Effects of di-(2-ethylhexyl) phthalate on Transcriptional Expression of Cellular Protection-Related HSP60 and HSP67B2 Genes in the Mud Crab Macrophthalmus japonicus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Preparation of M. japonicus Individuals
2.3. DEHP Exposure Experiments
2.4. Total RNA Extraction and cDNA Synthesis
2.5. Gene Expression Analysis Using Quantitative Reverse-Transcription PCR (RT-PCR) Amplification
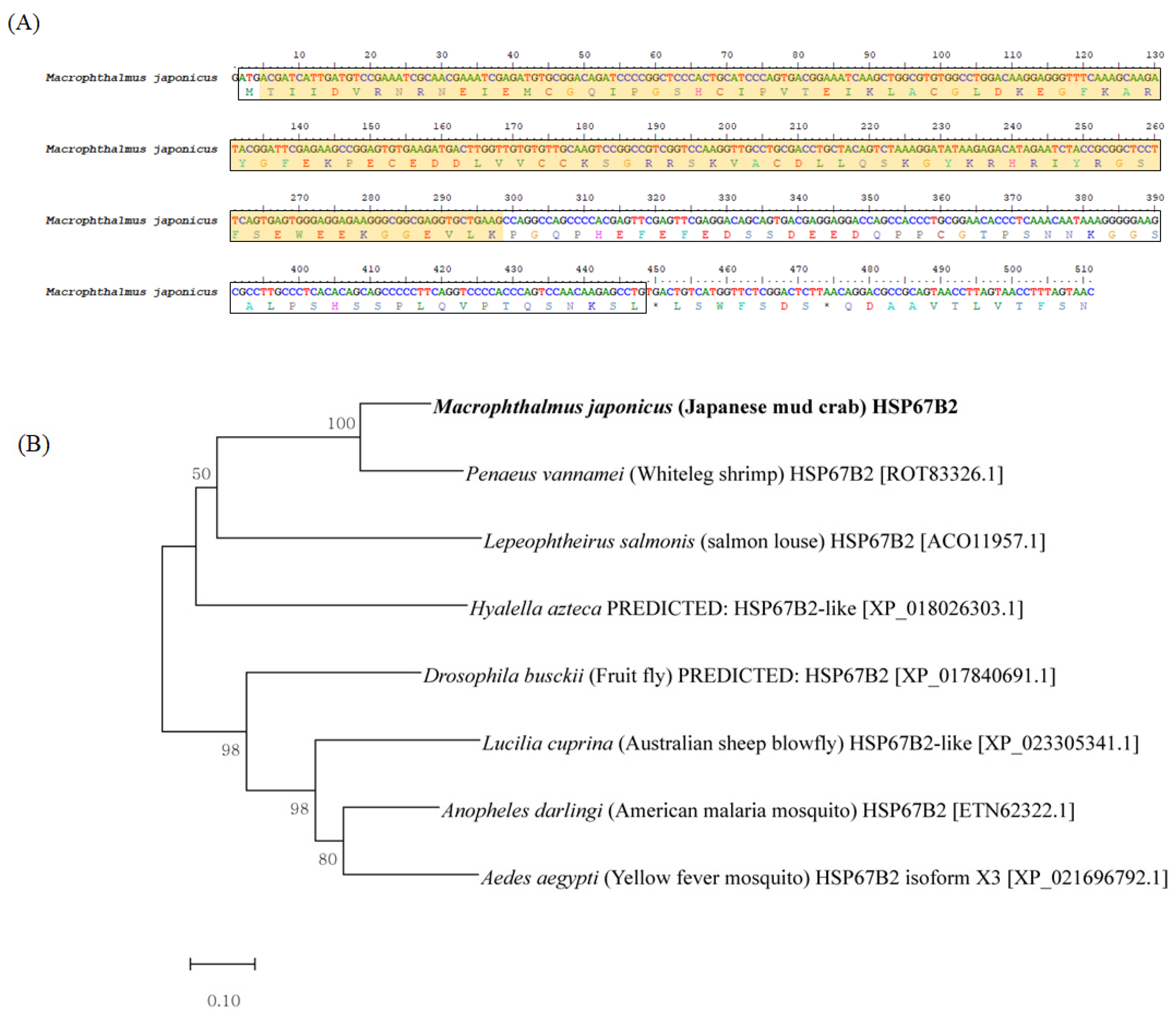
2.6. M. japonicus Hsp Identification and Bioinformatics Analysis
2.7. Statistical Analysis
3. Results
3.1. Characterization of Mj-HSP60 and Mj-HSP67B2 in M. japonicus
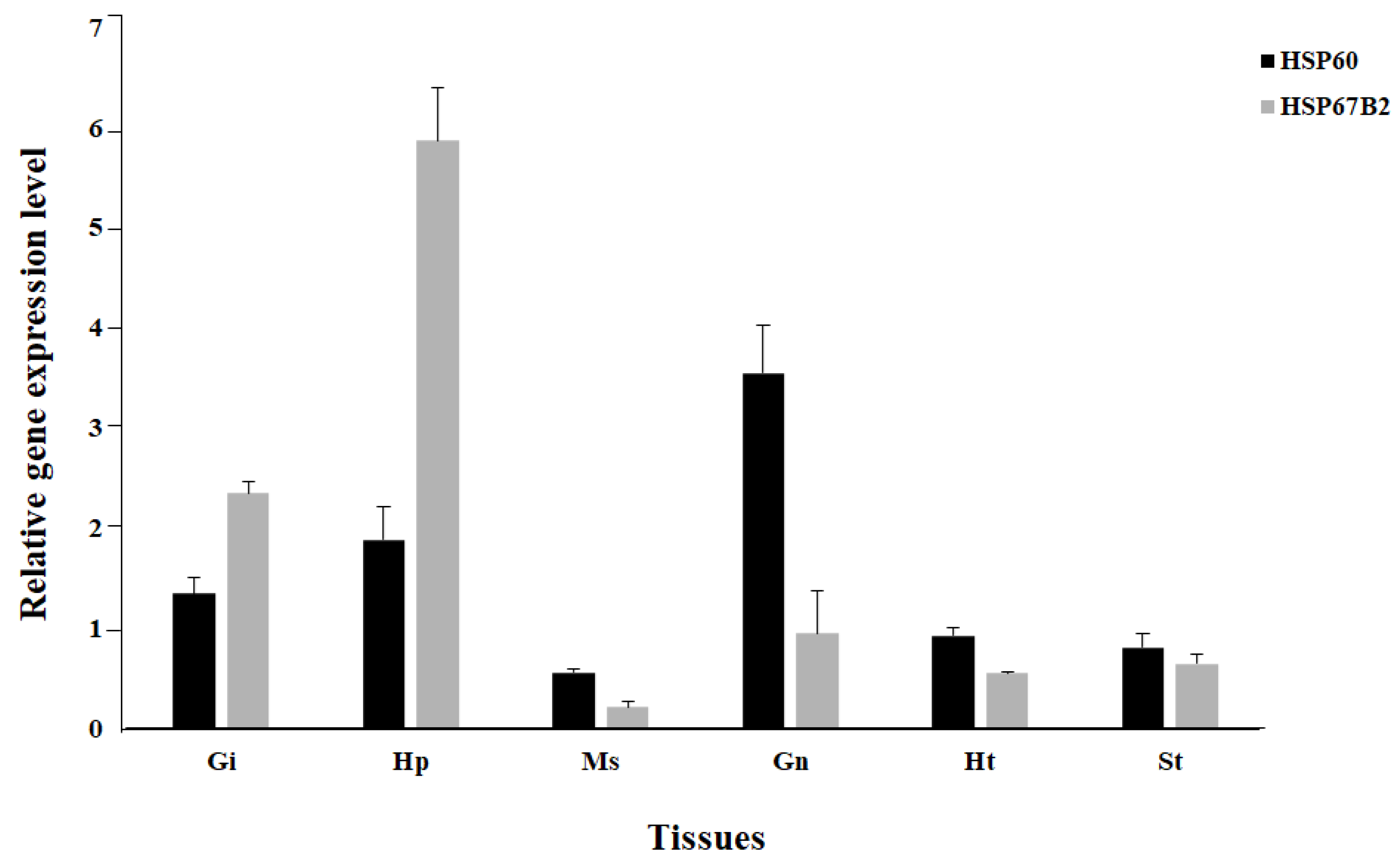
3.2. Expression Analysis of Mj-HSP60 and Mj-HSP67B2 in Various Tissues of M. japonicus
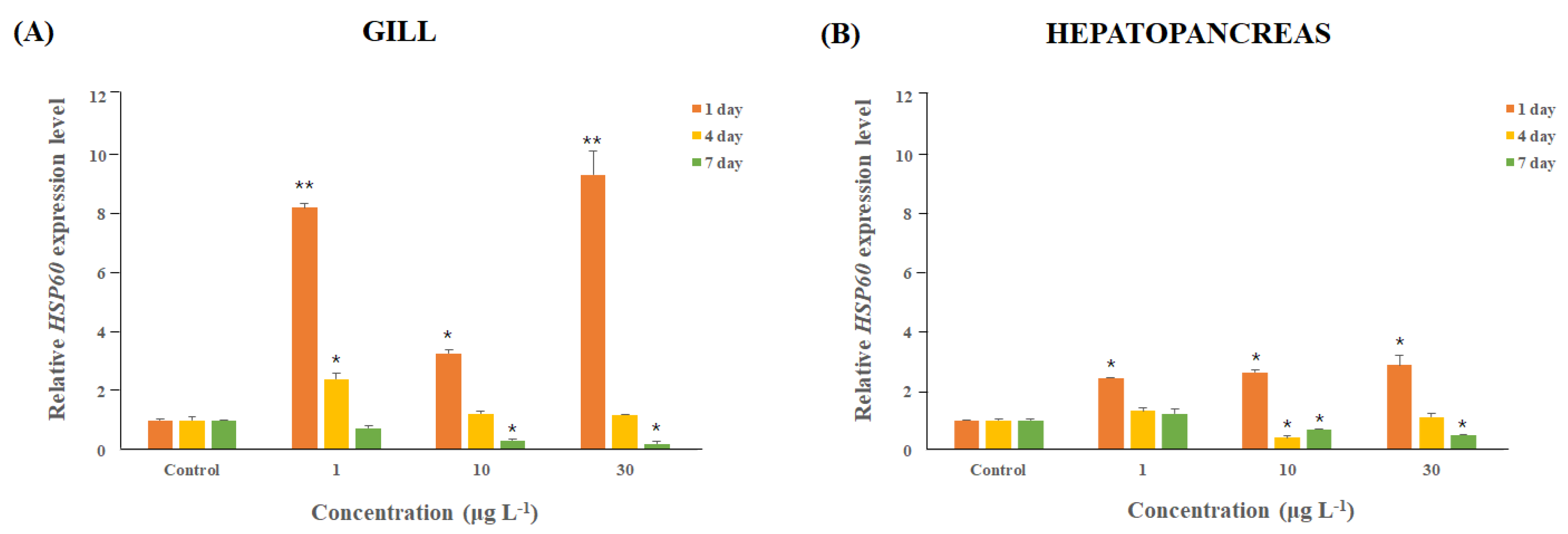
3.3. M. japonicus Mj-HSP60 Expression Changes after DEHP Exposure
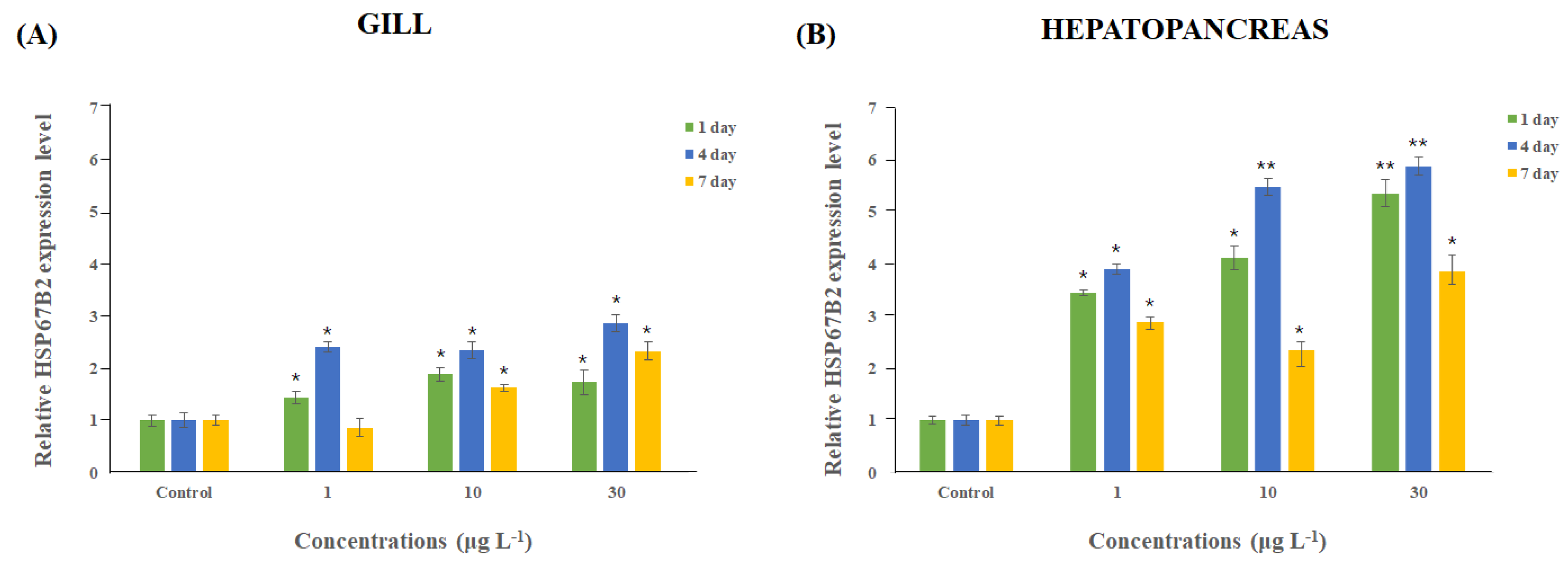
3.4. Variation in Expression of Mj-HSP67B2 after DEHP Exposure in M. japonicus
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Park, K.; Kwak, I.S. Molecular effects of endocrine-disrupting chemicals on the Chironomus riparius estrogen-related receptor gene. Chemosphere 2010, 79, 934–941. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, S.; Lim, J.E.; Moon, H.B. Occurrence and emission of phthalates and non-phthalate plasticizers in sludge from wastewater treatment plants in Korea. Sci. Total Environ. 2019, 692, 354–360. [Google Scholar] [CrossRef]
- Kim, S.; Lee, Y.S.; Moon, H.B. Occurrence, distribution, and sources of phthalates and non-phthalate plasticizers in sediment from semi-enclosed bays of Korea. Mar. Pollut. Bull. 2020, 151, 110824. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yin, P.; Zhao, L. Phthalate esters in water and surface sediments of the Pearl River estuary: Distribution, ecological, and human health risks. Environ. Sci. Pollut. Res. 2016, 23, 19341–19349. [Google Scholar] [CrossRef] [PubMed]
