
Characterization and Biotechnological Potential of Two Native Marine Microalgae Isolated from the Tunisian Coast

 , ,
, ,  and
and
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Axenic Uniclonal Cultures
2.2. DNA Extraction, PCR Amplification, Sequencing and Phylogenetic Analysis
2.3. Pigments Contents
2.4. Proteins Contents
2.5. Carbohydrates Contents
2.6. Lipids Contents and Fatty Acid Profiles
2.7. Determination of Phenols Contents
2.8. Crude Extracts Preparation
2.9. DPPH Free-Radical Scavenging Assay
2.10. ABTS Radical Scavenging Assay
2.11. Antioxidant Enzyme Assays
2.12. Antibacterial Activities
2.13. Cytotoxicity and Antitumor Assay
2.14. Cadmium Removal Capacity
2.15. Statistical Analysis
3. Results
3.1. Isolation and Identification of the Microalgae Strains
3.2. Biochemical Composition
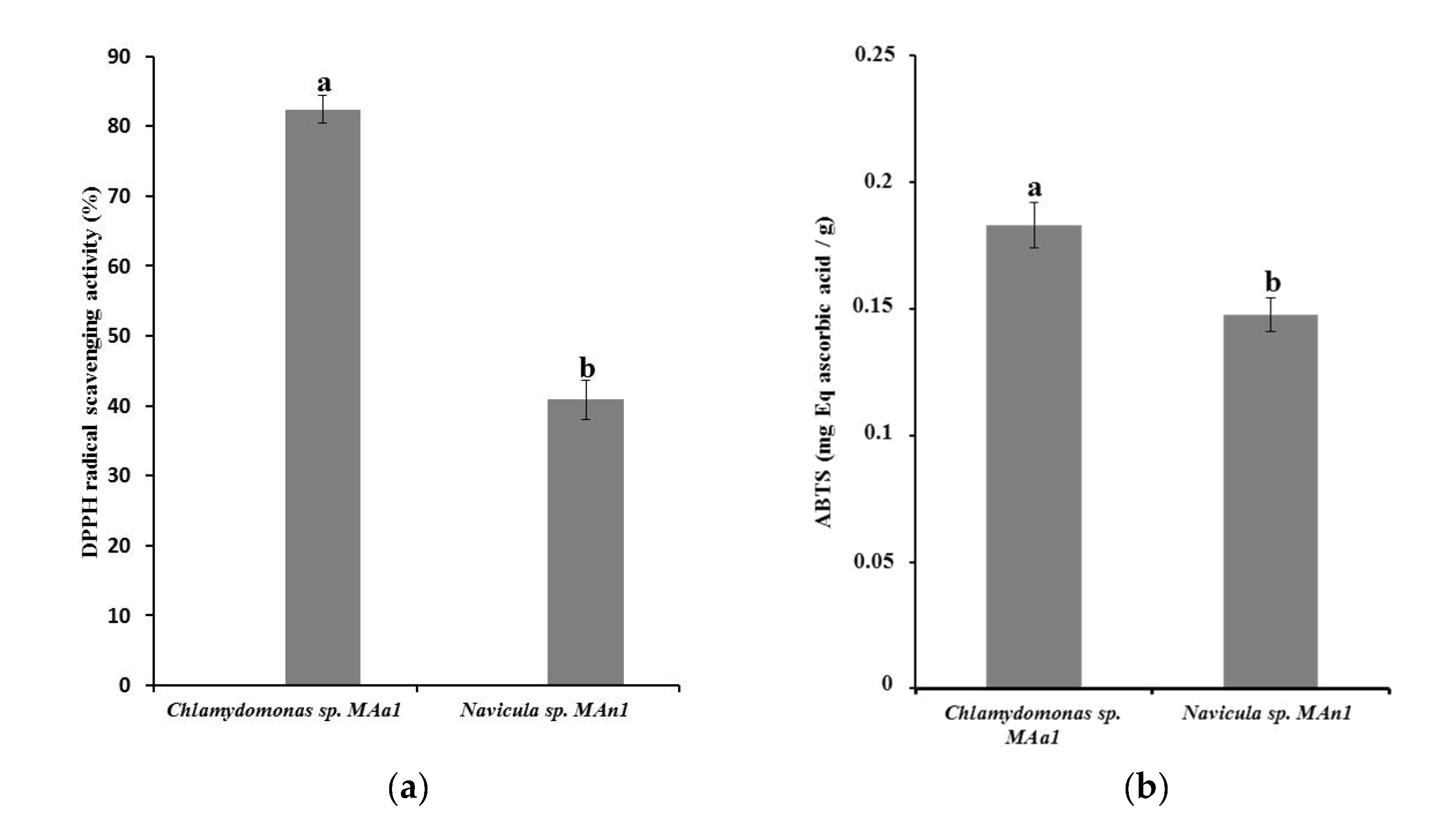
3.3. Antioxidant Activity
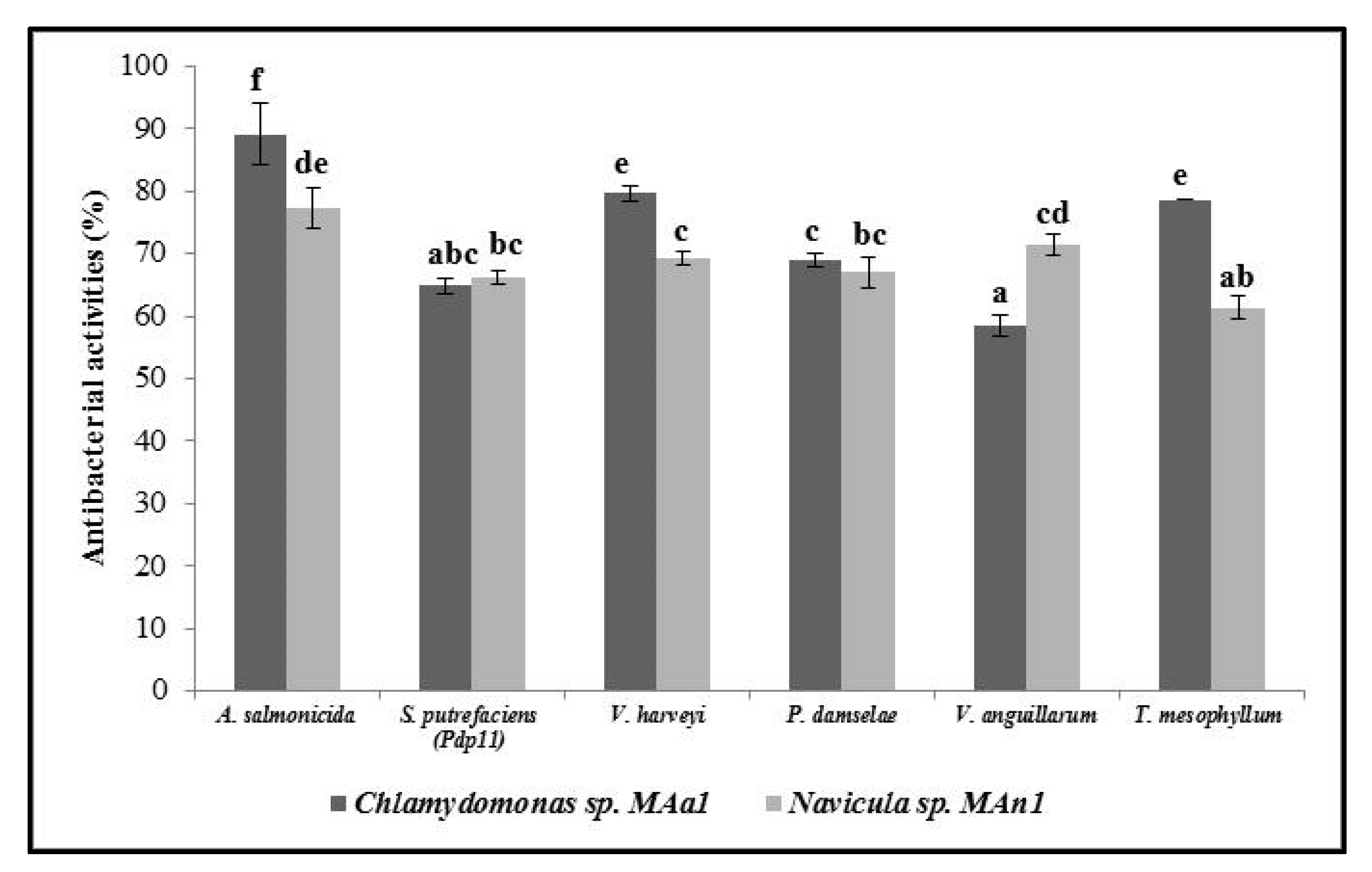
3.4. Antibacterial Activity
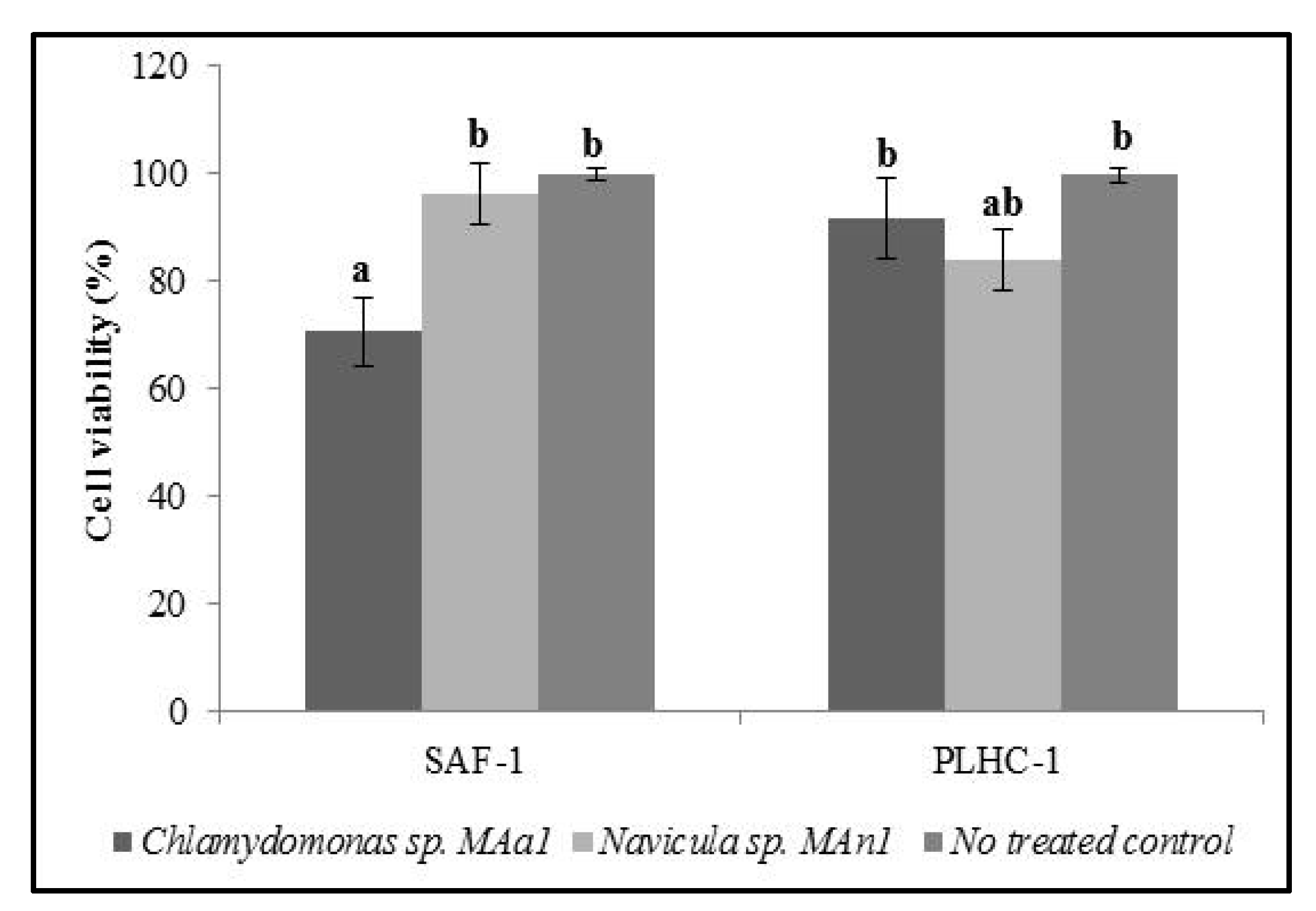
3.5. Cytotoxic Assay
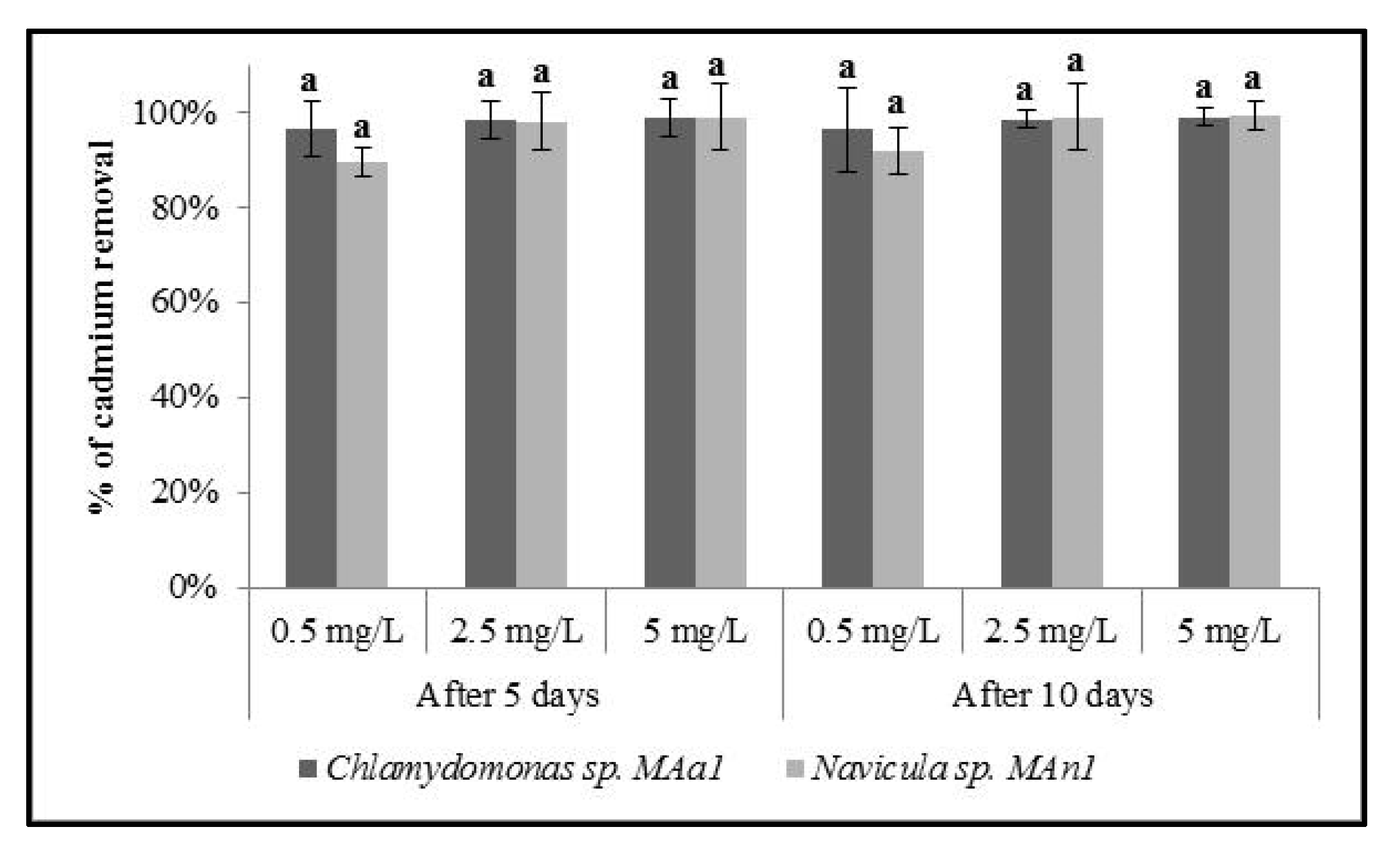
3.6. Capacity of Microalgae to Remove Heavy Metals
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Groendahl, S.; Kahlert, M.; Fink, P. The best of both worlds: A combined approach for analysing microalgal diversity via metabar coding and morphology-based methods. PLoS ONE 2017, 12, e0172808. [Google Scholar] [CrossRef] [PubMed]
- Dammak, M.; Haase, S.M.; Miladi, R.; Ben Amor, F.; Barkallah, M.; Gosset, D.; Pichon, C.; Huchzermeyer, B.; Fendri, I.; Denis, M.; et al. Enhanced lipid and biomass production by a newly isolated and identified marine microalga. Lipids Health Dis. 2016, 15, 209. [Google Scholar] [CrossRef] [Green Version]
- Nehmé, R.; Atieh, C.; Fayad, S.; Claude, B.; Chartier, A.; Tannoury, M.; Elleuch, F.; Abdelkafi, S.; Pichon, C.; Morin, P. Microalgae amino acid extraction and analysis at nanomolar level using electroporation and capillary electrophoresis with laser-induced fluorescence detection. J. Sep. Sci. 2016, 40, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Ben Hlima, H.; Bohli, T.; Kraiem, M.; Ouederni, A.; Mellouli, L.; Michaud, P.; Abdelkafi, S.; Smaoui, S. Combined effect of Spirulina platensis and Punica granatum peel extacts: Phytochemical content and antiphytophatogenic activity. Appl. Sci. 2019, 9, 5475. [Google Scholar] [CrossRef] [Green Version]
