
Commercial Red Food Dyes Preparations Modulate the Oxidative State in Three Model Organisms (Cucumis sativus, Artemia salina, and Danio rerio)

 , , , ,
, , , ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Food Dye Solutions
2.2. The Plant Model: Cucumis sativus L. Seedlings
2.2.1. Growth and Experimental Design
2.2.2. Photosynthetic Pigment Content, Chlorophyll Fluorescence Emission Measurements, and Dark Oxygen Consumption Determination
2.2.3. Endogenous H2O2 Determination and Plant Antioxidant Response
2.3. The Artemia salina Nauplii
2.3.1. Preparation of Nauplii
2.3.2. Oxygen Consumption, Total ROS Determination, and Susceptibility to In Vitro Oxidative Stress
2.4. The Danio rerio Embryos
2.4.1. Animal Maintenance and Treatments
2.4.2. In Vivo Routine Oxygen Consumption, Total ROS Levels, and Susceptibility to In Vitro Oxidative Stress
2.5. Statistical Analyses
3. Results
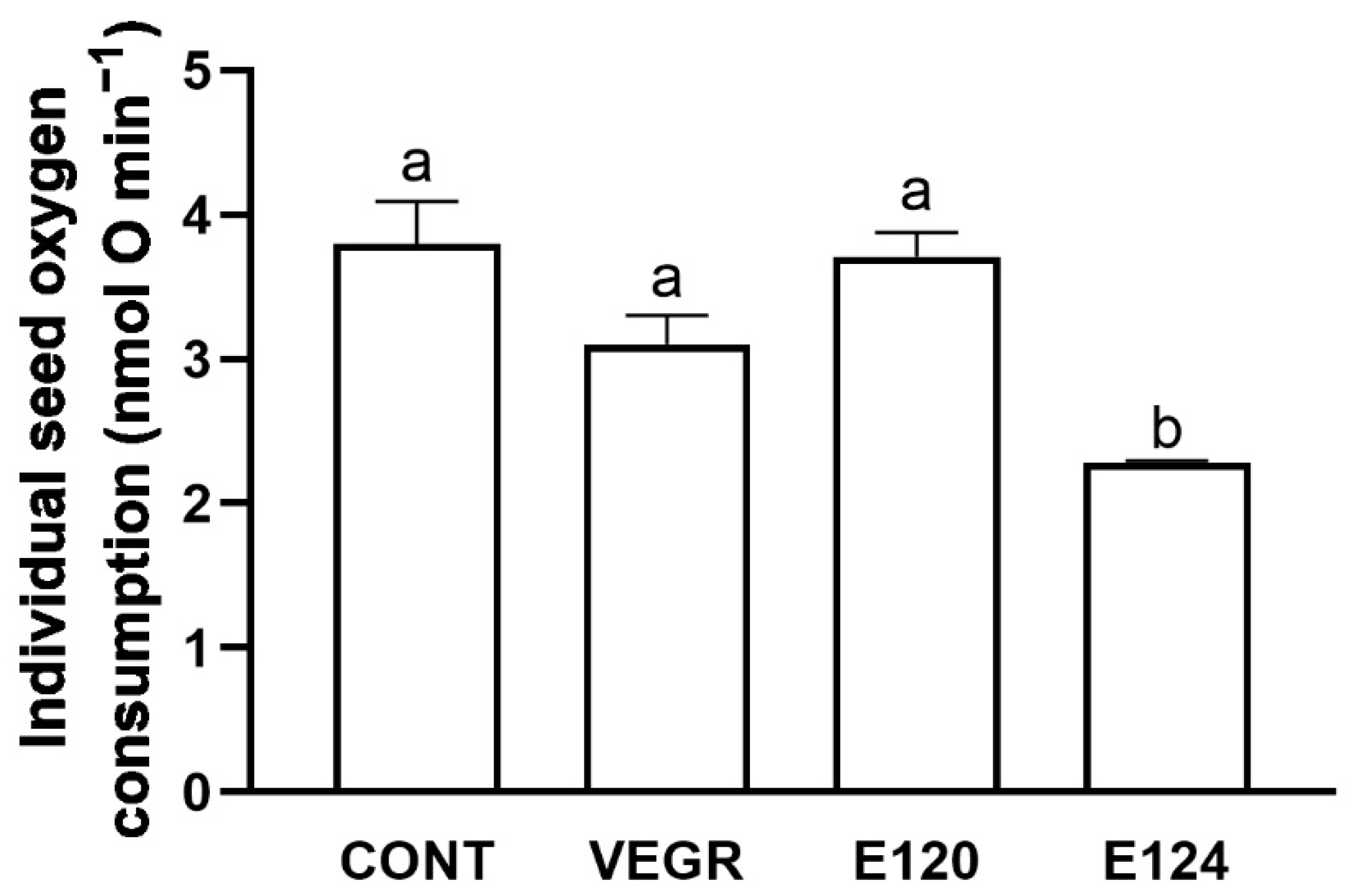
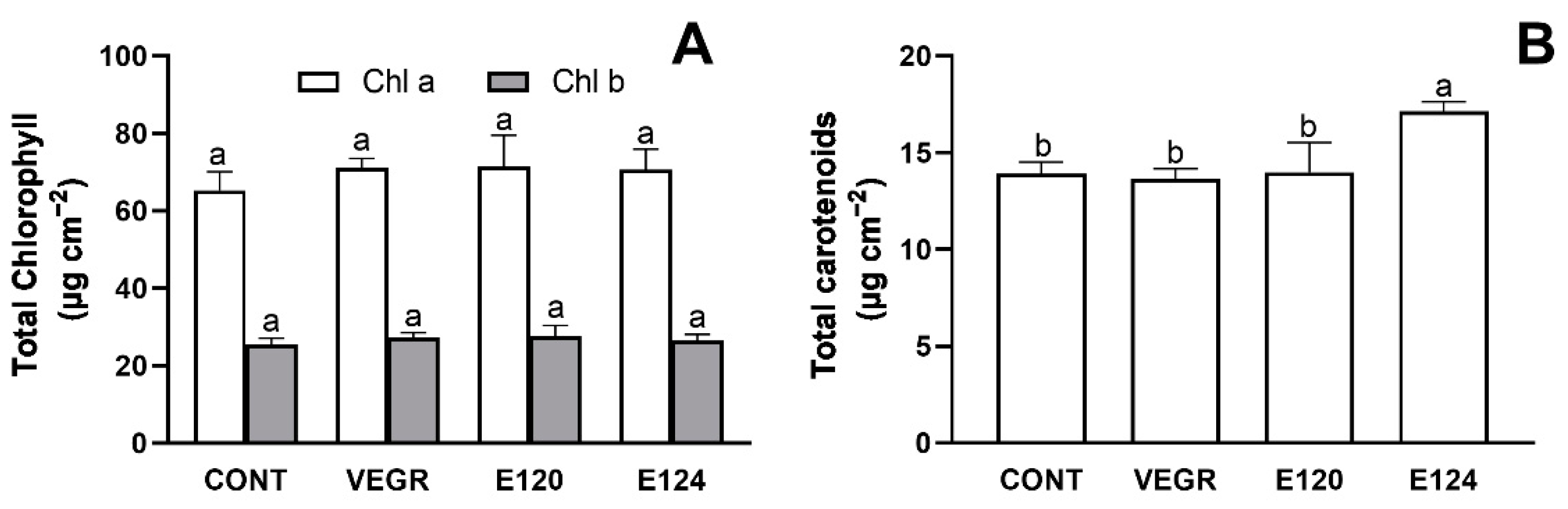
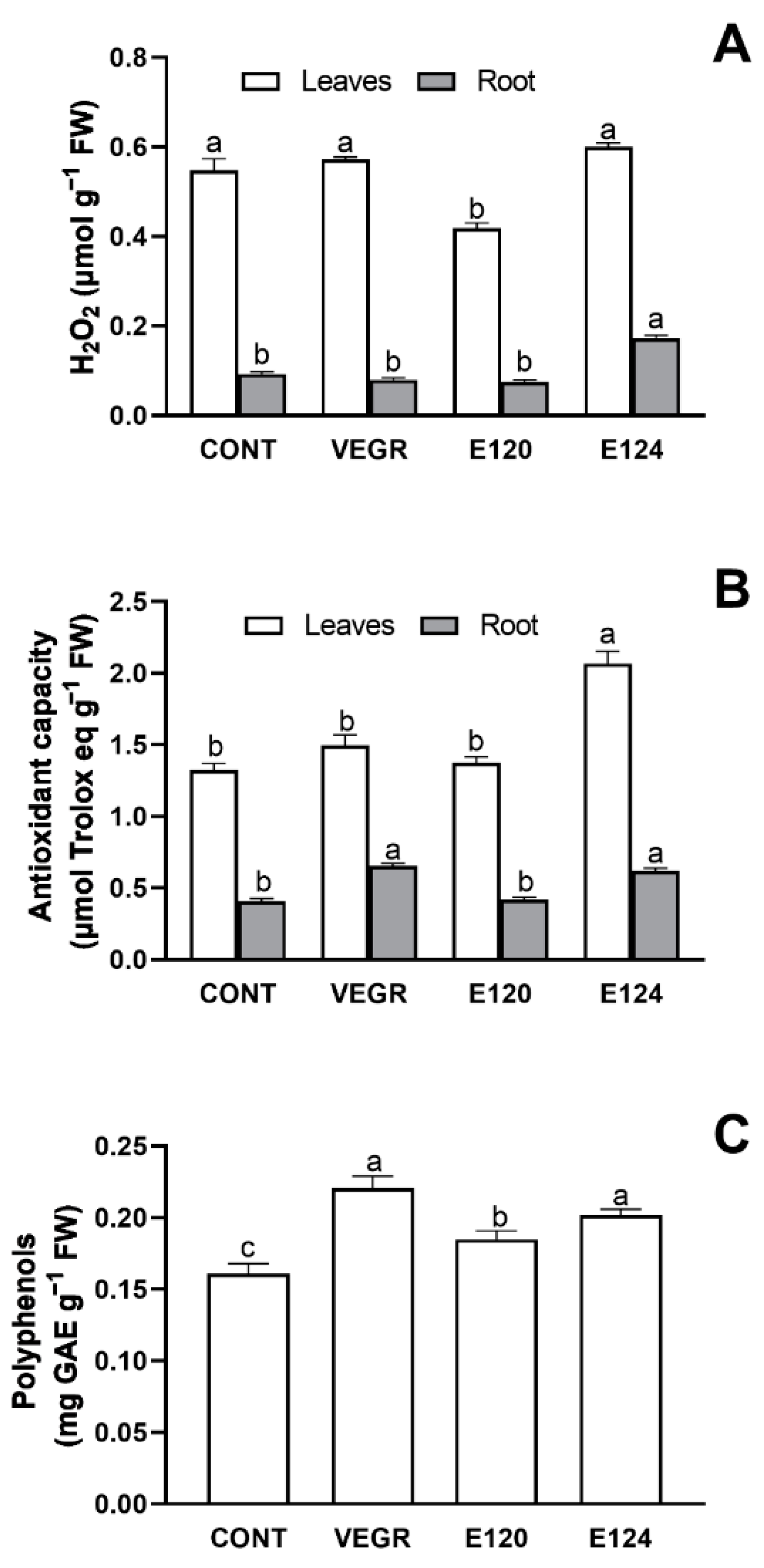
3.1. Growth and Antioxidant Response in C. sativus Seedlings Exposed to Different Red Food Dyes
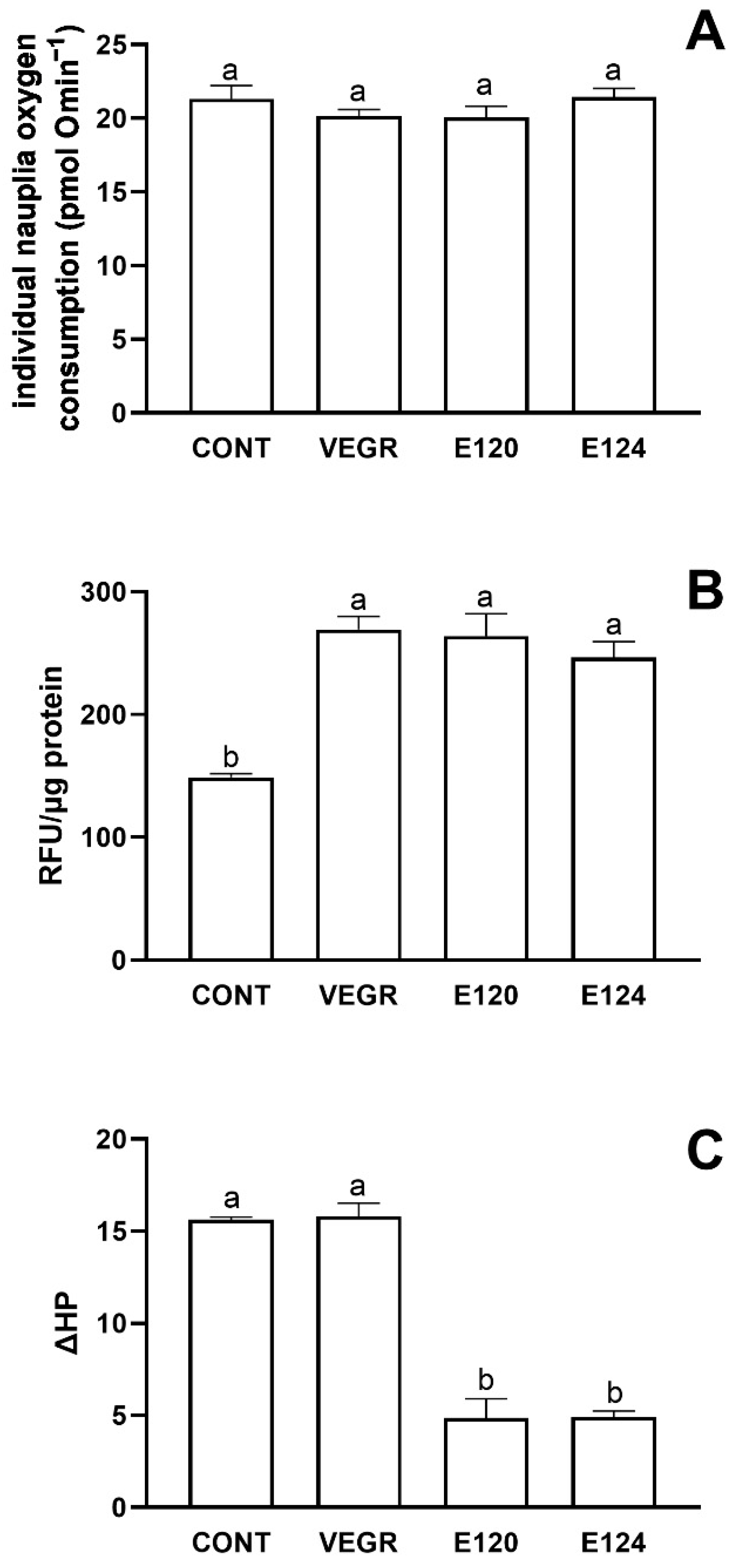
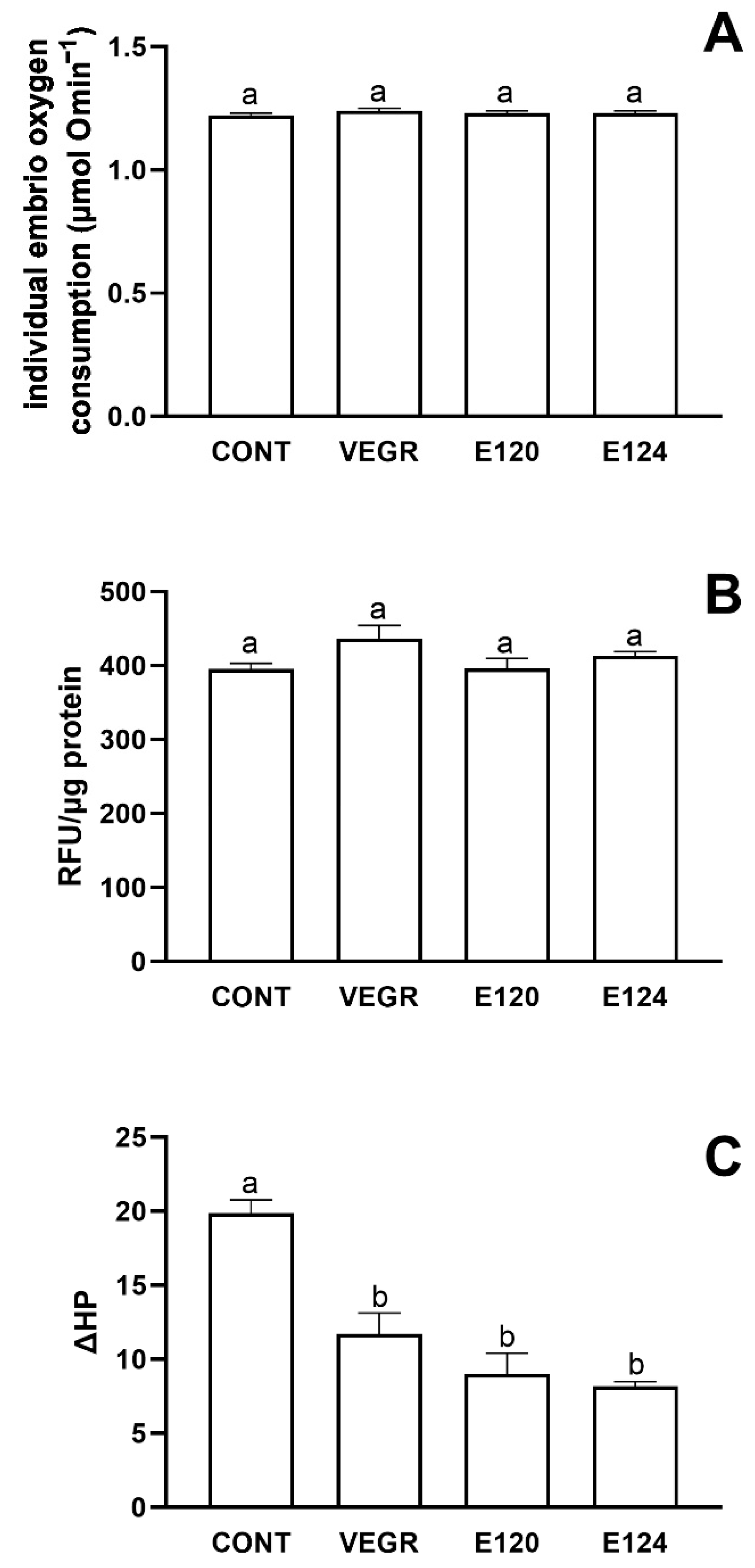
3.2. Artemia salina and Danio rerio Responses to Red Food Colors Exposure
4. Discussion
4.1. The Functional and Antioxidant Responses of the C. sativus Induced by Different Food Dyes
