
The Impact of Rural Fires on the Development of Invasive Species: Analysis of a Case Study with Acacia dealbata Link. in Casal do Rei (Seia, Portugal)

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
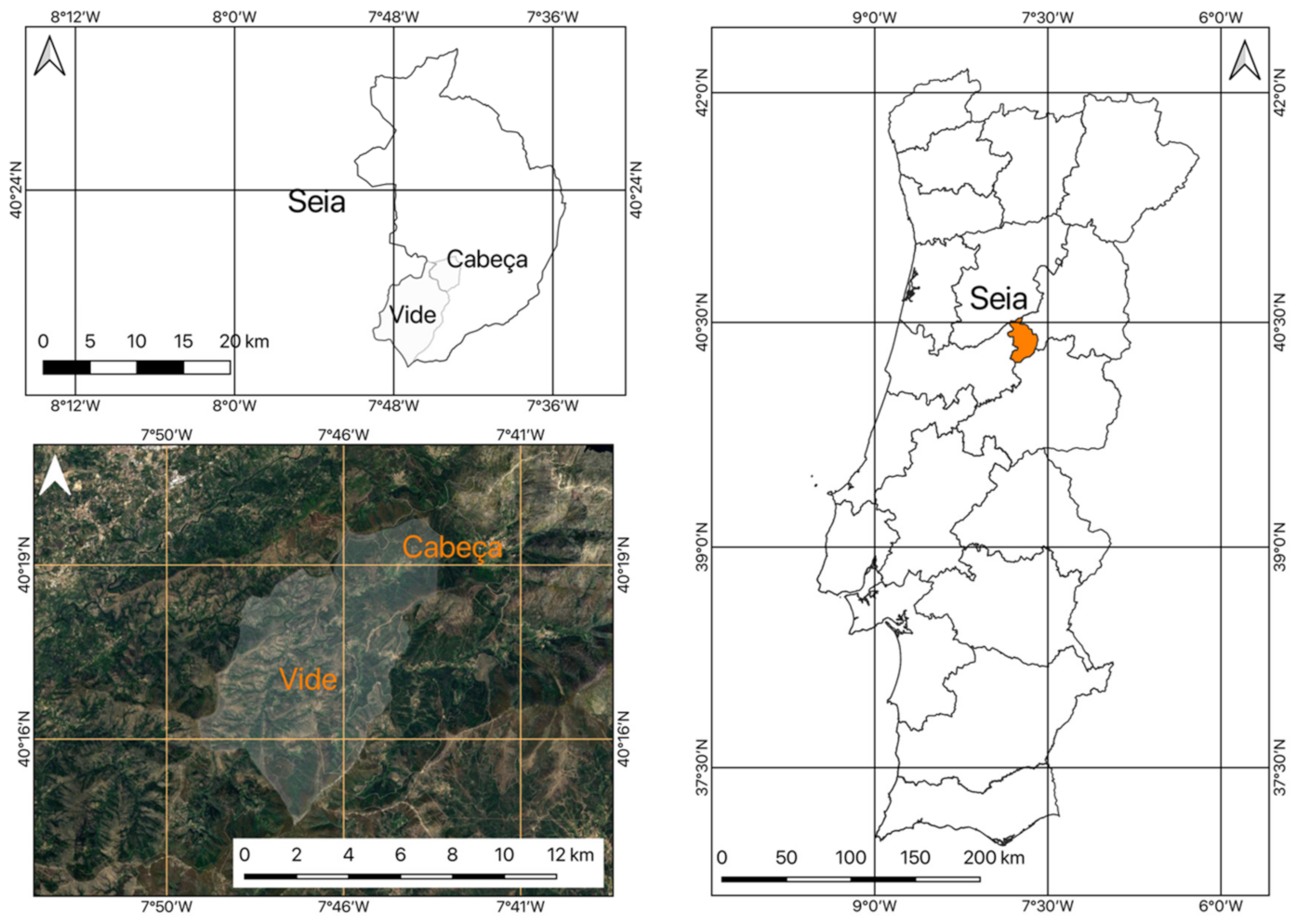
