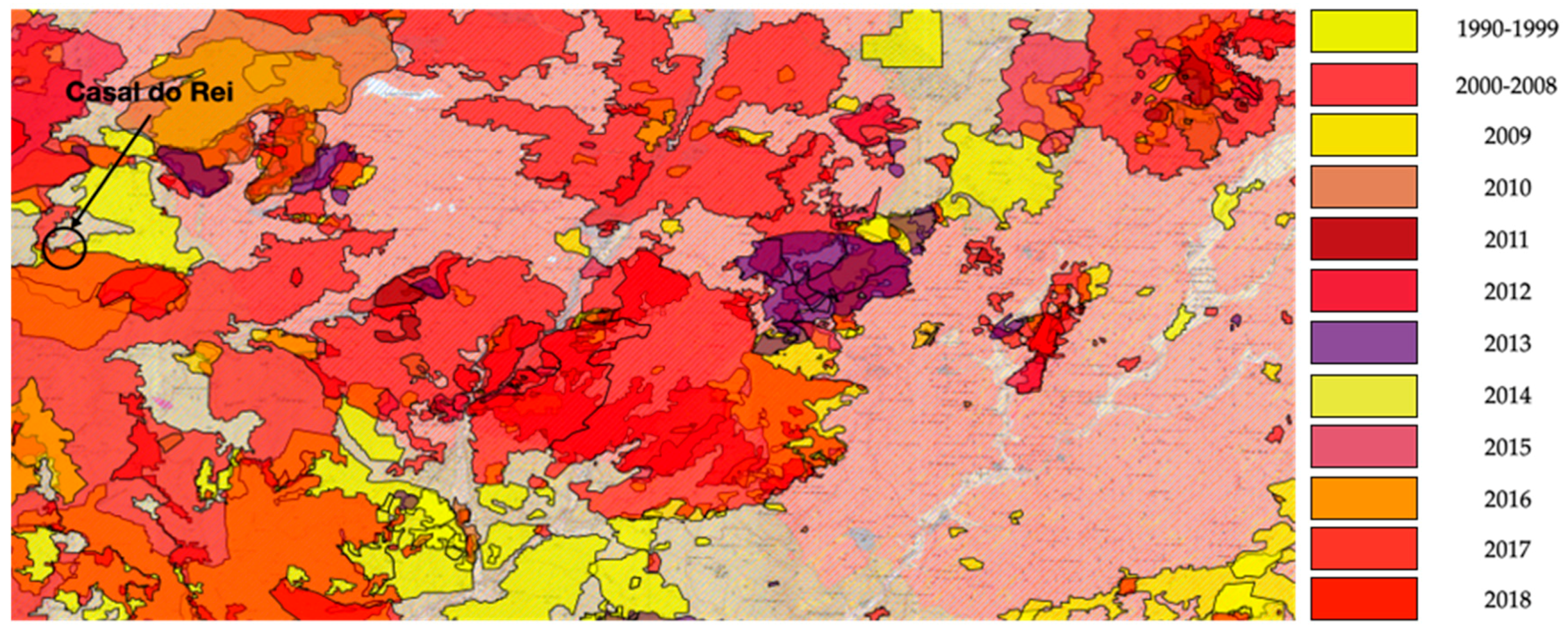
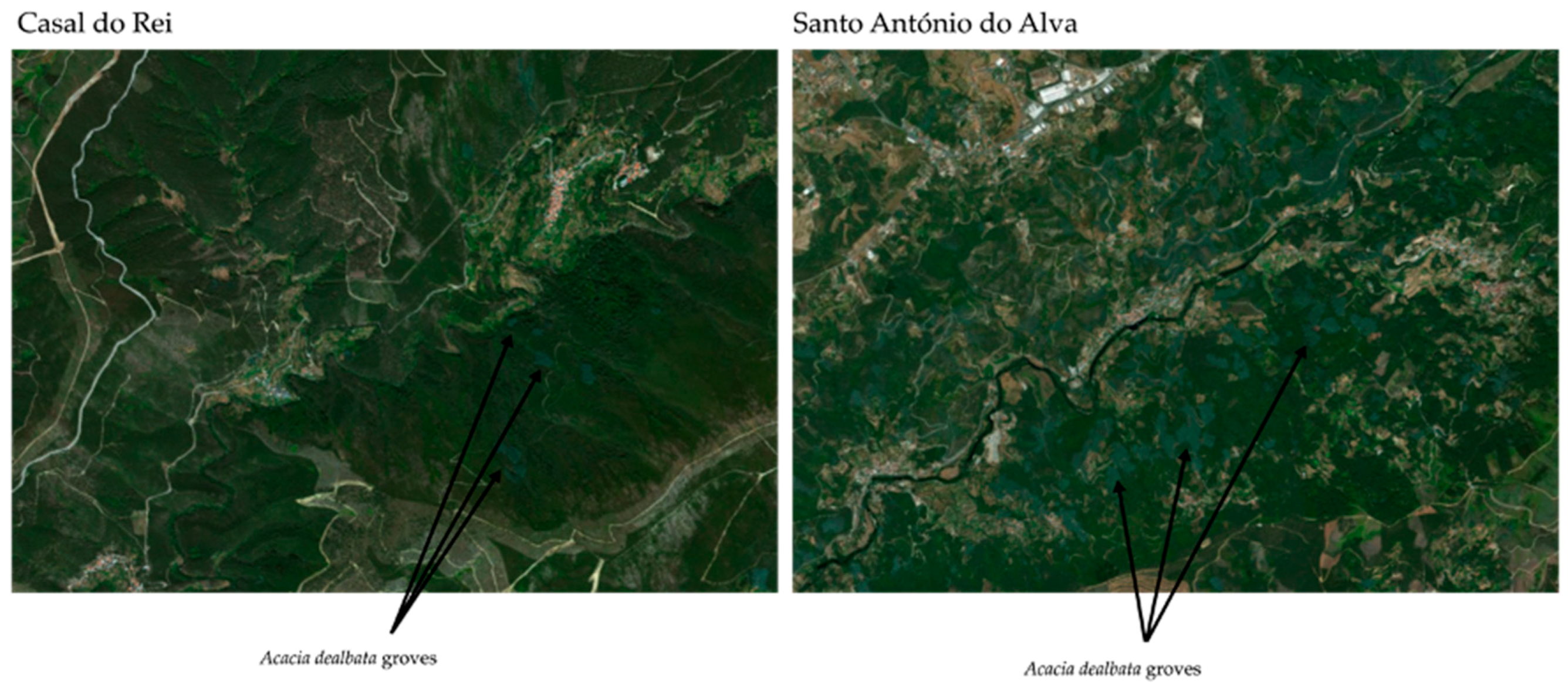
4. Impact of Fire on Vegetation Cover
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xu, Z.; Jiang, Y.; Zhou, G. Nitrogen cycles in terrestrial ecosystems: Climate change impacts and mitigation. Environ. Rev. 2016, 24, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Catullo, R.A.; Llewelyn, J.; Phillips, B.L.; Moritz, C.C. The potential for rapid evolution under anthropogenic climate change. Curr. Biol. 2019, 29, R996–R1007. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.D.; Friess, D.A.; Day, R.H.; MacKenzie, R.A. Impacts of climate change on mangrove ecosystems: A region by region overview. Ecosyst. Health Sustain. 2016, 2, e01211. [Google Scholar] [CrossRef] [Green Version]
- Saxe, H.; Cannell, M.G.; Johnsen, Ø.; Ryan, M.G.; Vourlitis, G. Tree and forest functioning in response to global warming. New Phytol. 2001, 149, 369–399. [Google Scholar] [CrossRef]
- Funk, C.; Harrison, L.; Alexander, L.; Peterson, P.; Behrangi, A.; Husak, G. Exploring trends in wet-season precipitation and drought indices in wet, humid and dry regions. Environ. Res. Lett. 2019, 14, 115002. [Google Scholar] [CrossRef]
- Ciceu, A.; Popa, I.; Leca, S.; Pitar, D.; Chivulescu, S.; Badea, O. Climate change effects on tree growth from Romanian forest monitoring Level II plots. Sci. Total Environ. 2020, 698, 134129. [Google Scholar] [CrossRef]
- Runting, R.K.; Bryan, B.A.; Dee, L.E.; Maseyk, F.J.; Mandle, L.; Hamel, P.; Wilson, K.A.; Yetka, K.; Possingham, H.P.; Rhodes, J.R. Incorporating climate change into ecosystem service assessments and decisions: A review. Glob. Chang. Biol. 2017, 23, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Williamson, C.E.; Neale, P.J.; Hylander, S.; Rose, K.C.; Figueroa, F.L.; Robinson, S.A.; Häder, D.-P.; Wängberg, S.-Å.; Worrest, R.C. The interactive effects of stratospheric ozone depletion, UV radiation, and climate change on aquatic ecosystems. Photochem. Photobiol. Sci. 2019, 18, 717–746. [Google Scholar] [CrossRef] [Green Version]
- Greaver, T.; Clark, C.; Compton, J.; Vallano, D.; Talhelm, A.; Weaver, C.; Band, L.; Baron, J.S.; Davidson, E.; Tague, C. Key ecological responses to nitrogen are altered by climate change. Nat. Clim. Chang. 2016, 6, 836–843. [Google Scholar] [CrossRef]