4.2. The Responses to Red Food Dyes Exposure in Artemia salina and Danio rerio
4.3. Effects of Red Food Dyes: Global Comparison and Conclusion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rymbai, H.; Sharma, R.R.; Srivastav, M. Biocolorants and its implications in Health and Food Industry—A Review. Int. J. Pharmacol. Res. 2011, 3, 2228–2244. [Google Scholar]
- Xu, D.P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.J.; Li, H.B. Natural Antioxidants in Foods and Medicinal Plants: Extraction, Assessment and Resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef] [PubMed]
- Schweiggert, R.M. Perspective on the Ongoing Replacement of Artificial and Animal-Based Dyes with Alternative Natural Pigments in Foods and Beverages. J. Agric. Food Chem. 2018, 66, 3074–3081. [Google Scholar] [CrossRef] [PubMed]
- Grand View Research. Global Natural Food Colors Market Size & Share, Industry Report, 2025: Natural Food Colors Market Size, Share & Trends Analysis Report by Product (Curcumin, Carotenoids, Anthocyanin, Carmine, Chlorophyllin), by Application, by Region, and Segment Forecasts 2018–2025. 2018. Available online: https://www.marketsandmarkets.com/Market-Reports/natural-colors-flavors-market-676.html?gclid=Cj0KCQjw1ZeUBhDyARIsAOzAqQKqRiXCfQn-TaRl5lmnRhjrBR-v3hwoaAYRP5ybns58jbHueqKYgtcaAu31EALw_wcB (accessed on 2 December 2021).