- Saeed, T.; Al-Jandal, N.; Abusam, A.; Taqi, H.; Al-Khabbaz, A.; Zafar, J. Sources and levels of endocrine disrupting compounds (EDCs) in Kuwait’s coastal areas. Mar. Pollut. Bull. 2017, 118, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.F.; Chen, C.W.; Ju, Y.R.; Dong, C.D. Determination and assessment of phthalate esters content in sediments from Kaohsiung Harbor, Taiwan. Mar. Pollut. Bull. 2017, 124, 767–774. [Google Scholar] [CrossRef]
- Paluselli, A.; Fauvelle, V.; Schmidt, N.; Galgani, F.; Net, S.; Sempéré, R. Distribution of phthalates in Marseille Bay (NW Mediterranean Sea). Sci. Total Environ. 2018, 621, 578–587. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.M.; Zhang, H.H.; Zou, Y.W.; Yang, G.P. Distribution and ecotoxicological state of phthalate esters in the sea-surface microlayer, seawater and sediment of the Bohai Sea and the Yellow Sea. Environ. Pollut. 2018, 240, 235–247. [Google Scholar] [CrossRef]
- Park, J.; Park, C.; Gye, M.C.; Lee, Y. Assessment of endocrine-disrupting activities of alternative chemicals for bis (2-ethylhexyl)phthalate. Environ. Res. 2019, 172, 10–17. [Google Scholar] [CrossRef]
- Park, K.; Jo, H.; Kim, D.K.; Kwak, I.S. Environmental pollutants impair transcriptional regulation of the vitellogenin gene in the burrowing mud crab (Macrophthalmus japonicus). Appl. Sci. 2019, 9, 1401. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Nikapitiya, C.; Kim, W.S.; Kwak, T.S.; Kwak, I.S. Changes of exoskeleton surface roughness and expression of crucial participation genes for chitin formation and digestion in the mud crab (Macrophthalmus japonicus) following the antifouling biocide irgarol. Ecotoxicol. Environ. Saf. 2016. 132, 186–195. [CrossRef]
- Kitaura, J.; Nishida, M.; Wada, K. Genetic and behavioral diversity in the Macrophthalmus japonicus species complex (Crustacea: Brachyura: Ocypodidae). Mar. Biol. 2002, 140, 1–8. [Google Scholar]