- Thom, D.; Rammer, W.; Seidl, R. Disturbances catalyze the adaptation of forest ecosystems to changing climate conditions. Glob. Chang. Biol. 2017, 23, 269–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macfadyen, S.; McDonald, G.; Hill, M.P. From species distributions to climate change adaptation: Knowledge gaps in managing invertebrate pests in broad-acre grain crops. Agric. Ecosyst. Environ. 2018, 253, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Millington, R.; Cox, P.M.; Moore, J.R.; Yvon-Durocher, G. Modelling ecosystem adaptation and dangerous rates of global warming. Emerg. Top. Life Sci. 2019, 3, 221–231. [Google Scholar]
- Prober, S.M.; Potts, B.M.; Bailey, T.; Byrne, M.; Dillon, S.; Harrison, P.A.; Hoffmann, A.A.; Jordan, R.; McLean, E.H.; Steane, D.A. Climate adaptation and ecological restoration in eucalypts. Proc. R. Soc. Vic. 2016, 128, 40–53. [Google Scholar] [CrossRef] [Green Version]
- Razgour, O.; Forester, B.; Taggart, J.B.; Bekaert, M.; Juste, J.; Ibáñez, C.; Puechmaille, S.J.; Novella-Fernandez, R.; Alberdi, A.; Manel, S. Considering adaptive genetic variation in climate change vulnerability assessment reduces species range loss projections. Proc. Natl. Acad. Sci. USA 2019, 116, 10418–10423. [Google Scholar] [CrossRef] [Green Version]
- Batllori, E.; De Cáceres, M.; Brotons, L.; Ackerly, D.D.; Moritz, M.A.; Lloret, F. Compound fire-drought regimes promote ecosystem transitions in Mediterranean ecosystems. J. Ecol. 2019, 107, 1187–1198. [Google Scholar] [CrossRef]
- Paraskevopoulou, A.T.; Nektarios, P.A.; Kotsiris, G. Post-fire attitudes and perceptions of people towards the landscape character and development in the rural Peloponnese, a case study of the traditional village of Leontari, Arcadia, Greece. J. Environ. Manag. 2019, 241, 567–574. [Google Scholar] [CrossRef]
- Wolpert, F.; Quintas-Soriano, C.; Plieninger, T. Exploring land-use histories of tree-crop landscapes: A cross-site comparison in the Mediterranean Basin. Sustain. Sci. 2020, 1–17. [Google Scholar] [CrossRef]
- Batllori, E.; De Cáceres, M.; Brotons, L.; Ackerly, D.D.; Moritz, M.A.; Lloret, F. Cumulative effects of fire and drought in Mediterranean ecosystems. Ecosphere 2017, 8, e01906. [Google Scholar] [CrossRef] [Green Version]