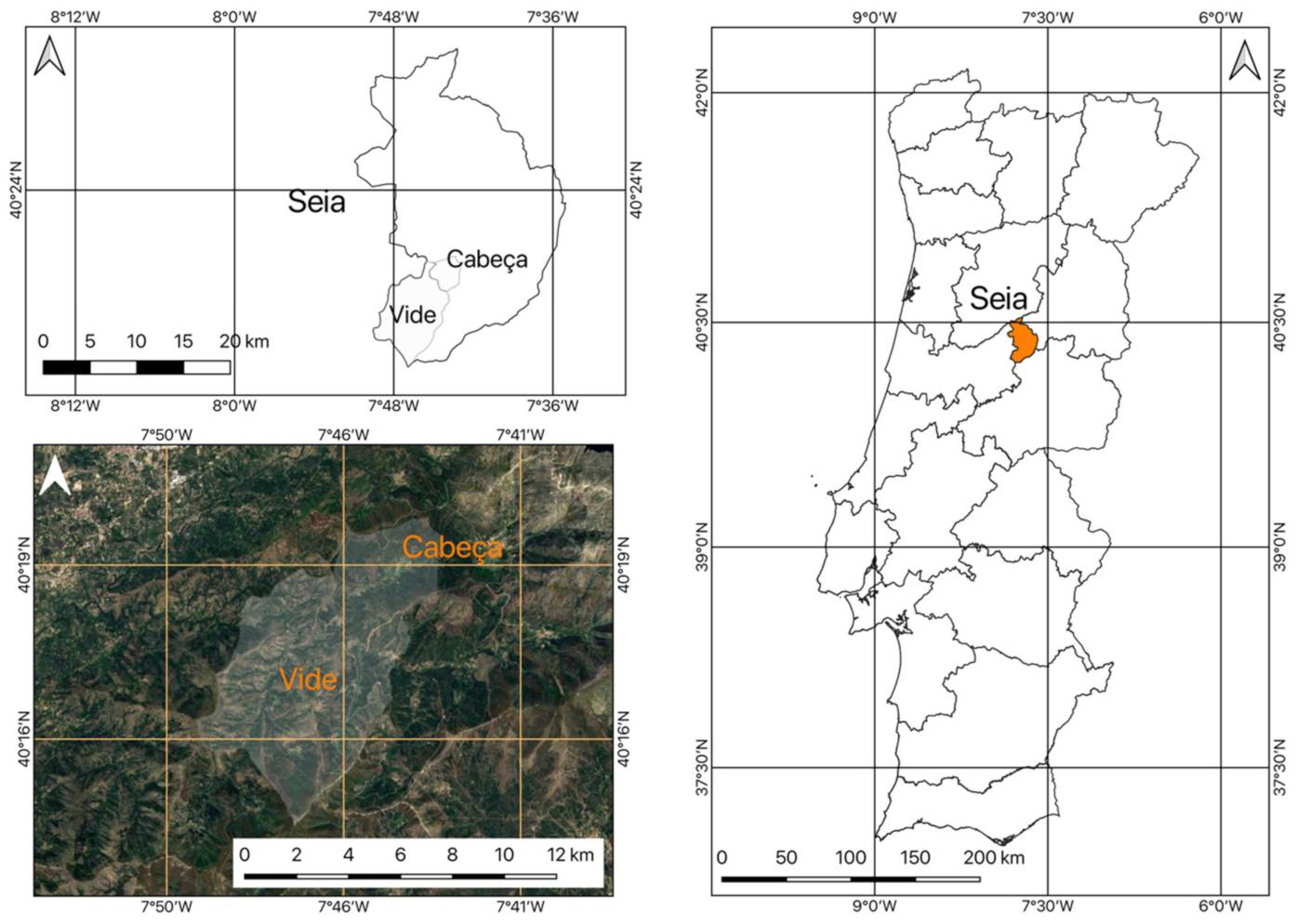
2.1. Location of the Study Area
2.2. Framework of the Area under Study
2.2.1. Geology and Geomorphology
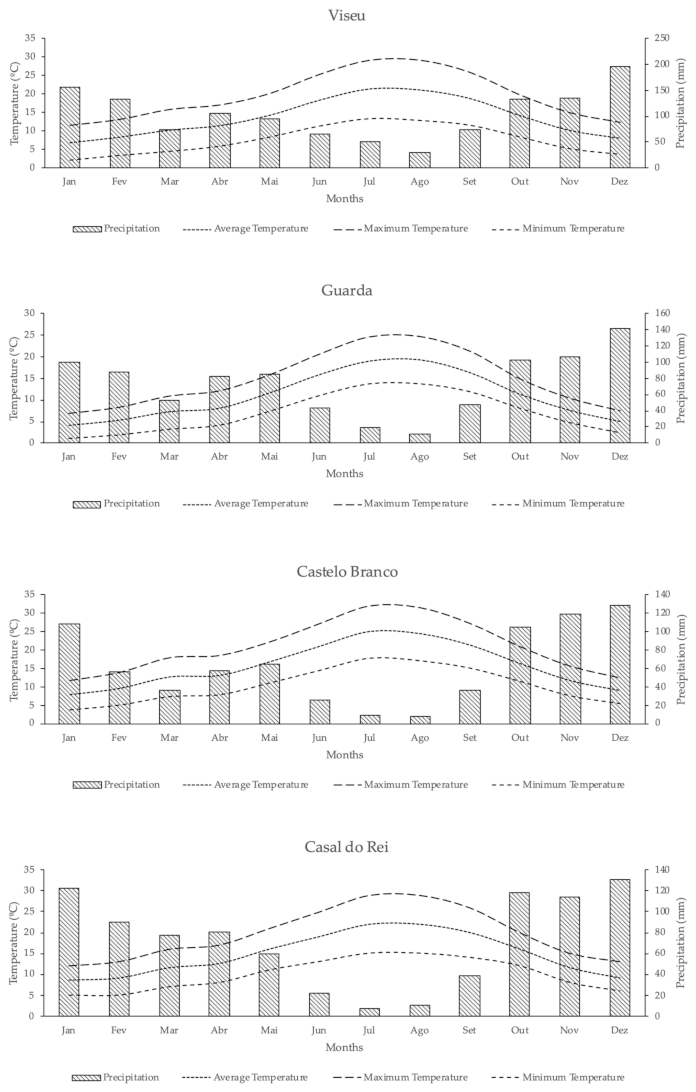
2.2.2. Climatology
2.3. Study Species
2.4. Data Acquisition
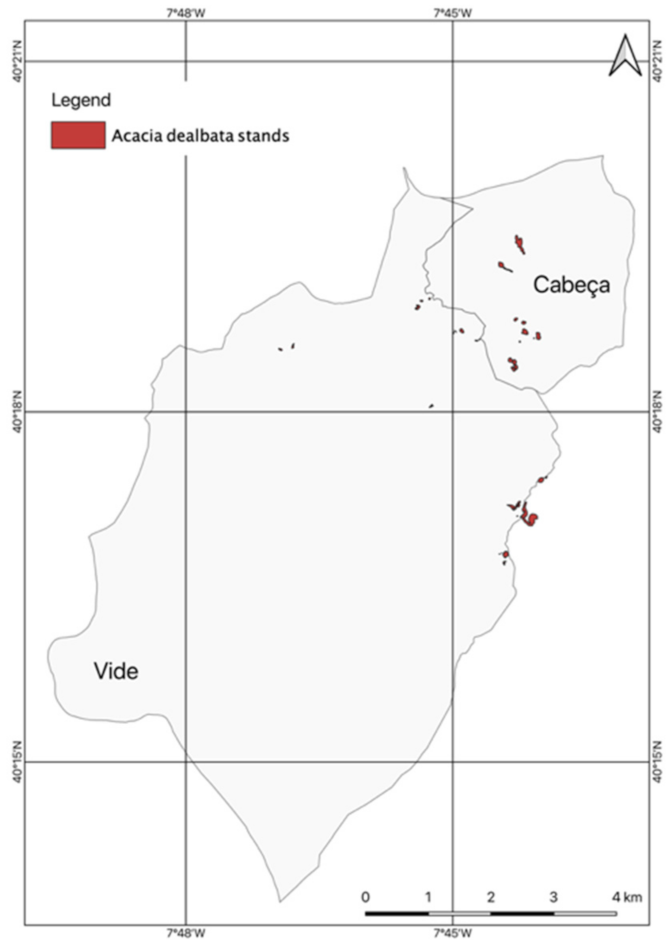