- Park, K.; Kwak, T.S.; Kim, W.S.; Kwak, I.S. Changes in exoskeleton surface roughness and expression of chitinase genes in mud crab Macrophthalmus japonicus following heavy metal differences of estuary. Mar. Pollut. Bull. 2019, 138, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Nikapitiya, C.; Kim, W.S.; Park, K.; Kim, J.; Lee, M.O.; Kwak, I.S. Chitinase gene responses and tissue sensitivity in an intertidal mud crab (Macrophthalmus japonicus) following low or high salinity stress. Cell Stress Chaperones. 2015, 20, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Nikapitiya, C.; Kim, W.S.; Park, K.; Kwak, I.S. Identification of potential markers and sensitive tissues for low or high salinity stress in an intertidal mud crab (Macrophthalmus japonicus). Fish Shellfish Immunol. 2014, 41, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Kwak, I.S. Characterize and gene expression of heat shock protein 90 in marine crab Charybdis japonica following bisphenol A and 4-nonylphenol exposures. Environ. Health Toxicol. 2014, 29, e2014002. [Google Scholar] [CrossRef]
- Park, K.; Kim, W.S.; Kwak, I.S. Endocrine-disrupting chemicals impair the innate immune prophenoloxidase system in the intertidal mud crab, Macrophthalmus japonicus. Fish Shellfish Immunol. 2019, 87, 322–332. [Google Scholar] [CrossRef]
- Díaz, F.; Orobio, R.F.; Chavarriaga, P.; Toro-Perea, N. Differential expression patterns among heat-shock protein genes and thermal responses in the whitefly Bemisia tabaci (MEAM 1). J. Therm. Biol. 2015, 52, 199–207. [Google Scholar] [CrossRef]
- Wu, J.; Liu, T.; Rios, Z.; Mei, Q.; Lin, X.; Cao, S. Heat Shock Proteins and Cancer. Trends Pharmacol. Sci. 2017, 38, 226–256. [Google Scholar] [CrossRef]
- Huang, L.H.; Le, K. Cloning and interspecific altered expression of heat shock protein genes in two leafminer species in response to thermal stress. Insect Mol. Biol. 2007, 16, 491–500. [Google Scholar] [CrossRef]