- Bazan, G.; Barba, A.C.; Rotolo, A.; Marino, P. Geobotanical approach to detect land-use change of a Mediterranean landscape: A case study in Central-Western Sicily. GeoJournal 2019, 84, 795–811. [Google Scholar] [CrossRef]
- Ojeda, F.; Midgley, J.; Pauw, A.; Lavola, A.; Casimiro-Soriguer, R.; Hattas, D.; Segarra-Moragues, J.G.; Julkunen-Tiitto, R. Flower colour divergence is associated with post-fire regeneration dimorphism in the fynbos heath Erica coccinea subsp. coccinea (Ericaceae). Evol. Ecol. 2019, 33, 345–367. [Google Scholar] [CrossRef]
- Kane, J.M.; Varner, J.M.; Saunders, M.R. Resurrecting the Lost Flames of American Chestnut. Ecosystems 2019, 22, 995–1006. [Google Scholar] [CrossRef]
- Soler, M.; Úbeda, X. Evaluation of fire severity via analysis of photosynthetic pigments: Oak, eucalyptus and cork oak leaves in a Mediterranean forest. J. Environ. Manag. 2018, 206, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Matusick, G.; Ruthrof, K.X.; Fontaine, J.B.; Hardy, G.E.S.J. Eucalyptus forest shows low structural resistance and resilience to climate change-type drought. J. Veg. Sci. 2016, 27, 493–503. [Google Scholar] [CrossRef] [Green Version]
- Franco-Manchón, I.; Salo, K.; Oria-de-Rueda, J.A.; Bonet, J.A.; Martín-Pinto, P. Are wildfires a threat to fungi in European pinus forests? A case study of Boreal and Mediterranean forests. Forests 2019, 10, 309. [Google Scholar] [CrossRef] [Green Version]
- Gea-Izquierdo, G.; Férriz, M.; García-Garrido, S.; Aguín, O.; Elvira-Recuenco, M.; Hernandez-Escribano, L.; Martin-Benito, D.; Raposo, R. Synergistic abiotic and biotic stressors explain widespread decline of Pinus pinaster in a mixed forest. Sci. Total Environ. 2019, 685, 963–975. [Google Scholar] [CrossRef]
- Dodet, M.; Collet, C. When should exotic forest plantation tree species be considered as an invasive threat and how should we treat them? Biol. Invasions 2012, 14, 1765–1778. [Google Scholar] [CrossRef]
- Moser, W.K.; Barnard, E.L.; Billings, R.F.; Crocker, S.J.; Dix, M.E.; Gray, A.N.; Ice, G.G.; Kim, M.-S.; Reid, R.; Rodman, S.U. Impacts of nonnative invasive species on US forests and recommendations for policy and management. J. For. 2009, 107, 320–327. [Google Scholar]
- Martins, F.; Alegria, C.; Gil, A. Mapping invasive alien Acacia dealbata Link using ASTER multispectral imagery: A case study in central-eastern of Portugal. For. Syst. 2016, 25, 13. [Google Scholar] [CrossRef] [Green Version]