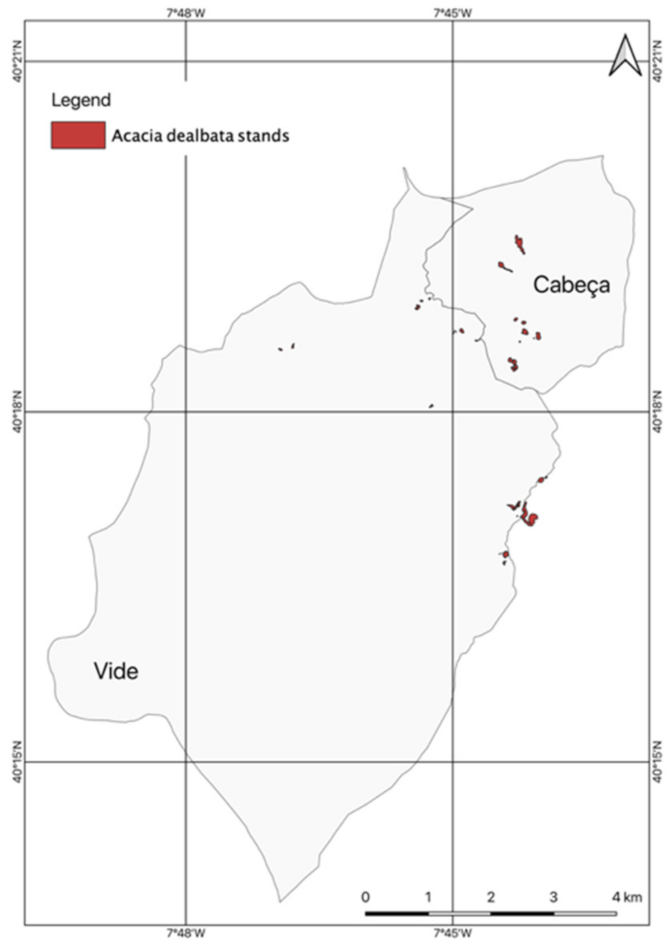
2.4.1. Occurrences of A. dealbata Stands in the União de Freguesia de Cabeça e Vide
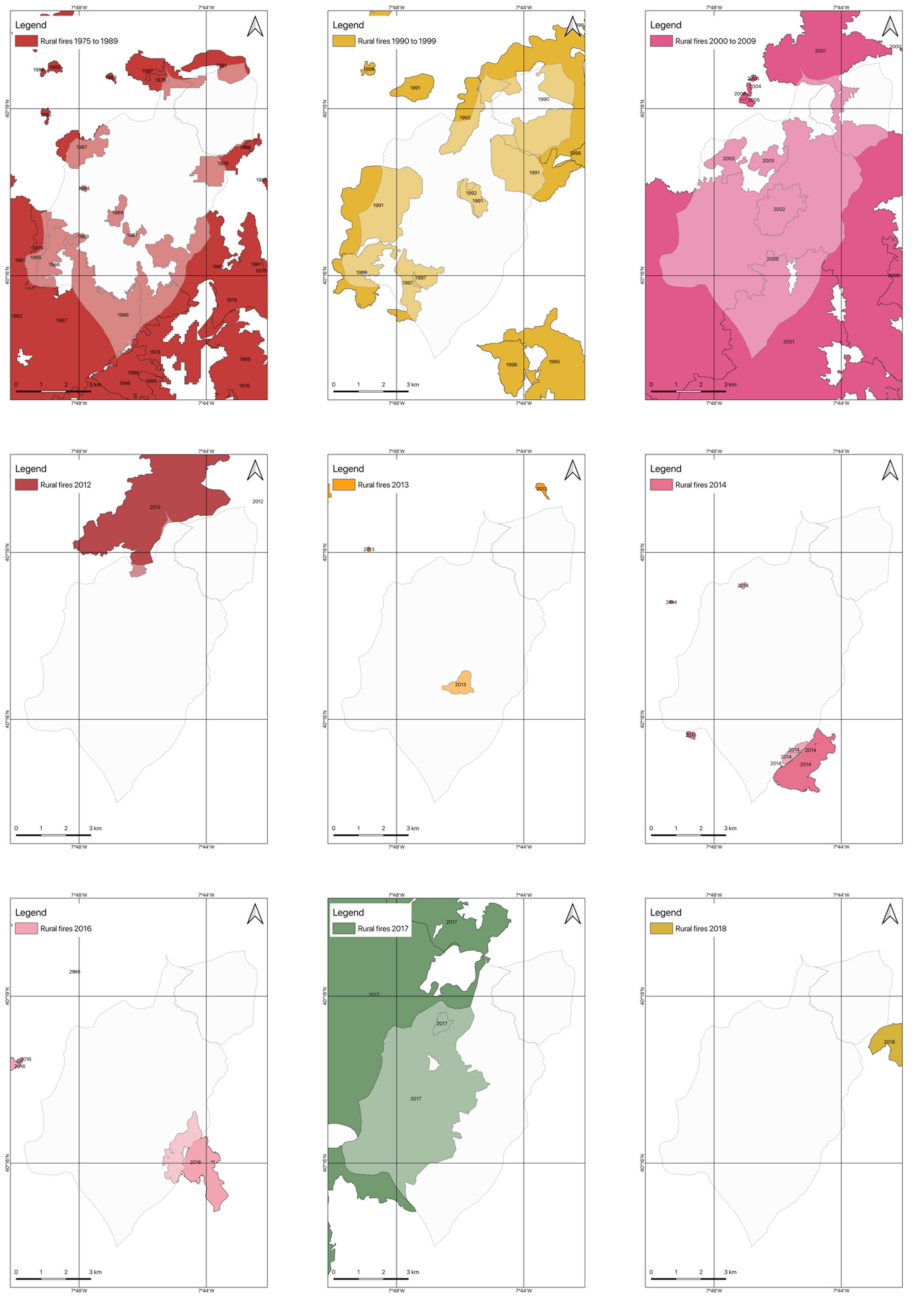
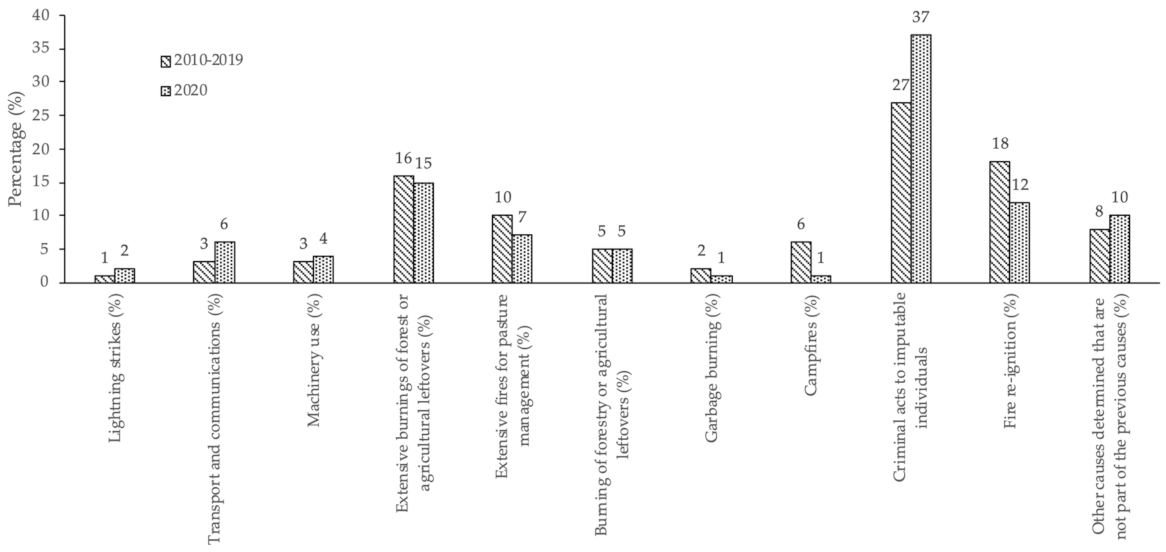
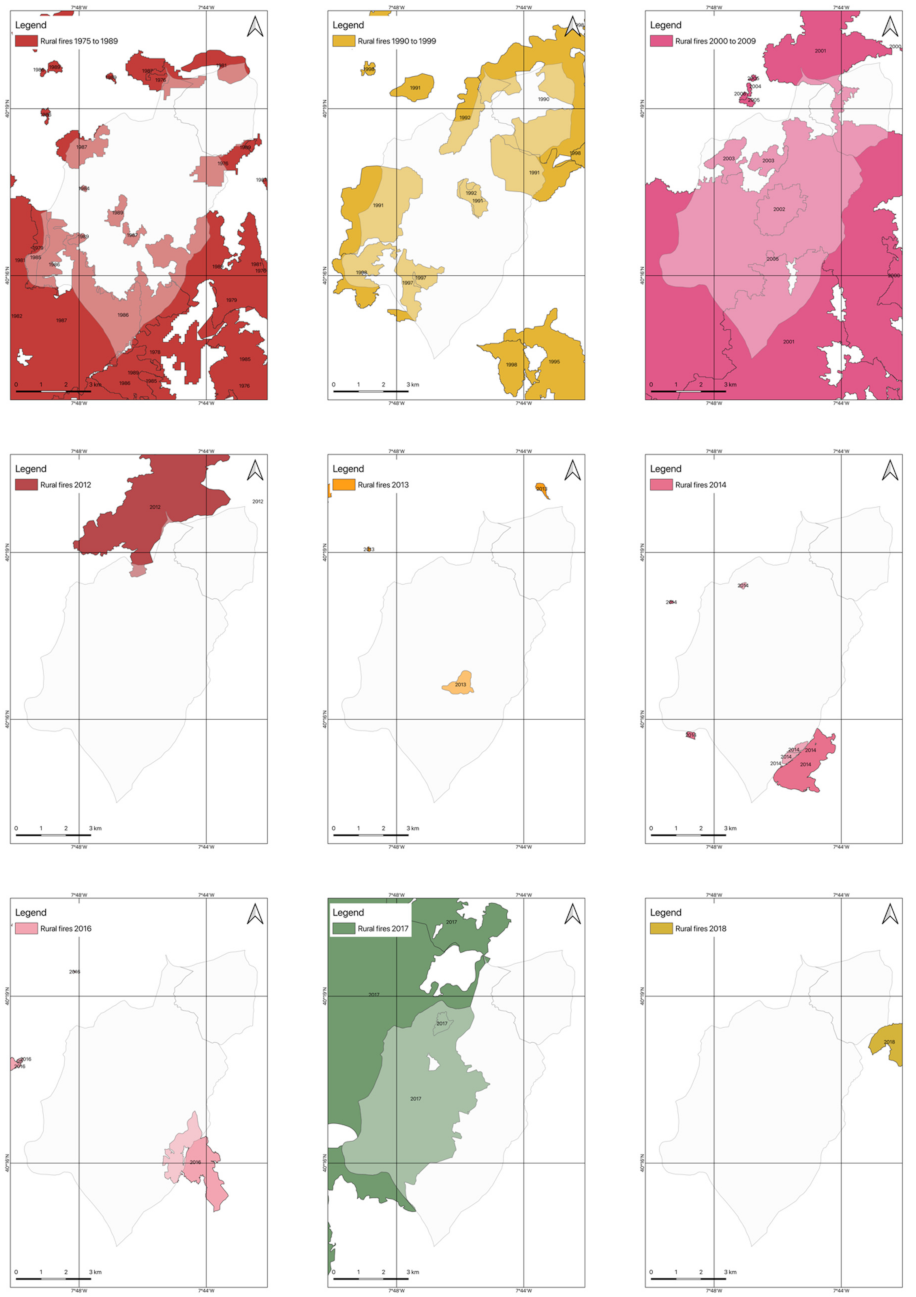