- Ben Hlima, H.; Dammak, M.; Karkouch, N.; Hentati, F.; Laroche, C.; Michaud, P.; Fendri, I.; Abdelkafi, S. Optimal cultivation towards enhanced biomass and floridean starch production by Porphyridium marinum. Int. J. Biol. Macromol. 2019, 129, 152–161. [Google Scholar] [CrossRef]
- Elleuch, F.; Baril, P.; Barkallah, M.; Perche, F.; Abdelkafi, S.; Fendri, I.; Pichon, C. Deciphering the biological activities of Dunaliella sp. aqueous extract from stressed conditions on breast cancer: From in vitro to in vivo investigations. Int. J. Mol. Sci. 2020, 21, 1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elleuch, J.; Hadj Kacem, F.; Amor, F.B.; Hadrich, B.; Michaud, P.; Fendri, I.; Abdelkafi, S. Extracellular neutral protease from Arthrospira platensis: Production, optimization and partial characterization. Int. J. Biol. Macromol. 2021, 167, 1491–1498. [Google Scholar] [CrossRef]
- Yasin, D.; Zafaryab, M.; Ansari, S.; Ahmad, N.; Khana, N.F.; Zaki, A.; Rizvi, M.M.A.; Fatma, T. Evaluation of antioxidant and anti-proliferative efficacy of Nostoc muscorum NCCU-442. Biocatal. Agric. Biotechnol. 2019, 17, 284–293. [Google Scholar] [CrossRef]
- Santos, D.S.; Lauria, P.S.S.; Evangelista, A.F.; Azeredo, F.J.; Costa, J.A.V.; Soares, M.B.P.; Druzian, J.I.; Villarreal, C.F. Beyond inflammation: Centrally mediated antinociceptive properties of Spirulina platensis LEB-18 biomass via the opioid system. J. Funct. Foods 2020, 72, 104083. [Google Scholar] [CrossRef]
- Barkallah, M.; Ben Slima, A.; Elleuch, F.; Fendri, I.; Pichon, C.; Abdelkafi, S.; Baril, P. Protective role of Spirulina platensis against bifenthrin-induced reprotoxicity in adult male mice by reversing expression of altered histological, biochemical, and molecular markers including microRNAs. Biomolecules 2020, 10, 7539. [Google Scholar] [CrossRef] [PubMed]
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a future food source. Biotechnol. Adv. 2020, 41, 107536. [Google Scholar] [CrossRef]