- Coumou, D.; Rahmstorf, S. A decade of weather extremes. Nat. Clim. Chang. 2012, 2, 491–496. [Google Scholar] [CrossRef]
- Alexander, L.V.; Zhang, X.; Peterson, T.C.; Caesar, J.; Gleason, B.; Klein Tank, A.; Haylock, M.; Collins, D.; Trewin, B.; Rahimzadeh, F. Global observed changes in daily climate extremes of temperature and precipitation. J. Geophys. Res. Space Phys. 2006, 111. [Google Scholar] [CrossRef] [Green Version]
- Kharin, V.V.; Zwiers, F.W.; Zhang, X.; Hegerl, G.C. Changes in temperature and precipitation extremes in the IPCC ensemble of global coupled model simulations. J. Clim. 2007, 20, 1419–1444. [Google Scholar] [CrossRef] [Green Version]
- Feio, M.; Norris, R.; Graça, M.; Nichols, S. Water quality assessment of Portuguese streams: Regional or national predictive models? Ecol. Indic. 2009, 9, 791–806. [Google Scholar] [CrossRef] [Green Version]
- Migoń, P.; Vieira, G. Granite geomorphology and its geological controls, Serra da Estrela, Portugal. Geomorphology 2014, 226, 1–14. [Google Scholar] [CrossRef]
- Martins, A.N. Sustainable Design, Cultural Landscapes and Heritage Parks; the Case of the Mondego River. Sustain. Dev. 2016, 24, 330–343. [Google Scholar] [CrossRef]
- Ferreira, V.; Graça, M.A.; Feio, M.J.; Mieiro, C. Water quality in the Mondego river basin: Pollution and habitat heterogeneity. Limnetica 2004, 23, 295–306. [Google Scholar]
- Martins, A.A.; Cunha, P.P.; Rosina, P.; Osterbeek, L.; Cura, S.; Grimaldi, S.; Gomes, J.; Buylaert, J.-P.; Murray, A.S.; Matos, J. Geoarchaeology of Pleistocene open-air sites in the Vila Nova da Barquinha-Santa Cita area (Lower Tejo River basin, central Portugal). Proc. Geol. Assoc. 2010, 121, 128–140. [Google Scholar] [CrossRef]
- Mora, C. A synthetic map of the climatopes of the Serra da Estrela (Portugal). J. Maps 2010, 6, 591–608. [Google Scholar] [CrossRef] [Green Version]
- Vieira, G. Combined numerical and geomorphological reconstruction of the Serra da Estrela plateau icefield, Portugal. Geomorphology 2008, 97, 190–207. [Google Scholar] [CrossRef]
- Vieira, G.T.; Mora, C.; Ramos, M. Ground temperature regimes and geomorphological implications in a Mediterranean mountain (Serra da Estrela, Portugal). Geomorphology 2003, 52, 57–72. [Google Scholar] [CrossRef]