- Liu, T.; Daniels, C.K.; Cao, S. Comprehensive review on the HSC70 functions, interactions with related molecules and involvement in clinical diseases and therapeutic potential. Pharmacol. Ther. 2012, 136, 354–374. [Google Scholar] [CrossRef] [PubMed]
- Macario, A.J.; Conway de Macario, E. Molecular chaperones: Multiple functions, pathologies, and potential applications. Front. Biosci. 2007, 12, 2588–2600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macario, A.J.; Macario, C.E. Sick chaperones, cellular stress, and disease. N. Engl. J. Med. 2005, 353, 1489–1501. [Google Scholar] [CrossRef]
- Chang, H.C.; Tang, Y.C.; Hayer-Hartl, M.; Hartl, U.C. SnapShot: Molecular chaperones. Part I. Cell 2007, 128, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.C.; Chang, H.C.; Hayer-Hartl, M.; Hartl, U.C. SnapShot: Molecular chaperones. Part II. Cell 2007, 128, 412. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.Y.; Shen, Y.B.; Fu, J.J.; Liu, F.; Guo, S.Z.; Yang, X.M.; Li, J.L. Molecular cloning, characterization and expression patterns of HSP60 in the grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2011, 31, 864–870. [Google Scholar] [CrossRef]
- Ning, M.X.; Xiu, Y.J.; Bi, J.X.; Liu, Y.H.; Hou, L.B.; Ding, Z.F.; Gu, W.; Wang, W.; Meng, Q.G. Interaction of heat shock protein 60 (HSP60) with microRNA in Chinese mitten crab during Spiroplasma eriocheiris infection. Dis. Aquat. Organ. 2017, 125, 207–215. [Google Scholar] [CrossRef]
- Wang, D.; Li, S.; Li, F. Screening of genes regulated by Relish in Chinese shrimp Fenneropenaeus chinensis. Dev. Comp. Immunol. 2013, 41, 209–216. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2 –ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bairoch, A.; Bucher, P.; Hofmann, K. The PROSITE database, its status in 1997. Nucleic Acids Res. 1997, 25, 217–221. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Park, K.; Nikapitiya, C.; Kwak, I.S. Identification and expression of proteolysis response genes for Macrophthalmus japonicus exposure to irgarol toxicity. Ann. Limnol. Int. J. Limnol. 2016, 52, 65–74. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlova, T.; Perezgasga, T.; Reynaud, E.; Zurita, M. The Drosophila melanogaster homologue of the hsp60 gene is encoded by the essential locus 1 (1)10Ac and is differentially expressed during fly development. Dev. Genes Evol. 1997, 207, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, A.; Wilhem, B.; Seitz, J. Expression of mitochondrial marker proteins during spermatogenesis. Hum. Reprod. Update 1999, 5, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Timakov, B.; Zhang, P. The hsp60B gene in Drosophila melanogaster is essential for the spermatid individualization process. Cell Stress Chaperones 2001, 6, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Choresh, O.; Ron, E.; Loya, Y. The 60-kDa heat shock protein (HSP60) of the sea anemone Anemonia viridis: A potential early warning system for monitoring environmental changes. Mar. Biotechnol. 2001, 3, 501–508. [Google Scholar] [CrossRef] [Green Version]
- Kammenga, J.E.; Arts, M.S.J.; Oude-Breuil, W.J.M. HSP60 as a potential biomarker of toxic stress in the Nematode Plectus acuminatus. Arch. Environ. Contam. Toxicol. 1998, 34, 253–258. [Google Scholar] [CrossRef]
- Chen, Z.; Christina, C.C.H.; Zhang, J.; Cao, L.; Chen, L.; Zhou, L.; Jin, Y.; Ye, H.; Deng, C.; Dai, Z.; et al. Transcriptomic and genomic evolution under constant cold in Antarctic notothenioid fish. Proc. Natl. Acad. Sci. USA 2008, 105, 12944–12949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vabulas, R.M.; Ahmad-Nejad, P.; Da Costa, C.; Miethke, T.; Kirschning, C.J.; Haucker, H.; Wagner, H. Endocytosed HSP60s use toll-like receptor 2 (TLR2) and TLR4 to activate the toll/interleukin-1 receptor signaling pathway in innate immune cells. J. Biol. Chem. 2001, 276, 31332–31339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindquist, S.; Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Parsell, D.A.; Lindquist, S. The function of heat-shock proteins in stress tolerance: Degradation and reactivation of damaged proteins. Annu. Rev. Genet. 