Natural Genetic Variation for Grapevine Phenology as a Tool for Climate Change Adaptation


Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Fruiting Cutting Growing Conditions
2.3. Phenology Assessment
2.4. Harvest Sampling
2.5. Statistical Analysis
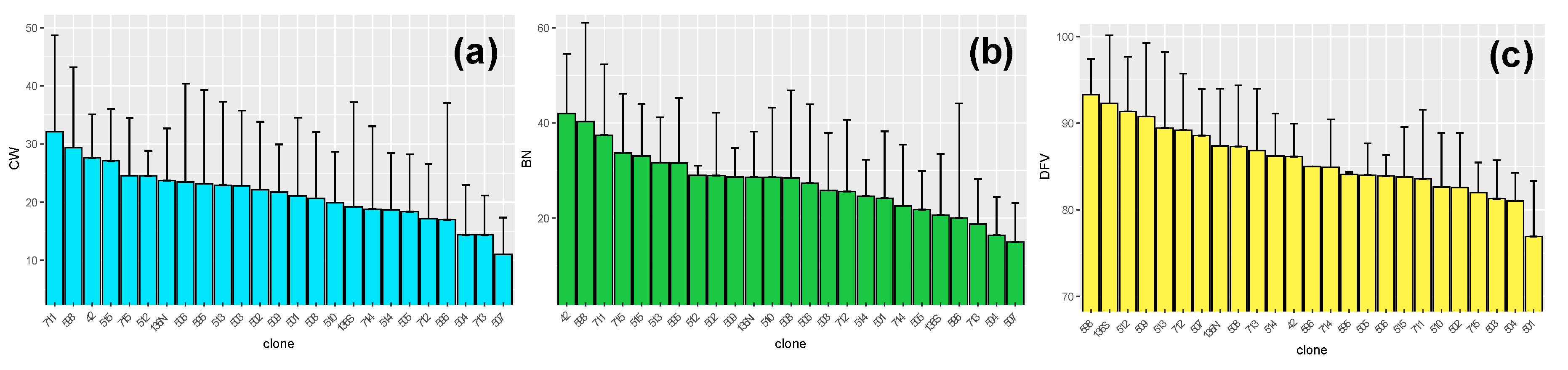
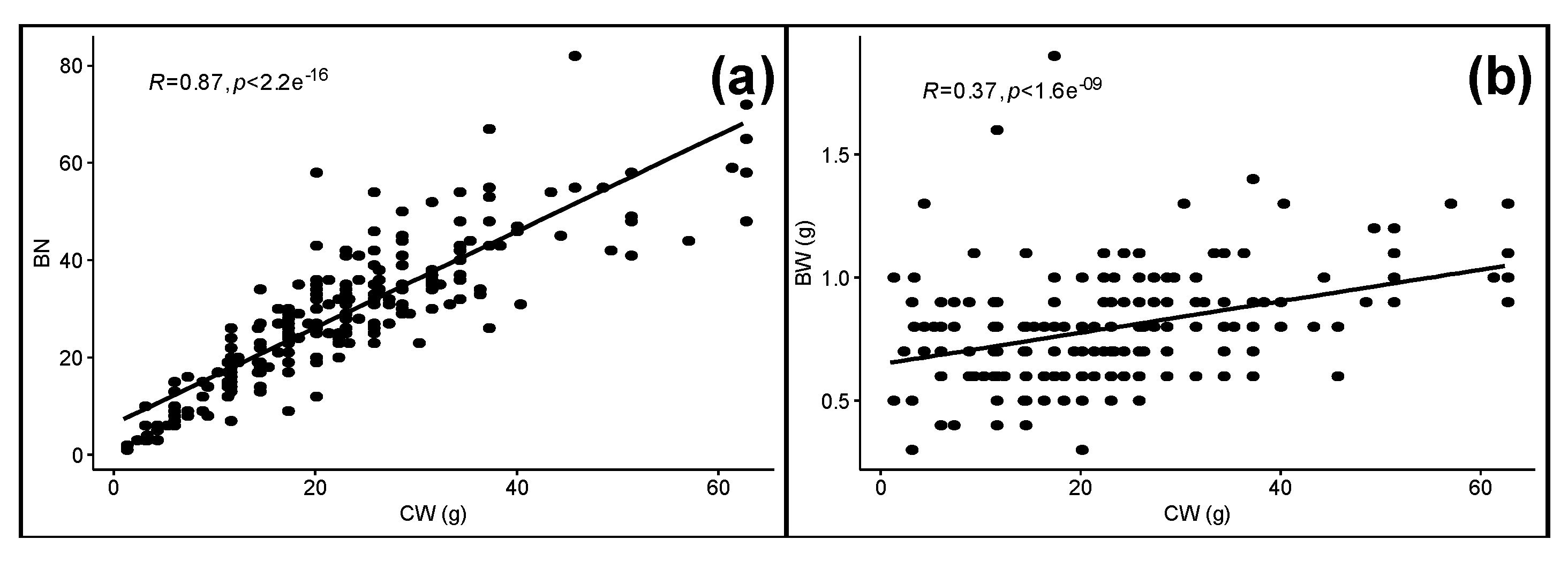
3. Results and Discussion
3.1. Fruiting Cuttings Allow Displaying the Variation for the Growing Conditions between ‘Malbec’ Clones
3.2. ‘Malbec’ Clones Show Significant Variation for Phenology and Berry Traits
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Voosen, P. New climate models predict a warming surge. Science (80-. ) 2019. [Google Scholar] [CrossRef]
- Carbonell-Bejerano, P.; Santa María, E.; Torres-Pérez, R.; Royo, C.; Lijavetzky, D.; Bravo, G.; Aguirreolea, J.; Sánchez-Díaz, M.; Antolín, M.C.C.; Martínez-Zapater, J.M.J.M. Thermotolerance Responses in Ripening Berries of Vitis vinifera L. cv Muscat Hamburg. Plant Cell Physiol. 2013, 54, 1200–1216. [Google Scholar] [CrossRef] [Green Version]
- Pastore, C.; Dal Santo, S.; Zenoni, S.; Movahed, N.; Allegro, G.; Valentini, G.; Filippetti, I.; Tornielli, G.B. Whole Plant Temperature Manipulation Affects Flavonoid Metabolism and the Transcriptome of Grapevine Berries. Front. Plant Sci. 2017, 8, 929. [Google Scholar] [CrossRef]
- Lecourieux, F.; Kappel, C.; Pieri, P.; Charon, J.; Pillet, J.; Hilbert, G.; Renaud, C.; Gomès, E.; Delrot, S.; Lecourieux, D. Dissecting the Biochemical and Transcriptomic Effects of a Locally Applied Heat Treatment on Developing Cabernet Sauvignon Grape Berries. Front. Plant Sci. 2017, 8, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Movahed, N.; Pastore, C.; Cellini, A.; Allegro, G.; Valentini, G.; Zenoni, S.; Cavallini, E.; D’Incà, E.; Tornielli, G.B.; Filippetti, I. The grapevine VviPrx31 peroxidase as a candidate gene involved in anthocyanin degradation in ripening berries under high temperature. J. Plant Res. 2016, 129, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Sadras, V.O.; Moran, M.A. Nonlinear effects of elevated temperature on grapevine phenology. Agric. For. Meteorol. 2013, 173, 107–115. [Google Scholar] [CrossRef]