- Bošnjaković, M.; Sinaga, N. The perspective of large-scale production of algae biodiesel. Appl. Sci. 2020, 10, 8181. [Google Scholar] [CrossRef]
- Fattah, I.M.R.; Noraini, M.Y.; Mofijur, M.; Silitonga, A.S.; Badruddin, I.A.; Khan, T.M.Y.; Ong, H.C.; Mahlia, T.M.I. Lipid extraction maximization and enzymatic synthesis of biodiesel from microalgae. App. Sci. 2020, 10, 6103. [Google Scholar] [CrossRef]
- Gupta, S.; Pawar, S.B.; Pandey, R.A. Current practices and challenges in using microalgae for treatment of nutrient rich wastewater from agro-based industries. Sci. Total Environ. 2019, 687, 1107–1126. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, M.; Vecchi, V.; Barera, S.; Dall’Osto, L. Biomass from microalgae: The potential of domestication towards sustainable biofactories. Microb. Cell Factories 2018, 17, 173. [Google Scholar] [CrossRef] [Green Version]
- Elleuch, J.; Barkallah, M.; Smith, K.F.; Ben Neila, I.; Fendri, I.; Abdelkafi, S. PCR assay for the simultaneous identification and enumeration of multiple Karenia species. Environ. Sci. Poll. Res. 2020, 27, 36889–36899. [Google Scholar] [CrossRef]
- Ben Amor, F.; Elleuch, F.; Ben Hlima, H.; Garnier, M.; Saint-Jean, B.; Barkallah, M.; Pichon, C.; Abdelkafi, S.; Fendri, I. Proteomic analysis of the chlorophyta Dunaliella new strain AL-1 revealed global changes of metabolism during high carotenoid production. Mar. Drugs 2017, 15, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dammak, M.; Hadrich, B.; Barkallah, M.; Hentati, F.; Ben Hlima, H.; Pichon, C.; Denis, M.; Fendri, I.; Michaud, P.; Abdelkafi, S. Modelling Tetraselmis sp. growth-kinetics and optimizing bioactive-compound production through environmental conditions. Bioresour. Technol. 2018, 249, 510–518. [Google Scholar] [CrossRef]
- Wittkopp, T.M. Isolation of genomic DNA from Chlamydomonas reinhardtii. Bio-Protocol 2018, e2838. [Google Scholar] [CrossRef]
- Chtourou, H.; Dahmen, I.; Hassairi, I.; Abdelkafi, S.; Sayadi, S.; Dhouib, A. Dunaliella sp. a wild algal strain isolated from the Sfax-Tunisia solar evaporating salt-ponds, a high potential for biofuel production purposes. J. Biobased Mater. Bioenergy 2014, 8, 27–34. [Google Scholar] [CrossRef]