- Palacios, D.; Andrés, N.; Marcos, J.; Vázquez-Selem, L. Maximum glacial advance and deglaciation of the Pinar Valley (Sierra de Gredos, Central Spain) and its significance in the Mediterranean context. Geomorphology 2012, 177, 51–61. [Google Scholar] [CrossRef]
- Marques, J.E.; Marques, J.M.; Chaminé, H.I.; Carreira, P.M.; Fonseca, P.E.; Santos, F.A.M.; Moura, R.; Samper, J.; Pisani, B.; Teixeira, J. Conceptualizing a mountain hydrogeologic system by using an integrated groundwater assessment (Serra da Estrela, Central Portugal): A review. Geosci. J. 2013, 17, 371–386. [Google Scholar] [CrossRef]
- Sant’Ovaia, H.; Olivier, P.; Ferreira, N.; Noronha, F.; Leblanc, D. Magmatic structures and kinematics emplacement of the Variscan granites from Central Portugal (Serra da Estrela and Castro Daire areas). J. Struct. Geol. 2010, 32, 1450–1465. [Google Scholar] [CrossRef]
- Sá, A.A.; Gutiérrez-Marco, J.C.; Meireles, C.; García-Bellido, D.C.; Rábano, I. A revised correlation of Lower Ordovician sedimentary rocks in the Central Iberian Zone (Portugal and Spain). In STRATI 2013; Springer: Berlin/Heidelberg, Germany, 2014; pp. 441–445. [Google Scholar]
- Meireles, C.; Castro, P.; Ferreira, N. Evidências cartográficas, litoestratigráficas e estruturais sobre a presença de discordância cadomiana intra Grupo das Beiras. Comunicações Geológicas 2014, 101, 495–498. [Google Scholar]
- Marques, J.E.; Samper, J.; Pisani, B.; Alvares, D.; Carvalho, J.M.; Chaminé, H.I.; Marques, J.; Vieira, G.; Mora, C.; Borges, F.S. Evaluation of water resources in a high-mountain basin in Serra da Estrela, Central Portugal, using a semi-distributed hydrological model. Environ. Earth Sci. 2011, 62, 1219–1234. [Google Scholar] [CrossRef]
- Marques, J.E.; Duarte, J.M.; Constantino, A.T.; Martins, A.A.; Aguiar, C.; Rocha, F.T.; Inácio, M.; Marques, J.M.; Chaminé, H.I.; Teixeira, J. Vadose zone characterisation of a hydrogeologic system in a mountain region: Serra da Estrela case study (Central Portugal). In Aquifer Systems Management: Darcy’s Legacy in a World of Impending Water Shortage: Selected Papers on Hydrogeology; CRC Press: Boca Raton, FL, USA, 2007; p. 207. [Google Scholar]
- Connor, S.E.; Araújo, J.; van der Knaap, W.O.; van Leeuwen, J.F. A long-term perspective on biomass burning in the Serra da Estrela, Portugal. Quat. Sci. Rev. 2012, 55, 114–124. [Google Scholar] [CrossRef]