- Sadras, V.O.; Petrie, P.R. Climate shifts in south-eastern Australia: Early maturity of Chardonnay, Shiraz and Cabernet Sauvignon is associated with early onset rather than faster ripening. Aust. J. Grape Wine Res. 2011, 17, 199–205. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Kizildeniz, T.; Vučetić, V.; Dai, Z.; Luedeling, E.; van Leeuwen, C.; Gomès, E.; Pascual, I.; Irigoyen, J.J.; Morales, F.; et al. Sensitivity of Grapevine Phenology to Water Availability, Temperature and CO2 Concentration. Front. Environ. Sci. 2016, 4, 48. [Google Scholar] [CrossRef]
- Duchêne, E.; Huard, F.; Dumas, V.; Schneider, C.; Merdinoglu, D. The challenge of adapting grapevine varieties to climate change. Clim. Res. 2010, 41, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Torregrosa, L.; Bigard, A.; Doligez, A.; Lecourieux, D.; Rienth, M.; Luchaire, N.; Pieri, P.; Chatbanyong, R.; Shahood, R.; Farnos, M.; et al. Developmental, molecular and genetic studies on grapevine response to temperature open breeding strategies for adaptation to warming. J. Int. Sci. Vigne Vin 2017, 51, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Agasse, A.; Delrot, S.; Geros, H. Biochemical changes throughout grape berry development and fruit and wine quality. Food 2007, 1, 1–22. [Google Scholar]
- Caballero, A.; Segura, A. The quest for lower alcoholic wines. Microb. Biotechnol. 2017, 10, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Duchêne, E. How can grapevine genetics contribute to the adaptation to climate change? OENO One 2016, 50. [Google Scholar] [CrossRef] [Green Version]
- Van Leeuwen, C.; Darriet, P. The Impact of Climate Change on Viticulture and Wine Quality. J. Wine Econ. 2016, 11, 150–167. [Google Scholar] [CrossRef] [Green Version]
- Arrizabalaga, M.; Morales, F.; Oyarzun, M.; Delrot, S.; Gomès, E.; Irigoyen, J.J.; Hilbert, G.; Pascual, I. Tempranillo clones differ in the response of berry sugar and anthocyanin accumulation to elevated temperature. Plant Sci. 2018, 267, 74–83. [Google Scholar] [CrossRef] [PubMed]
- This, P.; Lacombe, T.; Thomas, M.R. Historical origins and genetic diversity of wine grapes. Trends Genet 2006, 22, 511–519. [Google Scholar] [CrossRef]