- Ben Amor, F.; Barkallah, M.; Elleuch, F.; Karkouch, N.; Dammak, M.; Baréa, B.; Villeneuve, P.; Abdelkafi, S.; Fendri, I. Cyanobacteria as source of marine bioactive compounds: Molecular specific detection based on Δ9 desaturase gene. Int. J. Biol. Macromol. 2017, 105, 1440–1445. [Google Scholar] [CrossRef]
- Fazeli, M.R.; Tofighi, H.; Samadi, N.; Jamalifar, H.; Fazeli, A. Carotenoids ccumulation by Dunaliella tertiolecta (Lake Urmia isolate) and Dunaliella salina (CCAP 19⁄18 & WT) under stress conditions. DARU J. Pharm. Sci. 2006, 14, 146–150. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Barbarino, E.; Lourenço, S.O. An evaluation of methods for extraction and quantification of protein from marine macro-and microalgae. J. Appl. Phycol. 2005, 17, 447–460. [Google Scholar] [CrossRef]
- Elleuch, J.; Jaoua, S.; Ginibre, C.; Chandre, F.; Tounsi, S.; Zghal, R.Z. Toxin stability improvement and toxicity increase against dipteran and lepidopteran larvae of Bacillus thuringiensis crystal protein Cry2Aa Pest. Manag. Sci. 2016, 72, 2240–2246. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Fendri, I.; Chaari, A.; Dhouib, A.; Jlassi, B.; Abousalham, A.; Carrière, F.; Sayadi, S.; Abdelkafi, S. Isolation, identification and characterization of a new lipolytic Pseudomonas sp., strain AHD-1, from Tunisian soil. Environ. Technol. 2010, 31, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidant by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Abdelkafi, S.; Labat, M.; Ben Ali Gam, Z.; Lorquin, J.; Sayadi, S. Optimized conditions for the synthesis of vanillic acid under hypersaline conditions by Halomonas elongata DSM 2581T resting cells. World J. Microbiol. Biotechnol. 2018, 24, 675–680. [Google Scholar] [CrossRef]
- Yan, X.; Nagata, T.; Fan, X. Antioxidative activities in some common seaweeds. Plant Foods Hum. Nutr. 1998, 52, 253–262. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. Methods to measure the antioxidant activity in plant material. A comparative discussion. Free Radic. Res. 1999, 31, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Scott, S.L.; Chen, W.J.; Bakac, A.; Espenson, J.H. Spectroscopic parameters, electrode potentials, acid ionisation constants and electron exchange rates of the 2,2′-azinobis(3-ethylbenzothiazoline-6-sulfonate) radicals and ions. J. Phys. Chem. 1993, 97, 6710–6714. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods. Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Singh, S.; Verma, A.; Dubey, V.K. Effectivity of anti-oxidative enzymatic system on diminishing the oxidative stress induced by aluminium in chickpea (Cicer arietinum L.) seedlings. Braz. J. Plant Physiol. 2012, 24, 47–54. [Google Scholar] [CrossRef]
- Ben Hsouna, A.; Ben Halima, N.; Abdelkafi, S.; Hamdi, N. Essential oil from Artemisia phaeolepis: Chemical composition and antimicrobial activities. J. Oleo Sci. 2013, 62, 973–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.G.; Kehrli, M.E., Jr.; Canning, P.C. A colorimetric assay for quantitating bovine neutrophil bactericidal activity. Vet. Immunol. Immunopathol. 1991, 28, 45–56. [Google Scholar] [CrossRef]
- Espinosa Ruiz, C.; Manuguerra, S.; Cuesta, A.; Santulli, A.; Messina, C.M. Oxidative stress, induced by sub-lethal doses of BDE 209, promotes energy management and cell cycle modulation in the marine fish cell line SAF-1. Int. J. Environ. Res. Public Health 2019, 16, 474. [Google Scholar] [CrossRef] [Green Version]
- Travieso, L.; Canizares, R.O.; Borja, R.; Benitez, F.; Dominguez, A.R.; Valiente, V. Heavy metal removal by microalgae. Bull. Environ. Contam. Toxicol. 1999, 62, 144–151. [Google Scholar] [CrossRef]
- Pradhan, J.; Das, S.; Das, B.K. Antibacterial activity of freshwater microalgae: A review. Afr. J. Pharm. Pharmacol. 2014, 8, 809–818. [Google Scholar] [CrossRef]
- Kroumov, A.D.; Scheufele, F.B.; Trigueros, D.E.G.; Modenes, A.N.; Zaharieva, M.; Najdenski, H. Algal Green Chemistry; Elsevier: Amsterdam, The Netherlands, 2017; pp. 201–241. [Google Scholar]
- Beszteri, B.; Acs, E.; Makk, J.; Kovács, G.; Márialigeti, K.; Kiss, K.T. Phylogeny of six naviculoid diatoms based on 18S rDNA sequences. Int. J. Syst. Evol. Microbiol. 2001, 51, 1581–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson, J.; Toth, G.B.; Albers, E. Biochemical composition of red, green and brown seaweeds on the Swedish west coast. J. Appl. Phycol. 2020, 32, 3305–3317. [Google Scholar] [CrossRef]