- Van der Knaap, W.; Van Leeuwen, J. Holocene vegetation succession and degradation as responses to climatic change and human activity in the Serra de Estrela, Portugal. Rev. Palaeobot. Palynol. 1995, 89, 153–211. [Google Scholar] [CrossRef]
- Meireles, C.; Pinto-Gomes, C.; Cano, E. Approach to climatophilous vegetation series of Serra da Estrela (Portugal) Approche des séries de végétation climatophiles de la Serra da Estrela (Portugal). Acta Bot. Gall. 2012, 159, 283–287. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Belmonte, D.; Cantó, P.; Fernández-González, F.; Fuente, V.; Moreno, J.; Sánchez-Mata, D.; GARCÍA SANCHO, L. Piornales, enebrales y pinares oromediterráneos (Pino-Cytision oromediterranei) en el Sistema Central. Lazaroa 1987, 7, 93–124. [Google Scholar]
- Ramos, C.; Ramos-Pereira, A. Landscapes of Portugal: Paleogeographic Evolution, Tectonics and Geomorphology. In Landscapes and Landforms of Portugal; Springer: Berlin/Heidelberg, Germany, 2020; pp. 3–31. [Google Scholar]
- Mugica, F.F.; Antón, M.G.; Ollero, H.S. Vegetation dynamics and human impact in the Sierra de Guadarrama, Central System, Spain. Holocene 1998, 8, 69–82. [Google Scholar] [CrossRef]
- Rubiales, J.M.; García-Amorena, I.; Hernández, L.; Génova, M.; Martínez, F.; Manzaneque, F.G.; Morla, C. Late Quaternary dynamics of pinewoods in the Iberian Mountains. Quat. Sci. Rev. 2010, 162, 476–491. [Google Scholar] [CrossRef]
- Rubiales, J.; García-Amorena, I.; Génova, M.; Manzaneque, F.G.; Morla, C. The Holocene history of highland pine forests in a submediterranean mountain: The case of Gredos mountain range (Iberian Central range, Spain). Quat. Sci. Rev. 2007, 26, 1759–1770. [Google Scholar] [CrossRef]
- Figueiral, I.; Carcaillet, C. A review of Late Pleistocene and Holocene biogeography of highland Mediterranean pines (Pinus type sylvestris) in Portugal, based on wood charcoal. Quat. Sci. Rev. 2005, 24, 2466–2476. [Google Scholar] [CrossRef]
- Rubiales, J.M.; Génova, M. Late Holocene pinewoods persistence in the Gredos Mountains (central Spain) inferred from extensive megafossil evidence. Quat. Rev. 2015, 84, 12–20. [Google Scholar] [CrossRef]