- Carbonell-Bejerano, P.; Royo, C.; Torres-Pérez, R.; Grimplet, J.; Fernandez, L.; Franco-Zorrilla, J.M.J.M.; Lijavetzky, D.; Baroja, E.; Martínez, J.; García-Escudero, E.; et al. Catastrophic unbalanced genome rearrangements cause somatic loss of berry color in grapevine. Plant Physiol. 2017, 175, 786–801. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Zapater, J.M.M.; Carmona, M.J.J.; Díaz-Riquelme, J.; Fernández, L.; Lijavetzky, D. Grapevine genetics after the genome sequence: Challenges and limitations. Aust. J. Grape Wine Res. 2010, 16, 33–46. [Google Scholar] [CrossRef]
- Mullins, M.G.; Rajasekaran, K. Fruiting Cuttings - Revised Method for Producing Test Plants of Grapevine Cultivars. Am. J. Enol. Vitic. 1981, 32, 35–40. [Google Scholar]
- Ollat, N.; Geny, L.; Soyer, J.-P. Grapevine fruiting cuttings: Validation of an experimental system to study grapevine physiology. I. Main vegetative characteristics. OENO One 1998, 32, 1. [Google Scholar] [CrossRef]
- Lebon, G.; Duchene, E.; Brun, O.; Clement, C. Phenology of Flowering and Starch Accumulation in Grape (Vitis vinifera L.) Cuttings and Vines. Ann Bot 2005, 95, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Vaillant-Gaveau, N.; Maillard, P.; Wojnarowiez, G.; Gross, P.; Clément, C.; Fontaine, F. Inflorescence of grapevine (Vitis vinifera L.): A high ability to distribute its own assimilates. J. Exp. Bot. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.W.; Léon, C.; Feil, R.; Lunn, J.E.; Delrot, S.; Gomès, E. Metabolic profiling reveals coordinated switches in primary carbohydrate metabolism in grape berry (Vitis vinifera L.), a non-climacteric fleshy fruit. J. Exp. Bot. 2013, 64, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Kizildeniz, T.; Pascual, I.; Irigoyen, J.J.; Morales, F. Using fruit-bearing cuttings of grapevine and temperature gradient greenhouses to evaluate effects of climate change (elevated CO2 and temperature, and water deficit) on the cv. red and white Tempranillo. Yield and must quality in three consecutive growin. Agric. Water Manag. 2018, 202, 299–310. [Google Scholar] [CrossRef]
- Ollat, N.; Gaudillere, J.P. The Effect of Limiting Leaf Area During Stage I of Berry Growth on Development and Composition of Berries of Vitis vinifera L. cv. Cabernet Sauvignon. Am. J. Enol. Vitic. 1998, 49, 251–258. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Found. Stat. Comput. 2019. Available online: http://finzi.psych.upenn.edu/R/library/dplR/doc/intro-dplR.pdf (accessed on 26 July 2020).