- Hussain, S.; Khan, N.; Gul, S.; Khan, S.; Khan, H. Contamination of Water Resources by Food Dyes and Its Removal Technologies. In Water Chemistry; Eyvaz, M., Yüksel, E., Eds.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Abe, F.R.; Gravato, C.; Soares, A.M.V.M.; de Oliveira, D.P. Biochemical approaches to assess oxidative stress induced by exposure to natural and synthetic dyes in early life stages in zebrafish. J. Toxicol. Environ. Health A 2017, 80, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Motta, C.M.; Simoniello, P.; Arena, C.; Capriello, T.; Panzuto, R.; Vitale, E.; Agnisola, C.; Tizzano, M.; Avallone, B.; Ferrandino, I. Effects of four food dyes on development of three model species, Cucumis sativus, Artemia salina and Danio rerio: Assessment of potential risk for the environment. Environ. Pollut. 2019, 253, 1126–1135. [Google Scholar] [CrossRef]
- Räisänen, R.; Primetta, A.; Nikunen, S.; Honkalampi, U.; Nygren, H.; Pihlava, J.M.; Vanden Berghe, I.; von Wright, A. Examining Safety of Biocolourants from Fungal and Plant Sources-Examples from Cortinarius and Tapinella, Salix and Tanacetum spp. and Dyed Woollen Fabrics. Antibiotics 2020, 9, 266. [Google Scholar] [CrossRef]
- Abe, F.R.; Soares, A.M.V.M.; de Oliveira, D.P.; Gravato, C. Toxicity of dyes to zebrafish at the biochemical level: Cellular energy allocation and neurotoxicity. Environ. Pollut. 2018, 235, 255–262. [Google Scholar] [CrossRef]
- Napolitano, G.; Venditti, P.; Fasciolo, G.; Esposito, D.; Uliano, E.; Agnisola, C. Acute hypoxia/reoxygenation affects muscle mitochondrial respiration and redox state as well as swimming endurance in zebrafish. J. Comp. Physiol. B 2019, 189, 97–108. [Google Scholar] [CrossRef]
- Cikcikoglu Yildirim, N.; Yaman, M. The usability of oxidative stress and detoxification biomarkers in Gammarus pulex for ecological risk assessment of textile dye methyl orange. Chem. Ecol. 2019, 35, 319–329. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlahogianni, T.; Dassenakis, M.; Scoullos, M. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar] [CrossRef]
- Venditti, P.; Napolitano, G.; Di Meo, S. Role of Mitochondria and Other ROS Sources in Hyperthyroidism-Linked Oxidative Stress. Immunol. Endocr. Metab. Agents Med. Chem. 2015, 15, 5–36. [Google Scholar] [CrossRef] [Green Version]
- Persoone, G.; Wells, P.G. Artemia in aquatic toxicology: A review. In Artemia. Research and Its Applications; Sorgeloos, P., Bengston, B.A., Decleir, W., Jaspers, E., Eds.; Universa Press: Wetteren, Belgium, 1987; pp. 259–275. [Google Scholar]
- Motta, C.; Cerciello, R.; De Bonis, S.; Mazzella, V.; Cirino, P.; Panzuto, R.; Ciaravolo, M.; Simoniello, P.; Toscanesi, M.; Trifuoggi, M.; et al. Potential toxicity of improperly discarded exhausted photovoltaic cells. Environ. Pollut. 2016, 216, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Capriello, T.; Grimaldi, M.C.; Cofone, R.; D’Aniello, S.; Ferrandino, I. Effects of aluminium and cadmium on hatching and swimming ability in developing zebrafish. Chemosphere 2019, 222, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Favorito, R.; Chiarelli, G.; Grimaldi, M.C.; Bonis, S.D.; Lancieri, M.; Ferrandino, I. Bioaccumulation of cadmium and its cytotoxic effect on zebrafish brain. Chem. Ecol. 2011, 27, 39–46. [Google Scholar] [CrossRef]
- Monaco, A.; Capriello, T.; Grimaldi, M.C.; Schiano, V.; Ferrandino, I. Neurodegeneration in zebrafish embryos and adults after cadmium exposure. J. List Eur. J. Histochem. 2017, 61, 2833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monaco, A.; Grimaldi, M.C.; Ferrandino, I. Aluminium chloride-induced toxicity in zebrafish larvae. J. Fish Dis. 2017, 40, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Capriello, T.; Visone, I.M.; Motta, C.; Ferrandino, I. Adverse effects of E150d on zebrafish development. Food Chem. Toxicol. 2021, 147, 111877. [Google Scholar] [CrossRef]
- Wang, X.; Sun, C.; Gao, S.; Wang, L.; Shuokui, H. Validation of germination rate and root elongation as indicator to assess phytotoxicity with Cucumis sativus. Chemosphere 2001, 44, 1711–1721. [Google Scholar] [CrossRef]
- Wu, X.; Wang, W.; Zhu, L. Enhanced organic contaminants accumulation in crops: Mechanisms, interactions with engineered nanomaterials in soil. Environ. Pollut. 2018, 240, 51–59. [Google Scholar] [CrossRef]
- Shakoor, S.; Nasar, A. Adsorptive treatment of hazardous methylene blue dye from artificially contaminated water using Cucumis sativus peel waste as a low-cost adsorbent. Groundw. Sustain. Dev. 2017, 5, 152–159. [Google Scholar] [CrossRef]