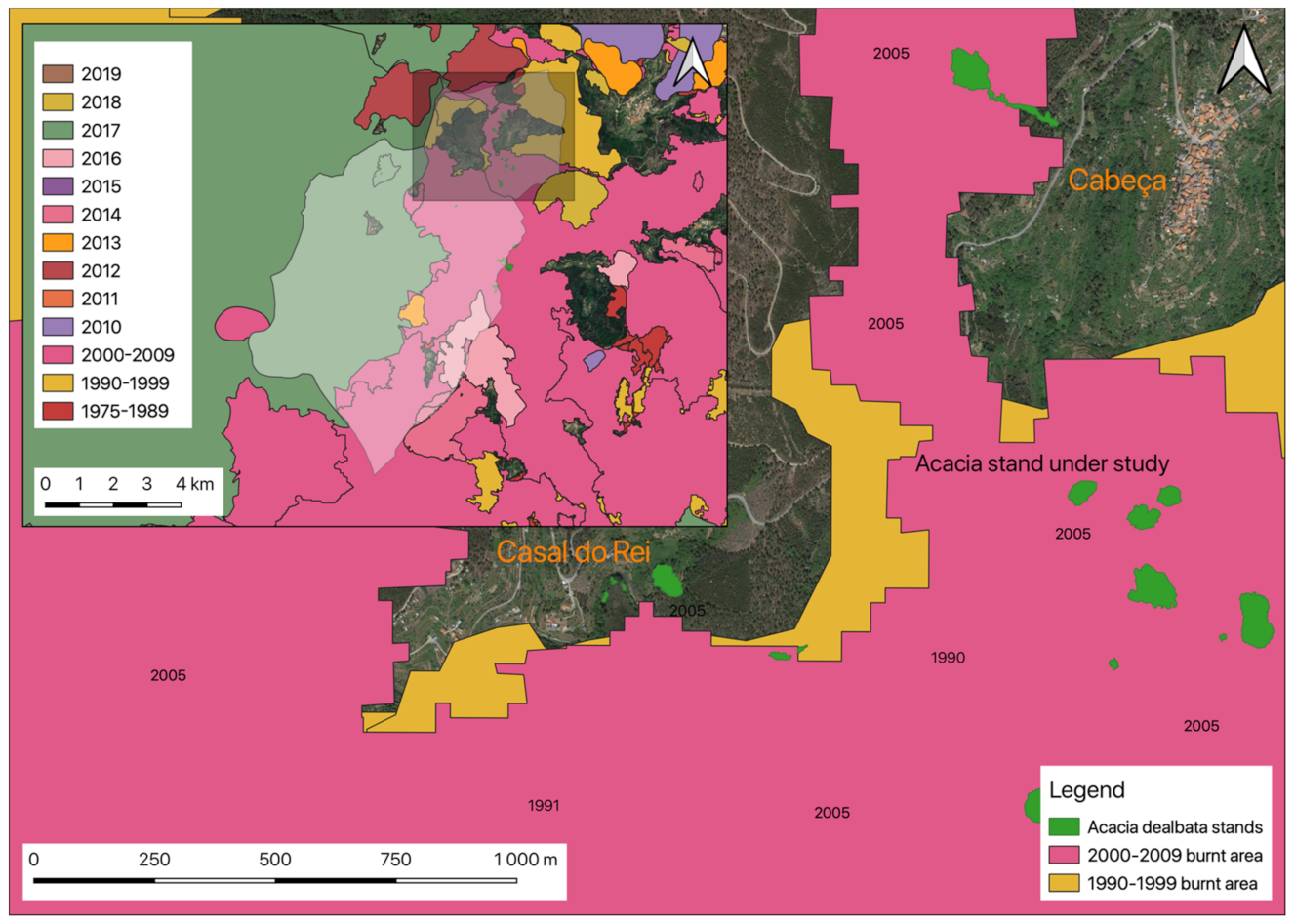
2.4.2. Rural Fires Occurrences
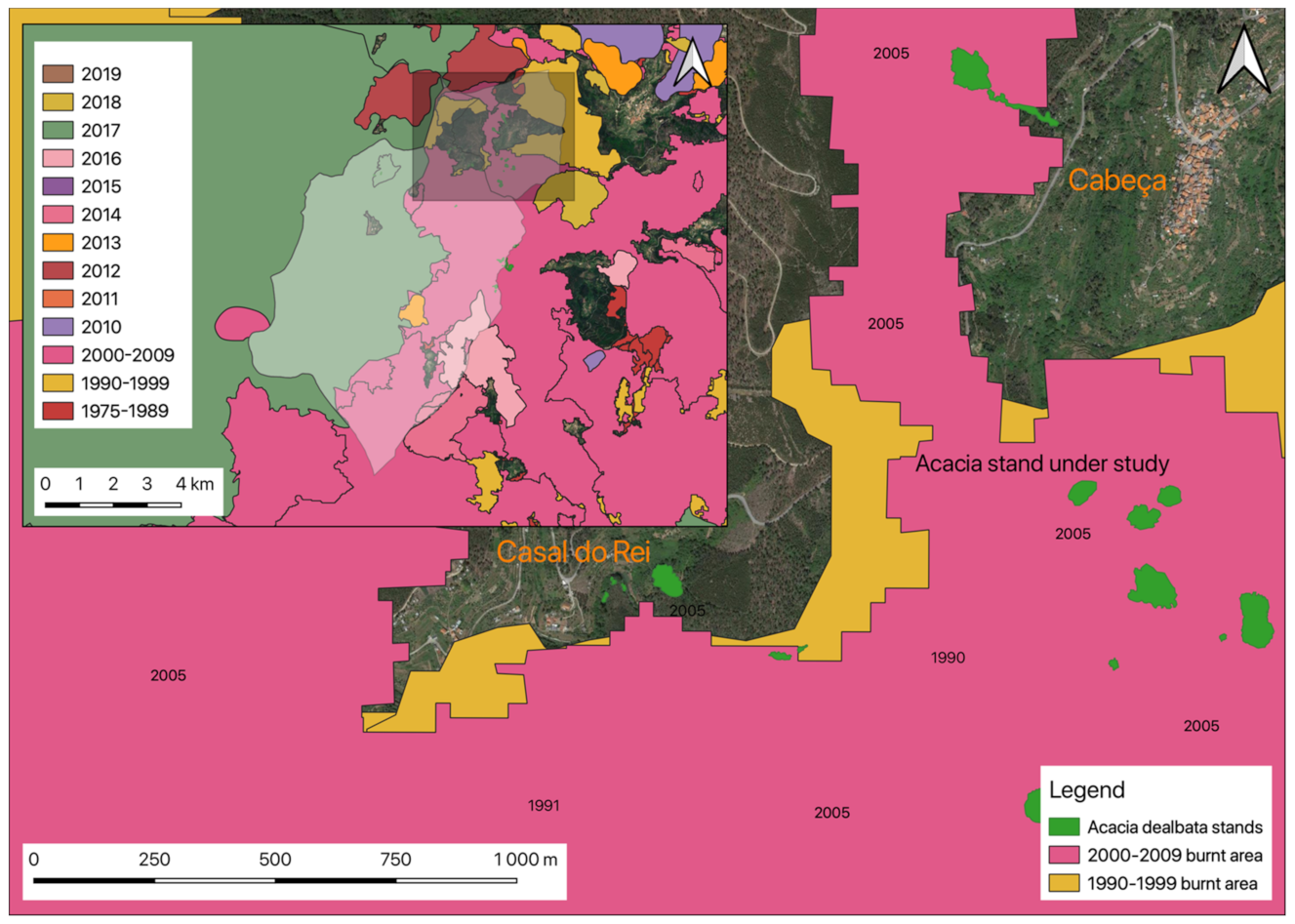
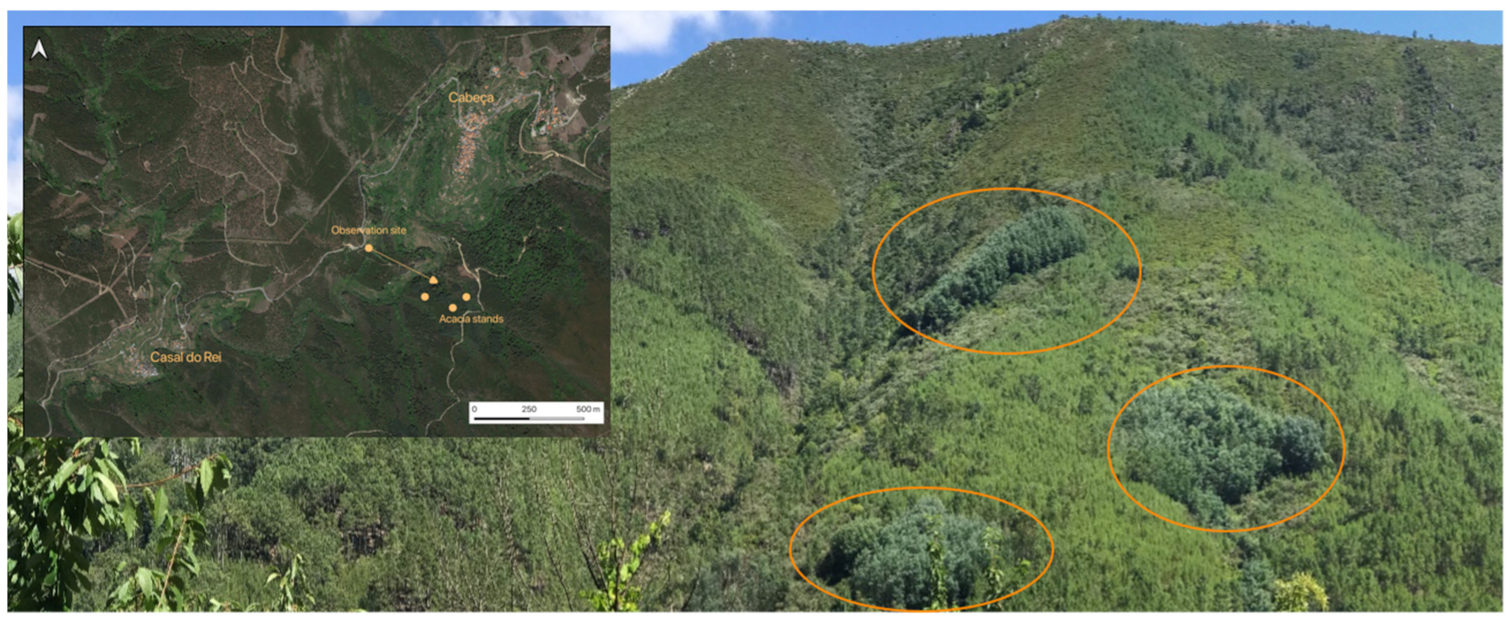
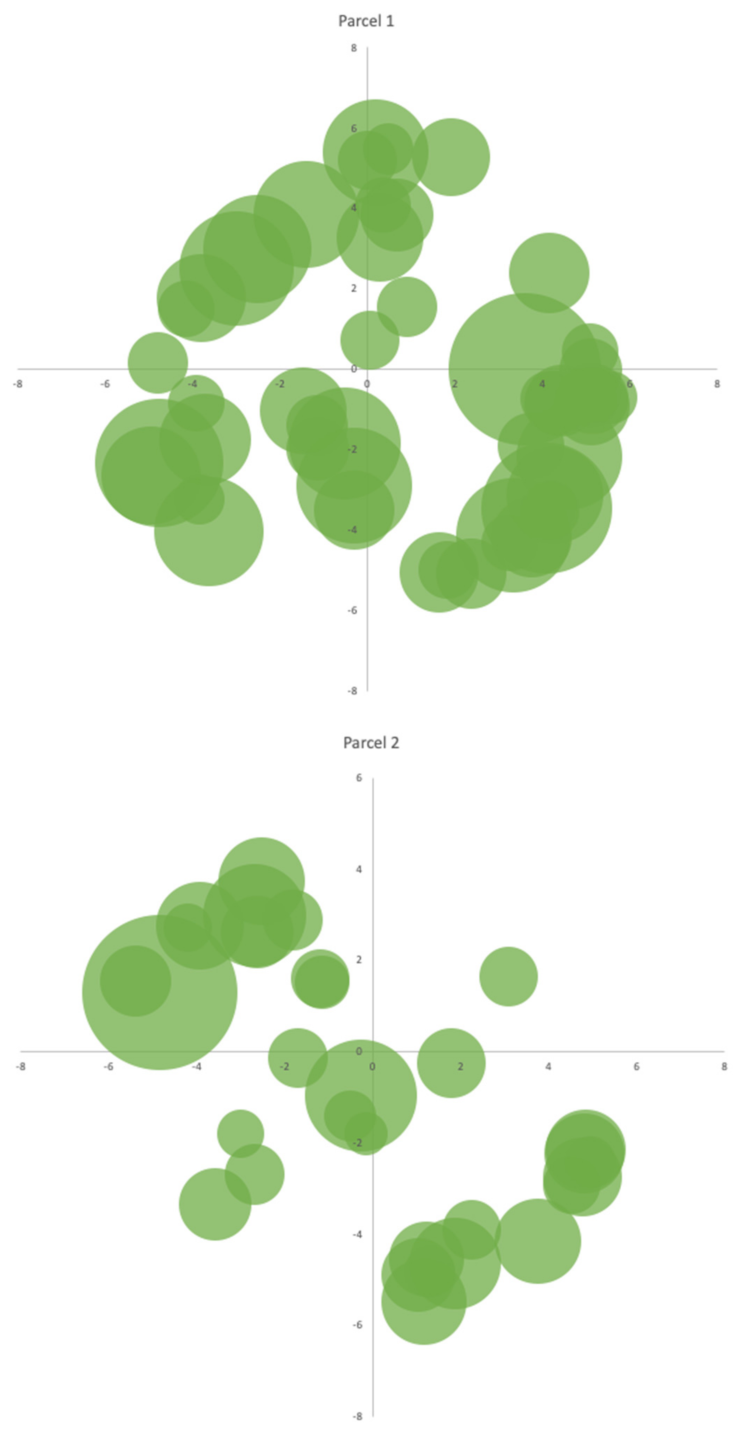
2.4.3. A. dealbata Stand Selection
- Class 5—DBH less than 5 cm;
- Class 10—DBH between 5 and 10 cm;
- Class 15—DBH between 10 and 15 cm;
- Class 20—DBH between 15 and 20 cm;
- Class 25—DBH between 20 and 25 cm;
- Class 30—DBH between 25 and 30 cm;
- Class 35—DBH between 30 and 35 cm;
- Class 40—DBH between 35 and 40 cm.