- Rstudio Team. RStudio: Integrated development for, R. RStudio, Inc., Boston MA. RStudio 2016, 42, 14. [Google Scholar]
- Royston, P. Remark AS R94: A Remark on Algorithm AS 181: The W-test for Normality. J. R. Stat. Soc. Ser. C (Appl. Stat.) 1995, 44, 547–551. [Google Scholar] [CrossRef]
- Hollander, M.; Wolfe, D.A.; Chicken, E. Nonparametric Statistical Methods, 3rd ed.; Hollander, M., Wolfe, D.A., Chicken, E., Eds.; John Wiley & Sons: New York, NY, USA, 2003; ISBN 9780470387375. [Google Scholar]
- Muñoz, C.J.; Gomez-Talquenca, S.; Chialva, C.; Ibáñez, J.; Martinez-Zapater, J.M.J.M.; Peña-Neira, A.; Lijavetzky, D.; Pena-Neira, A.; Lijavetzky, D. Relationships among gene expression and anthocyanin composition of Malbec grapevine clones. J. Agric. Food Chem. 2014, 62, 6716–6725. [Google Scholar] [CrossRef]
- Tortosa, I.; Escalona, J.M.; Douthe, C.; Pou, A.; Garcia-Escudero, E.; Toro, G.; Medrano, H. The intra-cultivar variability on water use efficiency at different water status as a target selection in grapevine: Influence of ambient and genotype. Agric. Water Manag. 2019, 223, 105648. [Google Scholar] [CrossRef]
- Grimplet, J.; Tello, J.; Laguna, N.; Ibáñez, J. Differences in Flower Transcriptome between Grapevine Clones Are Related to Their Cluster Compactness, Fruitfulness, and Berry Size. Front. Plant Sci. 2017, 8, 632. [Google Scholar] [CrossRef] [PubMed]
- Pelsy, F.; Hocquigny, S.; Moncada, X.; Barbeau, G.; Forget, D.; Hinrichsen, P.; Merdinoglu, D. An extensive study of the genetic diversity within seven French wine grape variety collections. Theor. Appl. Genet. 2010, 120, 1219–1231. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, M.R.; Sabalza, F.C.; Sarasa, A.S.; Abad, F.J.; Zapater, J.M.M.; Marcos, J.I. Intra-varietal diversity for agronomic traits in’Garnacha Blanca’. Vitis J. Grapevine Res. 2019, 58, 33–35. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Clone | Start (n) | Rooting (n) | (%) | Fruit Set (n) | (%) | Harvest (n) | (%) |
|---|---|---|---|---|---|---|---|
| 501 | 47 | 32 | 68 | 25 | 78 | 22 | 88 |
| 502 | 46 | 32 | 70 | 18 | 56 | 16 | 89 |
| 503 | 43 | 26 | 60 | 15 | 58 | 11 | 73 |
| 504 | 42 | 21 | 50 | 8 | 38 | 6 | 75 |
| 505 | 46 | 25 | 54 | 8 | 32 | 5 | 63 |
| 506 | 55 | 41 | 75 | 23 | 56 | 21 | 91 |
| 507 | 55 | 43 | 78 | 15 | 35 | 12 | 80 |
| 508 | 49 | 33 | 67 | 11 | 33 | 10 | 91 |
| 510 | 54 | 32 | 59 | 17 | 53 | 16 | 94 |
| 513 | 50 | 31 | 62 | 11 | 35 | 11 | 100 |
| 514 | 50 | 14 | 28 | 7 | 50 | 6 | 86 |
| 515 | 48 | 30 | 63 | 16 | 53 | 14 | 88 |
| 136_S | 42 | 22 | 52 | 12 | 55 | 10 | 83 |
| 136_N | 39 | 20 | 51 | 9 | 45 | 8 | 89 |
| 711 | 52 | 20 | 38 | 9 | 45 | 9 | 100 |
| 712 | 50 | 40 | 80 | 19 | 48 | 19 | 100 |
| 713 | 53 | 25 | 47 | 16 | 64 | 15 | 94 |
| 714 | 56 | 33 | 59 | 11 | 33 | 11 | 100 |
| 42 | 41 | 20 | 49 | 8 | 40 | 8 | 100 |
| 598 | 58 | 34 | 59 | 14 | 41 | 13 | 93 |
| 595 | 56 | 41 | 73 | 11 | 27 | 11 | 100 |
| mean | 49 | 29 | 59 | 13 | 46 | 12 | 89 |
| sdt | 6 | 8 | 13 | 5 | 13 | 5 | 10 |
| max | 58 | 43 | 80 | 25 | 78 | 22 | 100 |
| min | 39 | 14 | 28 | 7 | 27 | 5 | 63 |
| Trait | p-Value | H | df |
|---|---|---|---|
| DFV | <0.001 | 81 | 20 |
| CW | <0.001 | 40 | 20 |
| BN | <0.001 | 48 | 20 |
| BW | 0.03 | 33 | 20 |
| Brix | 0.01 | 37 | 20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Houten, S.; Muñoz, C.; Bree, L.; Bergamín, D.; Sola, C.; Lijavetzky, D. Natural Genetic Variation for Grapevine Phenology as a Tool for Climate Change Adaptation. Appl. Sci. 2020, 10, 5573. https://0-doi-org.brum.beds.ac.uk/10.3390/app10165573
van Houten S, Muñoz C, Bree L, Bergamín D, Sola C, Lijavetzky D. Natural Genetic Variation for Grapevine Phenology as a Tool for Climate Change Adaptation. Applied Sciences. 2020; 10(16):5573. https://0-doi-org.brum.beds.ac.uk/10.3390/app10165573
Chicago/Turabian Stylevan Houten, Silvina, Claudio Muñoz, Laura Bree, Daniel Bergamín, Cristobal Sola, and Diego Lijavetzky. 2020. "Natural Genetic Variation for Grapevine Phenology as a Tool for Climate Change Adaptation" Applied Sciences 10, no. 16: 5573. https://0-doi-org.brum.beds.ac.uk/10.3390/app10165573