- Tkaczyk, A.; Mitrowska, K.; Posyniak, A. Synthetic organic dyes as contaminants of the aquatic environment and their implications for ecosystems: A review. Sci. Total Environ. 2020, 717, 137222. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Food Additives and Nutrient Sources Added to Food. Scientific Opinion on the re-evaluation of Ponceau 4R (E 124) as a food additive. EFSA J. 2009, 7, 1328. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS). Scientific Opinion on the re-evaluation of cochineal, carminic acid, carmines (E 120) as a food additive. EFSA J. 2015, 13, 4288. [Google Scholar]
- ANS. Statement on Allura Red AC and other sulphonated mono azo dyes authorised as food and feed additives. EFSA J. 2013, 11, 3234. [Google Scholar] [CrossRef]
- Leo, L.; Loong, C.; Ho, X.L.; Raman, M.F.B.; Suan, M.Y.T.; Loke, W.M. Occurrence of azo food dyes and their effects on cellular inflammatory responses. Nutrition 2018, 46, 36–40. [Google Scholar] [CrossRef]
- Koley, T.K.; Khan, Z.; Oulkar, D.; Singh, B.K.; Maurya, A.; Singh, B.; Banerjee, K. High resolution LC-MS characterization of phenolic compounds and the evaluation of antioxidant properties of a tropical purple radish genotype. Arab. J. Chem. 2020, 13, 1355–1366. [Google Scholar] [CrossRef]
- Lambert, J.D.; Elias, R.J. The antioxidant and pro-oxidant activities of green tea polyphenols: A role in cancer prevention. Arch. Biochem. Biophys. 2010, 501, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Gosetti, F.; Chiuminatto, U.; Mazzucco, E.; Calabrese, G.; Gennaro, M.C.; Marengo, E. Identification of photodegradation products of Allura Red AC (E129) in a beverage by ultra-high-performance liquid chromatography–quadrupole-time-of-flight mass spectrometry. Anal. Chim. Acta 2012, 746, 84–89. [Google Scholar] [CrossRef]
- Kovacic, P.; Somanathan, R. Novel, unifying mechanism for aromatic primary- amines (therapeutics, carcinogens and toxins): Electron transfer, reactive oxygen species, oxidative stress and metabolites. Med. Chem. Comm. 2011, 2, 106–112. [Google Scholar] [CrossRef]
- Sabatino, L.; Scordino, M.; Gargano, M.; Lazzaro, F.; Borzì, M.A.; Traulo, P.; Gagliano, G. Aminocarminic acid in E120-labelled food additives and beverages. Food Addit. Contam. Part B 2012, 5, 295–300. [Google Scholar] [CrossRef]
- Nojavan, S.; Tahmasebi, Z.; Bidarmanesh, T.; Behdad, H.; Nasiri-Aghdam, M.; Mansori, S.; Pourahadi, A. Electrically enhanced liquid-phase microextraction of three textile azo dyes from wastewater and plant samples. J. Sep. Sci. 2013, 36, 3256–3263. [Google Scholar] [CrossRef] [PubMed]
- Krishnaraju, A.V.; Rao, T.V.N.; Sundararaju, D.; Vanisree, M.; Tsay, H.S.; Subbaraju, G.V. Assessment of bioactivity of Indian medicinal plants using brine shrimp (Artemia salina) lethality assay. Int. J. Appl. Sci. Eng. 2005, 3, 125–134. [Google Scholar]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2020. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology, Plant Cell Membranes; Academic Press: Cambridge, MA, USA, 1987; pp. 350–382. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, G.; Sun, B.; Zhang, S.; Zhang, Y.; Liao, Y.; Zhou, Y.; Xia, X.; Shi, K.; Yu, J. Stimulated Leaf Dark Respiration in Tomato in an Elevated Carbon Dioxide Atmosphere. Sci. Rep. 2013, 3, 3433. [Google Scholar] [CrossRef] [Green Version]
- Sergiev, I.; Alexieva, V.; Karanov, E. Effect of spermine, atrazine and combination between them on some endogenous protective systems and stress markers in plants. Compt. Rend. Acad. Bulg. Sci. 1997, 51, 121–124. [Google Scholar]
- George, B.; Kaur, C.; Khurdiya, D.S.; Kapoor, H.C. Antioxidants in tomato (Lycopersium esculentum) as a function of genotype. Food Chem. 2004, 84, 45–51. [Google Scholar] [CrossRef]
- Costanzo, G.; Iesce, M.R.; Naviglio, D.; Ciaravolo, M.; Vitale, E.; Arena, C. Comparative Studies on Different Citrus Cultivars: A Revaluation of Waste Mandarin Components. Antioxidants 2020, 9, 517. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, E.; Mele, B.H.; Cataletto, P.R.; Turano, M.; Simoniello, P.; Micco, V.D. Suitability of Solanum lycopersicum L. ‘Microtom’ for growth in Bioregenerative Life Support Systems: Exploring the effect of high-LET ionising radiation on photosynthesis, leaf structure and fruit traits. Plant Bio 2019, 21, 615–626. [Google Scholar] [CrossRef]
- Vanhaecke, P.; Persoone, G.; Claus, C.; Sorgeloos, P. Proposal for a short-term toxicity test with Artemia nauplii. Ecotoxicol. Environ. Saf. 1981, 5, 382–387. [Google Scholar] [CrossRef]
- Gornall, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar] [CrossRef]
- Driver, A.S.; Kodavanti, P.R.S.; Mundy, W.R. Age-related changes in reactive oxygen species production in rat brain homogenates. Neurotoxicol. Teratol. 2000, 22, 175–181. [Google Scholar] [CrossRef]