3. Results and Discussion
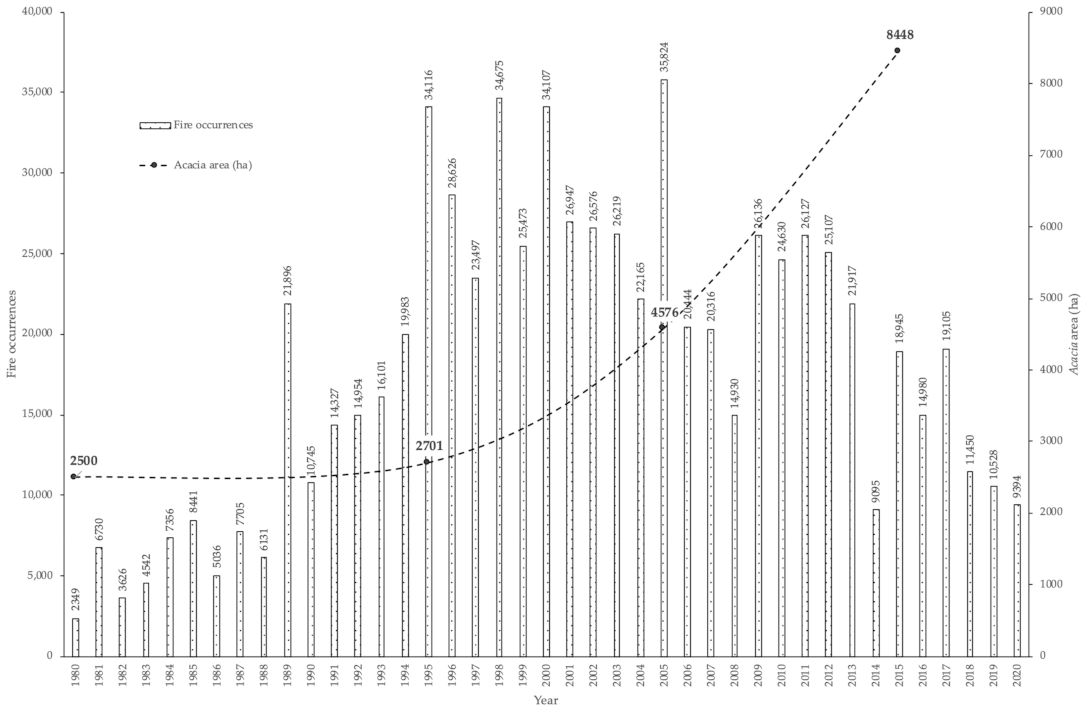
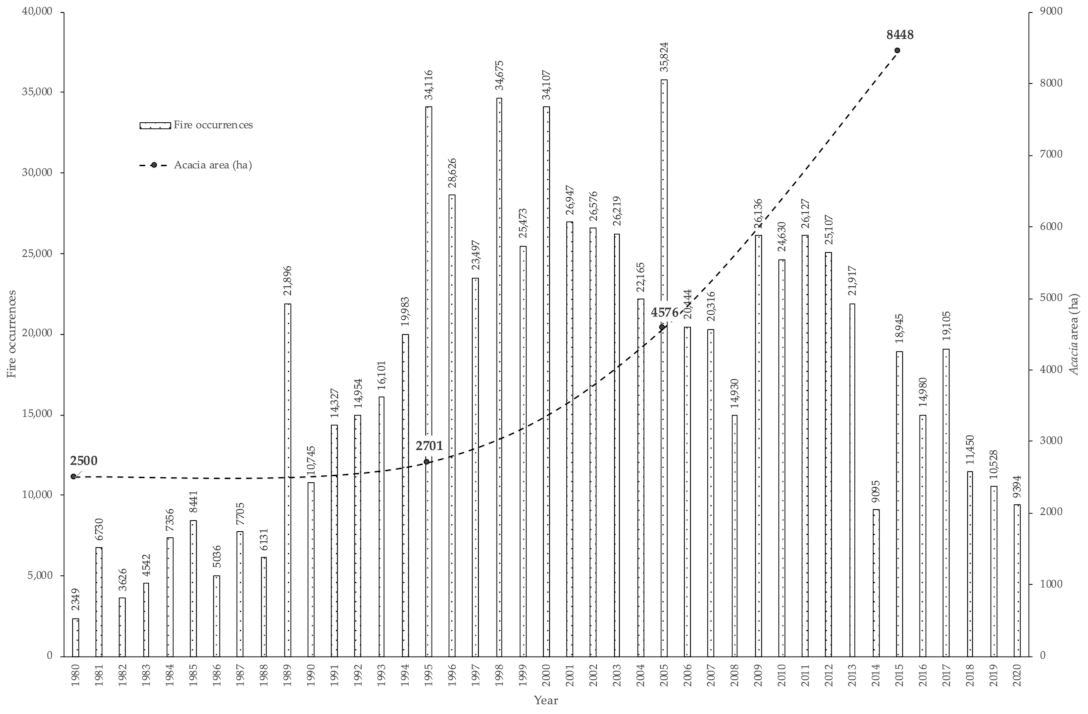
3.1. Occurrences of A. dealbata in the União de Freguesias de Vide e Cabeça
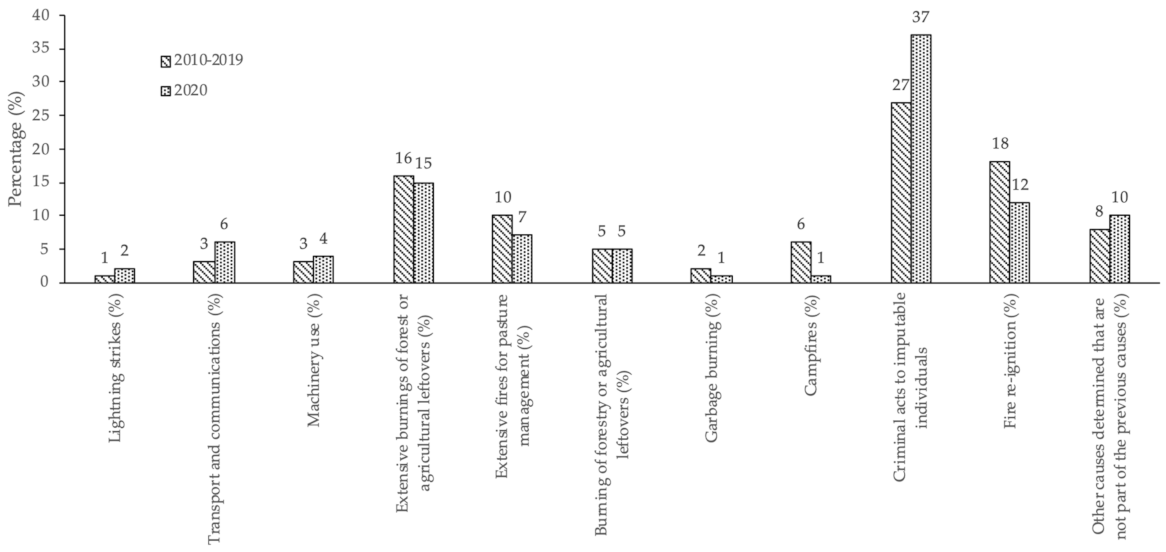
3.2. Occurrences of Rural Fires in the União de Freguesias de Vide e Cabeça
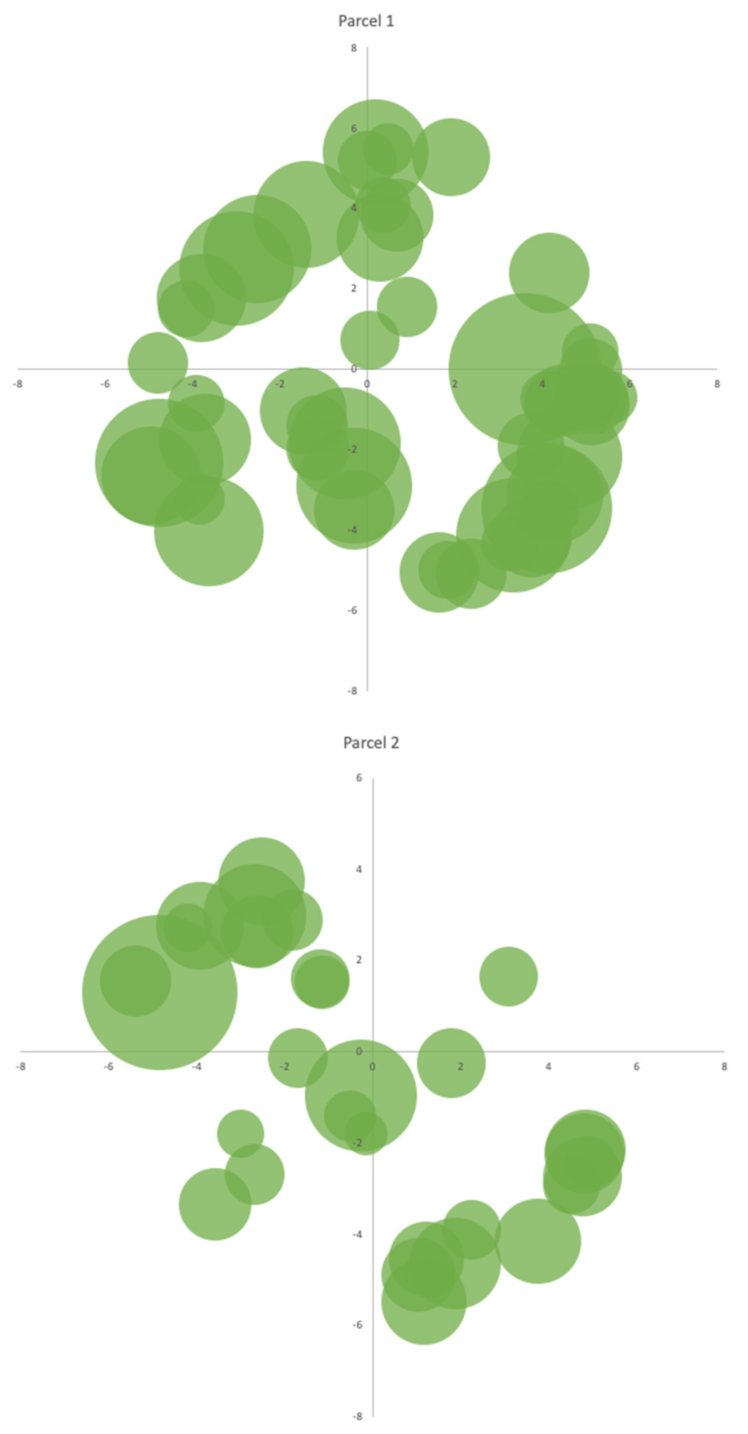
3.3. The Productivity of A. dealbata in the Casal do Rei Region
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dhillon, R.; von Wuehlisch, G. Mitigation of global warming through renewable biomass. Biomass Bioenergy 2013, 48, 75–89. [Google Scholar] [CrossRef]
- MacDonald, D.; Crabtree, J.R.; Wiesinger, G.; Dax, T.; Stamou, N.; Fleury, P.; Lazpita, J.G.; Gibon, A. Agricultural abandonment in mountain areas of Europe: Environmental consequences and policy response. J. Environ. Manag. 2000, 59, 47–69. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Filella, I.; Estiarte, M.; Llusià, J.; Ogaya, R.; Carnicer, J.; Bartrons, M.; Rivas-Ubach, A.; Grau, O. Impacts of global change on Mediterranean forests and their services. Forests 2017, 8, 463. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.; Lourenço, L.; Meira, A.C. Exploring spatial patterns and drivers of forest fires in Portugal (1980–2014). Sci. Total Environ. 2016, 573, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Estevão, C.; Costa, C.; Nunes, S.; Peraboa, F. Tourism sector competitiveness in central Portugal following the 2017 forest and rural fires: Evaluating the situation and its future implications. Rev. Tur. Desenvolv. 2020, 77–99. [Google Scholar] [CrossRef]
- Gomes, J. Forest fires in Portugal: How they happen and why they happen. Int. J. Environ. Stud. 2006, 63, 109–119. [Google Scholar] [CrossRef]
- Fernandes, M.; Devy-Vareta, N.; Rangan, H. Plantas exóticas invasoras e instrumentos de gestão territorial. O caso paradigmático do género Acacia em Portugal. Rev. Geogr. Ordenam. Territ. 2013, 1, 83–107. [Google Scholar] [CrossRef] [Green Version]
- Nunes, L.J.; Meireles, C.I.; Pinto Gomes, C.J.; Almeida Ribeiro, N. Historical development of the portuguese forest: The introduction of invasive species. Forests 2019, 10, 974. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, S.; Monteiro, E.; Brito, P.; Vilarinho, C. Biomass resources in Portugal: Current status and prospects. Renew. Sustain. Energy Rev. 2017, 78, 1221–1235. [Google Scholar] [CrossRef]
- Nunes, L.J.; Raposo, M.A.; Meireles, C.I.; Pinto Gomes, C.J.; Ribeiro, N.; Almeida, M. Control of Invasive Forest Species through the Creation of a Value Chain: Acacia dealbata Biomass Recovery. Environments 2020, 7, 39. [Google Scholar] [CrossRef]
- Minuto, L.; Casazza, G.; Dagnino, D.; Guerrina, M.; Macrì, C.; Zappa, E.; Mariotti, M.G. REPRODUCTIVE TRAITS OF THE INVASIVE SPECIES ACACIA DEALBATA LINK. IN THE NORTHERN MEDITERRANEAN BASIN. Ann. Bot. 2020, 10, 13–20. [Google Scholar]
- Martins, F.; Alegria, C.; Gil, A. Mapping invasive alien Acacia dealbata Link using ASTER multispectral imagery: A case study in central-eastern of Portugal. For. Syst. 2016, 25, 13. [Google Scholar] [CrossRef] [Green Version]
- Correia, M.; Montesinos, D.; French, K.; Rodríguez-Echeverría, S. Evidence for enemy release and increased seed production and size for two invasive Australian acacias. J. Ecol. 2016, 104, 1391–1399. [Google Scholar] [CrossRef]
- Lorenzo, P.; González, L.; Reigosa, M.J. The genus Acacia as invader: The characteristic case of Acacia dealbata Link in Europe. Ann. For. Sci. 2010, 67, 101. [Google Scholar] [CrossRef] [Green Version]
- DiTomaso, J.M.; Johnson, D.W. The use of fire as a tool for controlling invasive plants. Cal-IPC Publ. 2006, 1, 56. [Google Scholar]
- Lorenzo, P.; Pazos-Malvido, E.; Reigosa, M.J.; González, L. Differential responses to allelopathic compounds released by the invasive Acacia dealbata Link (Mimosaceae) indicate stimulation of its own seed. Aust. J. Bot. 2010, 58, 546–553. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, N.; Becerra, J.; Villaseñor-Parada, C.; Lorenzo, P.; González, L.; Hernández, V. Effects and identification of chemical compounds released from the invasive Acacia dealbata Link. Chem. Ecol. 2015, 31, 479–493. [Google Scholar] [CrossRef]