- Behl, K.; SeshaCharan, P.; Joshi, M.; Sharma, M.; Mathur, A.; Kareya, M.S.; Jutur, P.P.; Bhatnagar, A.; Nigam, S. Multifaceted applications of isolated microalgae Chlamydomonas sp. TRC-1 in wastewater remediation, lipid production and bioelectricity generation. Bioresour. Technol. 2020, 304, 122993. [Google Scholar] [CrossRef]
- Fimbres-Olivarria, D.; López-Elias, J.A.; Martínez-Córdova, L.R.; Carvajal-Millan, E.; Enriquez-Ocana, F.; Valdéz-Holguin, E.; Miranda-Baeza, A. Growth and biochemical composition of Navicula sp. cultivated at two light intensities and three wavelengths. Isr. J. Aquac. 2015, 67, 1155. [Google Scholar] [CrossRef]
- Brown, M.; Jeffrey, S.; Volkman, J.; Dunstan, G. Nutritional properties of microalgae for mariculture. Aquaculture 1997, 151, 315–331. [Google Scholar] [CrossRef]
- Yang, H.-L.; Lu, C.-K.; Chen, S.-F.; Chen, Y.-M.; Chen, Y.-M. Isolation and characterization of taiwanese heterotrophic microalgae: Screening of strains for docosahexaenoic acid (DHA) production. Mar. Biotechnol. 2010, 12, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Mai, K.; Sun, S. Total lipid and fatty acid composition of eight strains of marine diatoms. Chin. J. Oceanol. Limnol. 2000, 18, 345–349. [Google Scholar] [CrossRef]
- Balic, A.; Vlasic, D.; Zuzul, K.; Marinovic, B.; Mokos, Z.B. Omega-3 versus Omega-6 polyunsaturated fatty acids in the prevention and treatment of inflammatory skin diseases. Int. J. Mol. Sci. 2020, 21, 741. [Google Scholar] [CrossRef] [Green Version]
- Matsukawa, A.; Hogaboam, C.M.; Lukacs, N.W.; Lincoln, P.M.; Strieter, R.M.; Kunkel, S.L. Endogenous MCP-1 influences systemic cytokine balance in a murine model of acute septic peritonitis. Exp. Mol. Pathol. 2000, 68, 77–84. [Google Scholar] [CrossRef]
- Jaime, L.; Mendiola, J.A.; Herrero, M.; Soler-Rivas, C.; Santoyo, S.; Senorans, F.; Cifuentes, A. Ibanez, E. Separation and characterization of antioxidants from Spirulina platensis microalga combining pressurized liquid extraction, TLC, and HPLC-DAD. J. Sep. Sci. 2005, 28, 2111–2119. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-B.; Cheng, K.-W.; Wong, C.-C.; Fan, K.