- Napolitano, G.; Fasciolo, G.; Salbitani, G.; Venditti, P. Chlorella sorokiniana Dietary Supplementation Increases Antioxidant Capacities and Reduces Ros Release in Mitochondria of Hyperthyroid Rat Liver. Antioxidants 2020, 9, 883. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Tappel, A.L. A new sensitive assay for the measurement of hydroperoxides. Anal. Biochem. 1976, 76, 184–191. [Google Scholar] [CrossRef]
- European Parliament, Council of the European Union. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. Off. J. Eur. Union 2010, 276, 33–79. [Google Scholar]
- Arena, C.; De Maio, A.; De Nicola, F.; Santorufo, L.; Vitale, L.; Maisto, G. Assessment of eco-physiological performance of Quercus ilex L. leaves in urban area by an integrated approach. Water Air Soil Pollut. 2014, 225, 1824. [Google Scholar] [CrossRef]
- Navabpour, S.; Yamchi, A.; Bagherikia, S.; Kafi, H. Lead-induced oxidative stress and role of antioxidant defense in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2020, 26, 793–802. [Google Scholar] [CrossRef]
- Gonçalves, J.F.; Becker, A.G.; Cargnelutti, D.; Tabaldi, L.A.; Pereira, L.B.; Battisti, V.; Spanevello, R.M.; Morsch, V.M.; Nicoloso, F.T.; Schetinger, M.R.C. Cadmium toxicity causes oxidative stress and induces response of the antioxidant system in cucumber seedlings. Braz. J. Plant Physiol. 2007, 19, 223–232. [Google Scholar] [CrossRef]
- Jadhav, S.B.; Surwase, S.N.; Kalyani, D.C.; Gurav, R.G.; Jadhav, J.P. Biodecolorization of azo dye remazol orange by Pseudomonas aeruginosa BCH and toxicity (oxidative stress) reduction in Allium cepa root cells. Appl. Biochem. Biotechnol. 2012, 168, 1319–1334. [Google Scholar] [CrossRef]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Vitale, L.; Vitale, E.; Costanzo, G.; De Maio, A.; Arena, C. Photo-Protective mechanisms and the role of poly (ADP-Ribose) polymerase activity in a facultative CAM plant exposed to long-term water deprivation. Plants 2020, 9, 1192. [Google Scholar] [CrossRef]
- Geigenberger, P. Response of plant metabolism to too little oxygen. Curr. Opin. Plant Biol. 2003, 6, 247–256. [Google Scholar] [CrossRef]
- Hyson, D.A. A Comprehensive Review of apples and apple components and their relationship to human health. Adv. Nutr. 2011, 2, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Karjalainen, R.; Stewart, D.; McDougall, G.J.; Hilz, H.; Anttonen, M.; Saviranta, N.; Mattila, P.; Törrönen, R. Bioactive compounds in black currants (Ribes nigrum L.) and their potential health-promoting properties. In Proceedings of the International Symposium on Biotechnology of Fruit Species, Dresden, Germany, 1–5 September 2008. [Google Scholar]
- Manivannan, A.; Kim, J.H.; Kim, D.S.; Lee, E.S.; Lee, H.E. Deciphering the Nutraceutical Potential of Raphanus sativus—A Comprehensive Overview. Nutrients 2019, 11, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racchi, M.L. Antioxidant Defenses in Plants with Attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef]
- Kumar, A.; Prasad, M.N.V.; Sytar, O. Lead toxicity, defense strategies and associated indicative biomarkers in Talinum triangulare grown hydroponically. Chemosphere 2012, 89, 1056–1065. [Google Scholar] [CrossRef]
- Pospíšil, P. Production of Reactive Oxygen Species by Photosystem II as a Response to Light and Temperature Stress. Front. Plant Sci. 2016, 7, 1950. [Google Scholar] [CrossRef]
- Pospíšil, P.; Yamamoto, Y. Damage to photosystem II by lipid peroxidation products. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 457–466. [Google Scholar] [CrossRef]
- Demmig, B.; Björkman, O. Comparison of the effect of excessive light on chlorophyll fluorescence (77K) and photon yield of O2 evolution in leaves of higher plants. Planta 1987, 171, 171–184. [Google Scholar] [CrossRef]
- Arena, C.; Santorufo, L.; Cataletto, P.R.; Memoli, V.; Scudiero, R.; Maisto, G. Eco-physiological and antioxidant responses of holm oak (Quercus ilex L.) leaves to Cd and Pb. Water Air Soil Pollut. 2017, 228, 459. [Google Scholar] [CrossRef]
- Puvaneswari, N.; Muthukrishnan, J.; Gunasekaran, P. Toxicity assessment and microbial degradation of azo dyes. Indian. J. Exp. Biol. 2006, 44, 618–626. [Google Scholar]
- Tauqeer, H.M.; Ali, S.; Rizwan, M.; Ali, Q.; Saeed, R.; Iftikhar, U.; Ahmad, R.; Farid, M.; Abbasi, G.H. Phytoremediation of heavy metals by Alternanthera bettzickiana: Growth and physiological response. Ecotoxicol. Environ. Saf. 2016, 126, 138–146. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demmig-Adams, B.; Adams, W.W. The role of xanthophyll cycle carotenoids in the protection of photosynthesis. Trends Plant Sci. 1996, 1, 21–26. [Google Scholar] [CrossRef]