1993, 27, 437–496. [Google Scholar] [CrossRef] [PubMed]
- Brunet, M.; Arnaud, J.; Mazz, J. Gut structure and digestive cellular process in marine crustaceans. Oceanogra. Mar. Biol. 1990, 32, 335–367. [Google Scholar]
- Rungrassamee, W.; Leelatanawit, R.; Jiravanichpaisal, P.; Klinbunga, S.; Karoonuthaisiri, N. Expression and distribution of three heat shock protein genes under heat shock stress and under exposure to Vibrio harveyi in Penaeus monodon. Dev. Comp. Immunol. 2010, 34, 1082–1089. [Google Scholar] [CrossRef]
- Chaurasia, M.K.; Nizam, F.; Ravichandran, G.; Arasu, M.V.; Al-Dhabi, N.A.; Arshad, A.; Elumalai, P.; Arockiaraj, J. Molecular importance of prawn large heat shock proteins 60, 70 and 90. 2016. Fish Shellfish Immunol. 2016, 48, 228–238. [Google Scholar] [CrossRef]
- Luan, W.; Li, F.; Zhang, J.; Wen, R.; Li, Y.; Xiang, J. Identification of a novel inducible cytosolic Hsp70 gene in Chinese shrimp Fenneropenaeus chinensis and comparison of its expression with the cognate Hsc70 under different stresses. Cell Stress Chaperones 2010, 15, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Kwak, I.S. Characterization of heat shock protein 40 and 90 in Chironomus riparius larvae: Effects of di (2-ethylhexyl) phthalate exposure on gene expressions and mouthpart deformities. Chemosphere 2008, 74, 89–95. [Google Scholar] [CrossRef]
- Morales, M.; Planelló, R.; Martínez-Paz, P.; Herrero, O.; Cortés, E.; Martínez-Guitarte, J.L.; Morcillo, G. Characterization of Hsp70 gene in Chironomus riparius: Expression in response to endocrine disrupting pollutants as a marker of ecotoxicological stress. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 150–158. [Google Scholar] [CrossRef]