-W.; Chen, F.; Jiang, Y. Evaluation of antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food. Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- Wu, L.; Ho, J.A.; Shieh, M.-C.; Lu, I.-W. Antioxidant and antiproliferative activities of Spirulina and Chlorella water extracts. J. Agric. Food Chem. 2005, 53, 4207–4212. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.; Jaime, L.; Martín-Álvarez, P.J.; Cifuentes, A.; Ibañez, E. Optimization of the extraction of antioxidants from Dunaliella salina microalga by pressurized liquids. J. Agric. Food Chem. 2006, 54, 5597–5603. [Google Scholar] [CrossRef]
- Rao, A.R.; Sarada, R.; Baskaran, V.; Ravishankar, G.A. Antioxidant activity of Botryococcus braunii extract elucidated in vitro models. J. Agric. Food. Chem. 2006, 54, 4593–4599. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, P.; Manimaran, K.; Sampathkumar, P.; Jaikumar, M.; Robin, R.S.; Saravanakumar, C.; Sureshkuma, C. In vitro antioxidant activity of marine diatoms. OSR J. Environ. Sci. Toxicol. Food. Technol. 2013, 5, 32–37. [Google Scholar] [CrossRef]
- Jimenez-Escrig, A.; Jimenez-Jimenez, I.; Pulido, R.; Saura-Calixto, F. Antioxidant activity of fresh and processed edible seaweeds. J. Sci. Food Agric. 2001, 81, 530–534. [Google Scholar] [CrossRef]
- Safafar, H.; van Wagenen, J.; Moller, P.; Jacobsen, C. Carotenoids, phenoliccompounds and tocopherols contribute to the antioxidative properties of some microalgae species grown on industrial wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef] [Green Version]
- Dineshkumar, R.; Narendran, R.; Jayasingam, P.; Sampathkumar, P. Cultivation and chemical composition of microalgae Chlorella vulgaris and its antibacterial activity against human pathogens. J. Aquac. Mar. Biol. 2017, 5, 00119. [Google Scholar] [CrossRef]
- Little, S.M.; Senhorinho, G.N.A.; Saleh, M.; Basiliko, N.; Scott, J.A. Antibacterial compounds in green microalgae from extremeenvironments: A review. Algae 2021, 36, 61–72. [Google Scholar] [CrossRef]