- Nunes, A.N. Regional variability and driving forces behind forest fires in Portugal an overview of the last three decades (1980–2009). Appl. Geogr. 2012, 34, 576–586. [Google Scholar] [CrossRef]
- Nunes, L.J.R.; Raposo, M.; Meireles, C.I.R.; Gomes, C.J.P.; Nuno, M.C.; Almeida, R. The propagation of invasive species as a consequence of rural Fires: A case study in Casal do Rei (Seia). In Proceedings of the IV PhD Students Meeting in Environmental and Agriculture, Évora, Portugal, 11–12 November 2019; p. 62. [Google Scholar]
- Ferreira-Leite, F.; Lourenço, L.; Bento-Gonçalves, A. Large forest fires in mainland Portugal, brief characterization. Méditerranée. Revue géographique des pays méditerranéens/J. Mediterr. Geogr. 2013, 121, 53–65. [Google Scholar] [CrossRef]
- Nunes, A. Incêndios florestais no Parque Natural da Serra da Estrela: Quadro físico-geográfico da sua ocorrência. Cadernos de Geografia 2001, 20, 93–112. [Google Scholar] [CrossRef]
- Directive, H. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–50. [Google Scholar]
- Fernández-García, V.; Fulé, P.Z.; Marcos, E.; Calvo, L. The role of fire frequency and severity on the regeneration of Mediterranean serotinous pines under different environmental conditions. For. Ecol. Manag. 2019, 444, 59–68. [Google Scholar] [CrossRef]
- Jansen, J.; Diemont, H. Implementing Natura 2000 in farmed landscapes: The Serra da Estrela, Portugal. Ekológia 2011, 30, 199–215. [Google Scholar] [CrossRef]
- Chirino, E.; Vilagrosa, A.; Cortina, J.; Valdecantos, A.; Fuentes, D.; Trubat, R.; Luis, V.C.; Puértolas, J.; Bautista, S.; Baeza, J. Ecological restoration in degraded drylands: The need to improve the seedling quality and site conditions in the field. In Forest Management; Nova Publisher: New York, NY, USA, 2009; pp. 85–158. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Data | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Viseu Location: 40°39’ N 7°54’ W Altitude: 443 m | Taverage (°C) | 6.9 | 8.4 | 10.3 | 11.5 | 14.3 | 18.4 | 21.4 | 21.1 | 18.8 | 14.2 | 10.2 | 8.1 |
| Tmax (°C) | 11.6 | 13.2 | 15.9 | 17.1 | 20.2 | 25.4 | 29.2 | 29.2 | 25.9 | 19.8 | 15.0 | 12.4 | |
| Tmin (°C) | 2.2 | 3.5 | 4.6 | 6.0 | 8.5 | 11.5 | 13.5 | 13.0 | 11.7 | 8.5 | 5.3 | 3.8 | |
| Paverage (mm) | 155.7 | 133.6 | 74.8 | 105.2 | 95.9 | 66.0 | 50.5 | 30.6 | 75.2 | 133.2 | 135.9 | 195.4 | |