- Tang, T.; Sun, H.; Li, Y.; Chen, P.; Liu, F. MdRDH1, a HSP67B2-like rhodanese homologue plays a positive role in maintaining redox balance in Musca domestica. Mol. Immunol. 2019, 107, 115–122. [Google Scholar] [CrossRef]
- Xu, Q.; Qin, Y. Molecular cloning of heat shock protein 60 (PtHSP60) from Portunus trituberculatus and its expression response to salinity stress. Cell Stress Chaperones 2012, 17, 589–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, D.; Shi, Z.; Zhang, S.; Cao, Z.; Liu, W.; Li, L. Extra small virus-like particles (XSV) and nodavirus associated with whitish muscle disease in the giant freshwater prawn, Macrobrachium rosenbergii. J. Fish Dis. 2003, 26, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Clayton, M.E.; Steinmann, R.; Fent, K. Different expression patterns of heat shock proteins hsp 60 and hsp 70 in zebra mussels (Dreissena polymorpha) exposed to copper and tributyltin. Aquat. Toxicol. 2000, 47, 213–226. [Google Scholar] [CrossRef]
- Karouna-Renier, N.K.; Zehr, J.P. Short-term exposures to chronically toxic copperconcentrations induce HSP70 proteins in midge larvae (Chironomus tentans). Sci. Total Environ. 2003, 312, 267–272. [Google Scholar] [CrossRef]
- Li, F.H.; Luan, W.; Zhang, C.S.; Zhang, J.Q.; Wang, B.; Xie, Y.S. Cloning of cytoplasmic heat shock protein 90 (FcHSP90) from Fenneropenaeus chinensis and its expression response to heat shock and hypoxia. Cell Stress Chaperones 2009, 14, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wang, W.N.; He, W.Y.; Zheng, Y.; Wang, L.; Xin, Y.; Liu, Y.; Wang, A.L. Expression of HSP60 and HSP70 in white shrimp, Litopenaeus vannamei in response to bacterial challenge. J. Invertebr. Pathol. 2010, 103, 170–178. [Google Scholar] [CrossRef]