- Sant, K.E.; Sinno, P.P.; Jacobs, H.M.; Timme-Laragy, A.R. Nrf2a modulates the embryonic antioxidant response to perfluorooctanesulfonic acid (PFOS) in the zebrafish, Danio rerio. Aquat. Toxicol. 2018, 198, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Laszczyca, P.; Kawka-Serwecińska, E.; Witas, I.; Dolezych, B.; Migula, P. Iron ascorbate-stimulated lipid peroxidation in vitro. Why is the method controversial? Gen. Physiol. Biophys. 1995, 14, 3–18. [Google Scholar] [PubMed]
- Shaw, P.; Chattopadhyay, A. Nrf2–ARE signaling in cellular protection: Mechanism of action and the regulatory mechanisms. J. Cell. Physiol. 2020, 235, 3119–3130. [Google Scholar] [CrossRef] [PubMed]
- Kasai, S.; Shimizu, S.; Tatara, Y.; Mimura, J.; Itoh, K. Regulation of Nrf2 by Mitochondrial Reactive Oxygen Species in Physiology and Pathology. Biomolecules 2020, 10, 320. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, H.M.; Touraud, R.E.; Thomas, O. Aromatic amines from azo dye reduction: Status review with emphasis on direct UV spectrophotometric detection in textile industry wastewaters. Dye Pigment. 2004, 61, 121–139. [Google Scholar] [CrossRef]
- Seesuriyachan, P.; Takenaka, S.; Kuntiya, A.; Klayraung, S.; Murakami, S.; Aoki, K. Metabolism of azo dyes by Lactobacillus casei TISTR 1500 and effects of various factors on decolorization. Water Res. 2007, 41, 985–992. [Google Scholar] [CrossRef] [Green Version]
- Baldin, J.C.; Michelin, E.C.; Polizer, Y.J.; Rodrigues, I.; de Godoy, S.H.S.; Fregonesi, R.P.; Pires, M.A.; Carvalho, L.T.; Fávaro-Trindade, C.S.; de Lima, C.G.; et al. Microencapsulated jabuticaba (Myrciaria cauliflora) extract added to fresh sausage as natural dye with antioxidant and antimicrobial activity. Meat Sci. 2016, 118, 15–21. [Google Scholar] [CrossRef]
- Silva, M.C.; Souza, V.; Thomazini, M.; da Silva, E.R.; Smaniotto, T.; de Carvalho, R.A.; Genovese, M.I.; Favaro-Trindade, C.S. Use of the jabuticaba (Myrciaria cauliflora) depulping residue to produce a natural pigment powder with functional properties. LWT—Food Sci. Technol. 2014, 55, 203–209. [Google Scholar] [CrossRef]
- LCI. Is ‘Red’ Always ‘Red’? 4-aminocarminic acid as an accompanying substance in ‘carmine’. LCI Focus 2017, 4, 5. [Google Scholar]
- Stackley, K.D.; Beeson, C.C.; Rahn, J.J.; Chan, S.S.L. Bioenergetic Profiling of Zebrafish Embryonic Development. PLoS ONE 2011, 6, e25652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-García, D.; Wood, C.D.; Castro-Obregón, S.; Covarrubias, L. Reactive oxygen species: A radical role in development? Free Radic. Biol. Med. 2010, 49, 130–143. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CONT | VEGR | E120 | E124 | |
|---|---|---|---|---|
| Total biomass (g FW plant−1) | 0.410 ± 0.01 a | 0.352 ± 0.012 b | 0.451 ± 0.02 a | 0.414 ± 0.01 a |
| Fv/Fm | 0.810 ± 0.01 a | 0.814 ± 0.02 a | 0.812 ± 0.01 a | 0.814 ± 0.02 a |
| VEGR | E120 | E124 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Effect | Cucumis sativus | Artemia salina | Danio rerio | Cucumis sativus | Artemia salina | Danio rerio | Cucumis sativus | Artemia salina | Danio rerio |
| Functional performance | 0 | 0 | 0 | 0 | 0 | 0 | − | 0 | 0 |
| Pro-oxidant | 0 | + | 0 | − | + | 0 | + | + | 0 |
| Antioxidant | + | 0 | + | + | + | + | + | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Napolitano, G.; Motta, C.M.; Agnisola, C.; Venditti, P.; Fasciolo, G.; Ferrandino, I.; Capriello, T.; Vitale, E.; Costanzo, G.; Avallone, B.; et al. Commercial Red Food Dyes Preparations Modulate the Oxidative State in Three Model Organisms (Cucumis sativus, Artemia salina, and Danio rerio). Environments 2022, 9, 63. https://0-doi-org.brum.beds.ac.uk/10.3390/environments9050063
Napolitano G, Motta CM, Agnisola C, Venditti P, Fasciolo G, Ferrandino I, Capriello T, Vitale E, Costanzo G, Avallone B, et al. Commercial Red Food Dyes Preparations Modulate the Oxidative State in Three Model Organisms (Cucumis sativus, Artemia salina, and Danio rerio). Environments. 2022; 9(5):63. https://0-doi-org.brum.beds.ac.uk/10.3390/environments9050063
Chicago/Turabian StyleNapolitano, Gaetana, Chiara Maria Motta, Claudio Agnisola, Paola Venditti, Gianluca Fasciolo, Ida Ferrandino, Teresa Capriello, Ermenegilda Vitale, Giulia Costanzo, Bice Avallone, and et al. 2022. "Commercial Red Food Dyes Preparations Modulate the Oxidative State in Three Model Organisms (Cucumis sativus, Artemia salina, and Danio rerio)" Environments 9, no. 5: 63. https://0-doi-org.brum.beds.ac.uk/10.3390/environments9050063