| Guarda Location: 40°31’ N 7°15’ W Altitude: 1019 m | Taverage (°C) | 4.0 | 5.2 | 7.2 | 8.0 | 11.6 | 15.9 | 19.1 | 19.4 | 16.4 | 11.3 | 7.5 | 4.9 |
| Tmax (°C) | 6.8 | 8.2 | 10.8 | 12.0 | 15.7 | 20.5 | 24.5 | 24.6 | 21.2 | 14.8 | 10.3 | 7.4 | |
| Tmin (°C) | 1.2 | 2.0 | 3.3 | 4.2 | 7.4 | 11.0 | 13.7 | 13.7 | 11.9 | 8.1 | 4.8 | 2.5 | |
| Paverage (mm) | 100.1 | 87.4 | 53.3 | 83.1 | 84.9 | 43.4 | 19.6 | 11.0 | 47.5 | 103.2 | 106.7 | 141.8 | |
| Castelo Branco Location: 39°50’ N 7°28’ W Altitude: 386 m | Taverage (°C) | 7.9 | 9.6 | 12.7 | 13.1 | 16.8 | 21.0 | 25.0 | 24.4 | 21.3 | 16.3 | 11.7 | 9.0 |
| Tmax (°C) | 11.8 | 14.0 | 18.0 | 18.6 | 22.3 | 27.3 | 32.1 | 31.6 | 27.3 | 21.0 | 15.7 | 112.5 | |
| Tmin (°C) | 3.9 | 5.2 | 7.5 | 8.0 | 11.2 | 14.6 | 17.9 | 17.2 | 15.2 | 11.6 | 7.7 | 5.6 | |
| Paverage (mm) | 108.0 | 56.7 | 36.9 | 58.1 | 65.1 | 25.2 | 8.9 | 8.4 | 36.5 | 105.5 | 118.8 | 128.2 | |
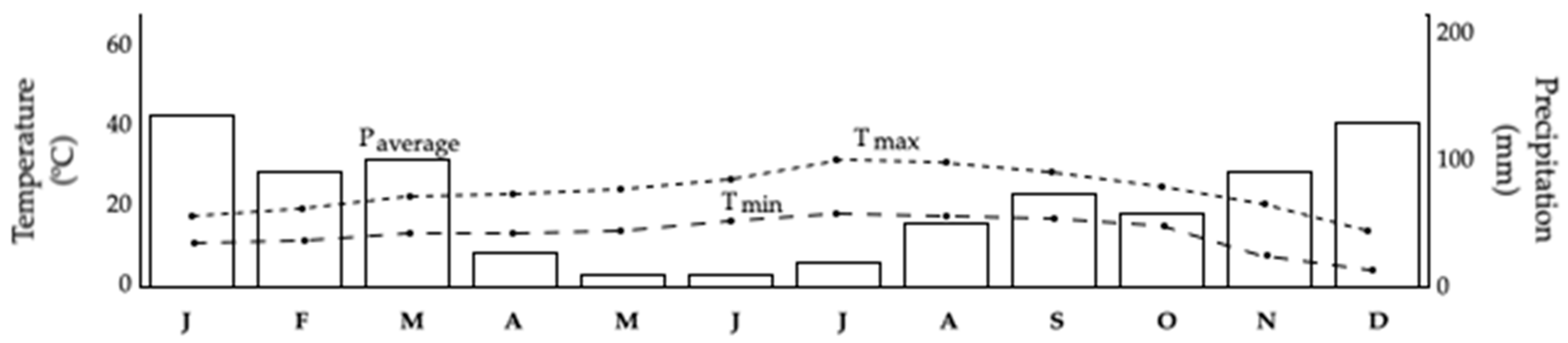
| Casal do Rei Location: 40°31’ N 7°55’ W Altitude: 470 m | Taverage (°C) | 8.5 | 9.0 | 11.5 | 12.5 | 16.0 | 19.0 | 22.0 | 22.0 | 20.0 | 16.0 | 11.5 | 9.0 |
| Tmax (°C) | 12.0 | 13.0 | 16.0 | 17.0 | 21.0 | 25.0 | 29.0 | 29.0 | 26.0 | 20.0 | 15.0 | 13.0 | |
| Tmin (°C) | 5.0 | 5.0 | 7.0 | 8.0 | 11.0 | 13.0 | 15.0 | 15.0 | 14.0 | 12.0 | 8.0 | 6.0 | |
| Paverage (mm) | 123 | 90 | 77 | 81 | 60 | 22 | 7 | 10 | 39 | 118 | 114 | 131 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes, L.J.R.; Raposo, M.A.M.; Meireles, C.I.R.; Pinto Gomes, C.J.; Ribeiro, N.M.C.A. Fire as a Selection Agent for the Dissemination of Invasive Species: Case Study on the Evolution of Forest Coverage. Environments 2020, 7, 57. https://0-doi-org.brum.beds.ac.uk/10.3390/environments7080057
Nunes LJR, Raposo MAM, Meireles CIR, Pinto Gomes CJ, Ribeiro NMCA. Fire as a Selection Agent for the Dissemination of Invasive Species: Case Study on the Evolution of Forest Coverage. Environments. 2020; 7(8):57. https://0-doi-org.brum.beds.ac.uk/10.3390/environments7080057
Chicago/Turabian StyleNunes, Leonel J. R., Mauro A. M. Raposo, Catarina I. R. Meireles, Carlos J. Pinto Gomes, and Nuno M. C. Almeida Ribeiro. 2020. "Fire as a Selection Agent for the Dissemination of Invasive Species: Case Study on the Evolution of Forest Coverage" Environments 7, no. 8: 57. https://0-doi-org.brum.beds.ac.uk/10.3390/environments7080057