- Le Maitre, D.C.; Gaertner, M.; Marchante, E.; Ens, E.J.; Holmes, P.M.; Pauchard, A.; O’Farrell, P.J.; Rogers, A.M.; Blanchard, R.; Blignaut, J. Impacts of invasive Australian acacias: Implications for management and restoration. Divers. Distrib. 2011, 17, 1015–1029. [Google Scholar] [CrossRef]
- Gibson, M.R.; Richardson, D.M.; Marchante, E.; Marchante, H.; Rodger, J.G.; Stone, G.N.; Byrne, M.; Fuentes-Ramírez, A.; George, N.; Harris, C. Reproductive biology of Australian acacias: Important mediator of invasiveness? Divers. Distrib. 2011, 17, 911–933. [Google Scholar] [CrossRef]
- Orchard, A.E.; Maslin, B.R. (1584) Proposal to conserve the name Acacia (Leguminosae: Mimosoideae) with a conserved type. Taxon 2003, 52, 362–363. [Google Scholar] [CrossRef]
- Bouchenak-Khelladi, Y.; Maurin, O.; Hurter, J.; Van der Bank, M. The evolutionary history and biogeography of Mimosoideae (Leguminosae): An emphasis on African acacias. Mol. Phylogenet. Evol. 2010, 57, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Andrade, P. Das mimosas e outras acácias. In Histórias da Vida e da Terra—Um blog de História Natural; Wordpress.com: San Francisco, CA, USA, 2012; Volume 2019. [Google Scholar]
- Aguiar, F.; Ferreira, M. Plant invasions in the rivers of the Iberian Peninsula, south-western Europe: A review. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2013, 147, 1107–1119. [Google Scholar] [CrossRef]
- Fernandes, P.M. Combining forest structure data and fuel modelling to classify fire hazard in Portugal. Ann. For. Sci. 2009, 66, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, M.M. Acácias e geografia histórica: Rotas de um percurso global (parte1). Cad. Curso Doutor. Geogr. 2012, 4, 23–40. [Google Scholar]
- Fernandes, M. Recuperação Ecológica de Áreas Invadidas por Acacia Dealbata Link no Vale do rio Gerês: Um Trabalho de Sísifo. Master’s Thesis, Universidade de Trás-os-Montes e Alto-Douro, Vila Real, Portugal, 2008. [Google Scholar]
- Simões, C.A.d.C.P. A Degradação da Paisagem e a Sua Perceção Após Invasão pela Espécie Acacia Dealbata Link.: O Caso da Região do Alto Ceira. Master’s Thesis, Universidade Nova de Lisboa, Lisbon, Portugal, 2016. [Google Scholar]
- González, L.; Souto, X.C.; Reigosa, M. Allelopathic effects of Acacia melanoxylon R. Br. phyllodes during their decomposition. For. Ecol. Manag. 1995, 77, 53–63. [Google Scholar] [CrossRef]
- Ramoliya, P.; Patel, H.; Pandey, A. Effect of salinisation of soil on growth and macro-and micro-nutrient accumulation in seedlings of Acacia catechu (Mimosaceae). Ann. Appl. Biol. 2004, 144, 321–332. [Google Scholar] [CrossRef]
- Martins-Corder, M.P.; Borges, R.Z.; Junior, N.B. Fotoperiodismo e quebra de dormência em sementes de acácia-negra (Acacia mearnsii De Wild.). Ciência Florest. 1999, 9, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, L.M.; Antunes, P.M.; Martins-Loução, M.A.; Klironomos, J.N. Disturbance influences the outcome of plant–soil biota interactions in the invasive Acacia longifolia and in native species. Oikos 2010, 119, 1172–1180. [Google Scholar] [CrossRef]
- Coelho, S.I.D.B.F. Factores Facilitadores da Invasibilidade de Acacia Dealbata em Função do Uso do Solo. Master’s Thesis, Universidade de Lisboa, Lisboa, Portugal, 2014. [Google Scholar]
- González-Muñoz, N.; Costa-Tenorio, M.; Espigares, T. Invasion of alien Acacia dealbata on Spanish Quercus robur forests: Impact on soils and vegetation. For. Ecol. Manag. 2012, 269, 214–221. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 2003, 6, 503–523. [Google Scholar] [CrossRef]
- Lorenzo, P.; Pereira, C.S.; Rodríguez-Echeverría, S. Differential impact on soil microbes of allelopathic compounds released by the invasive Acacia dealbata Link. Soil Biol. Biochem. 2013, 57, 156–163. [Google Scholar] [CrossRef]
- Wilson, J.R.; Gairifo, C.; Gibson, M.R.; Arianoutsou, M.; Bakar, B.B.; Baret, S.; Celesti-Grapow, L.; DiTomaso, J.M.; Dufour-Dror, J.M.; Kueffer, C. Risk assessment, eradication, and biological control: Global efforts to limit Australian acacia invasions. Divers. Distrib. 2011, 17, 1030–1046. [Google Scholar] [CrossRef] [Green Version]