- Park, K.; Kwak, I.S. Expression of stress response HSP70 gene in Asian paddle crabs, Charybdis japonica, exposure to endocrine disrupting chemicals, bisphenol A (BPA) and 4-nonylphenol (NP). Ocean Sci. J. 2013, 48, 207–214. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene Name | Accession Number | RHOD Superfamily Domain Length | Identity (%) | Similarity (%) | Gap (%) |
|---|---|---|---|---|---|---|
| Macrophthalmus japonicus | Heat Shock protein 67B2 | 98 | ||||
| Penaeus vannamei | Heat Shock protein 67B2 | ROT83326.1 | 103 | 72.8 | 81.6 | 4.9 |
| Lepeophtheirus salmonis | Heat Shock protein 67B2 | ACO11957.1 | 106 | 43.4 | 60.4 | 7.5 |
| Lucilia cuprina | Heat Shock protein 67B2-like | XP_023305341.1 | 101 | 40.0 | 60.0 | 10.5 |
| Hyalella azteca | PREDICTED: heat shock protein 67B2-like | XP_018026303.1 | 106 | 44.3 | 58.5 | 7.5 |
| Drosophila busckii | PREDICTED: heat shock protein 67B2 | XP_017840691.1 | 99 | 39.8 | 58.3 | 8.7 |
| Anopheles darlingi | Heat Shock protein 67B2 | ETN62322.1 | 103 | 39.8 | 56.3 | 4.9 |
| Aedes aegypti | Heat Shock protein 67B2 isoform X3 | XP_021696792.1 | 99 | 35.9 | 54.4 | 8.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, K.; Kim, W.-S.; Kwak, I.-S. Effects of di-(2-ethylhexyl) phthalate on Transcriptional Expression of Cellular Protection-Related HSP60 and HSP67B2 Genes in the Mud Crab Macrophthalmus japonicus. Appl. Sci. 2020, 10, 2766. https://0-doi-org.brum.beds.ac.uk/10.3390/app10082766
Park K, Kim W-S, Kwak I-S. Effects of di-(2-ethylhexyl) phthalate on Transcriptional Expression of Cellular Protection-Related HSP60 and HSP67B2 Genes in the Mud Crab Macrophthalmus japonicus. Applied Sciences. 2020; 10(8):2766. https://0-doi-org.brum.beds.ac.uk/10.3390/app10082766
Chicago/Turabian StylePark, Kiyun, Won-Seok Kim, and Ihn-Sil Kwak. 2020. "Effects of di-(2-ethylhexyl) phthalate on Transcriptional Expression of Cellular Protection-Related HSP60 and HSP67B2 Genes in the Mud Crab Macrophthalmus japonicus" Applied Sciences 10, no. 8: 2766. https://0-doi-org.brum.beds.ac.uk/10.3390/app10082766