- Albaugh, T.J.; Rubilar, R.A.; Maier, C.A.; Acuna, E.A.; Cook, R.L. Biomass and nutrient mass of Acacia dealbata and Eucalyptus globulus bioenergy plantations. Biomass Bioenergy 2017, 97, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Gouws, A.J.; Shackleton, C.M. Abundance and correlates of the Acacia dealbata invasion in the northern Eastern Cape, South Africa. For. Ecol. Manag. 2019, 432, 455–466. [Google Scholar] [CrossRef]
- Hunt, M.A.; Beadle, C.L. Whole-tree transpiration and water-use partitioning between Eucalyptus nitens and Acacia dealbata weeds in a short-rotation plantation in northeastern Tasmania. Tree Physiol. 1998, 18, 557–563. [Google Scholar] [CrossRef]
- Forrester, D.I.; Theiveyanathan, S.; Collopy, J.J.; Marcar, N.E. Enhanced water use efficiency in a mixed Eucalyptus globulus and Acacia mearnsii plantation. For. Ecol. Manag. 2010, 259, 1761–1770. [Google Scholar] [CrossRef]
- Vertessy, R.A.; Watson, F.G.; Sharon, K. Factors determining relations between stand age and catchment water balance in mountain ash forests. For. Ecol. Manag. 2001, 143, 13–26. [Google Scholar] [CrossRef]
- Bowd, E.J.; Banks, S.C.; Strong, C.L.; Lindenmayer, D.B. Long-term impacts of wildfire and logging on forest soils. Nat. Geosci. 2019, 12, 113–118. [Google Scholar] [CrossRef]
- Je, K. Fire management impacts on invasive plant species in the western United States. Conserv. Biol. 2006, 20, 375–384. [Google Scholar]
- Florentine, S.; Milberg, P.; Gibson, M.; Westbrooke, M. Post-wildfire seedling colonisation patterns in a Eucalyptus delegatensis (Myrtaceae) windthrow site at Snowy River National Park, Victoria. Aust. For. 2008, 71, 48–53. [Google Scholar] [CrossRef]
- Gordon, C.E.; Price, O.F.; Tasker, E.M.; Denham, A.J. Acacia shrubs respond positively to high severity wildfire: Implications for conservation and fuel hazard management. Sci. Total Environ. 2017, 575, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.L.; Torres, R.C.; Renison, D. Do wildfires promote woody species invasion in a fire-adapted ecosystem? Post-fire resprouting of native and non-native woody plants in central Argentina. Environ. Manag. 2016, 57, 308–317. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parcel | Sample | DBH Class | Height (m) | Canopy Diameter (m) | Growth Start Year | Age (Years) | Total Weight (kg) | Annual Biomass Increment (kg.year−1) |
|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 15 | 23.6 | 1 | 2008 | 10 | 54.6 | 5.5 |
| 5 | 10 | 15.4 | 1 | 2013 | 5 | 28.8 | 5.8 | |
| 6 | 10 | 13.2 | 2.1 | 2004 | 14 | 33.5 | 2.4 | |
| 8 | 30 | 22.9 | 9.5 | 1998 | 20 | 594.8 | 29.7 | |
| 16 | 10 | 18.5 | 2 | 2008 | 10 | 47.8 | 4.8 | |
| 17 | 10 | 11.3 | 2.5 | 2009 | 9 | 18.5 | 2.1 | |
| 19 | 15 | 17.9 | 4.3 | 2006 | 12 | 166.8 | 13.9 | |
| 26 | 20 | 25.1 | 1.2 | 2004 | 14 | 249.1 | 17.8 | |
| 29 | 5 | 9.9 | 1.8 | 2012 | 6 | 4.1 | 0.7 | |
| 30 | 10 | 23.7 | 1 | 2009 | 9 | 45.5 | 5.1 | |
| 33 | 15 | 23.4 | 3.8 | 2004 | 14 | 223.9 | 16.0 | |
| 2 | 9 | 15 | 19.9 | 3.4 | 2003 | 15 | 138.9 | 9.3 |
| 10 | 10 | 15.7 | 1.9 | 2008 | 10 | 40.7 | 4.1 | |
| 11 | 5 | 12.5 | 1.9 | 2009 | 9 | 13.6 | 1.5 | |
| 15 | 25 | 32.7 | 2.65 | 2001 | 17 | 333.4 | 19.6 | |
| 21 | 20 | 10.3 | 3.2 | 2006 | 12 | 19.7 | 1.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes, L.J.R.; Raposo, M.A.M.; Meireles, C.I.R.; Gomes, C.J.P.; Ribeiro, N.M.C.A. The Impact of Rural Fires on the Development of Invasive Species: Analysis of a Case Study with Acacia dealbata Link. in Casal do Rei (Seia, Portugal). Environments 2021, 8, 44. https://0-doi-org.brum.beds.ac.uk/10.3390/environments8050044
Nunes LJR, Raposo MAM, Meireles CIR, Gomes CJP, Ribeiro NMCA. The Impact of Rural Fires on the Development of Invasive Species: Analysis of a Case Study with Acacia dealbata Link. in Casal do Rei (Seia, Portugal). Environments. 2021; 8(5):44. https://0-doi-org.brum.beds.ac.uk/10.3390/environments8050044
Chicago/Turabian StyleNunes, Leonel J. R., Mauro A. M. Raposo, Catarina I. R. Meireles, Carlos J. Pinto Gomes, and Nuno M. C. Almeida Ribeiro. 2021. "The Impact of Rural Fires on the Development of Invasive Species: Analysis of a Case Study with Acacia dealbata Link. in Casal do Rei (Seia, Portugal)" Environments 8, no. 5: 44. https://0-doi-org.brum.beds.ac.uk/10.3390/environments8050044