- Costa, M.; Costa-Rodrigues, J.; Fernandes, M.H.; Barros, P.; Vasconcelos, V.; Martins, R. Marine cyanobacteria compounds with anticancer properties: A review on the implication of apoptosis. Mar. Drugs 2012, 10, 2181–2207. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Ratha, S.K.; Sood, A.; Chaudhary, V.; Prasanna, R. New insights into the biodiversity and applications of cyanobacteria (blue-green algae): Prospects and challenges. Algal. Res. 2013, 2, 79–97. [Google Scholar] [CrossRef]
- Ward, O.P.; Singh, A. Omega-3/6 fatty acids: Alternative sources of production. Process. Biochem. 2005, 40, 3627–3652. [Google Scholar] [CrossRef]
- Smith, V.J.; Desbois, A.P.; Dyrynda, E.A. Conventional and unconventional antimicrobials from fish, marine invertebrates and micro-algae. Mar. Drugs 2010, 8, 1213–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okunowo, W.O.; Oyedeji, A.O.; Ilesanmi, J.A.; Afolabi, L.O.; Umunnakwe, I.E. Antimicrobial, antioxidant potential and chemical composition of the methanolic extracts of Spirogyra setiformis and Navicula species. J. Sci. Res. Dev. 2018, 2, 15–20. [Google Scholar]
- Amaro, H.M.; Guedes, A.C.; Malcata, F.X. Antimicrobial activities of microalgae: An invited review. In Science against Microbial pathogens: Communicating Current Research and Technological Advances; Méndez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2011; pp. 1272–1280. [Google Scholar]
- Darah, I.; Lim, S.H.; Ninthianantham, K. Effects of methanolic extract of Wedelia chinensis Osbeck (Asteraceae) leaves against pathogenic bacteria with emphasize on Bacillus cereus. Indian J. Pharm. Sci. 2013, 75, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Abu-Shanab, B.; Adwan, G.; Abu-Safiya, D.; Adwan, K.; Abu-Shanab, M. Antibacterial activity of Rhus coriaria. L. extracts growing in Palestine. J. Islamic Univ. Gaza 2005, 13, 147–153. [Google Scholar]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.; Romano, G.; Ianora, A. Bioactivity screening of microalgae for antioxidant, anti-inflammatory, anticancer, anti-diabetes, and antibacterial activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Abdillahi, H.S.; Verschaeve, L.; Finnie, J.F.; Van Staden, J. Mutagenicity, antimutagenicity and cytotoxicity evaluation of south African Podocarpus species. J. Ethnopharmacol. 2012, 139, 728–738. [Google Scholar] [CrossRef]
- Cerezuela, R.; Guardiola, F.A.; Meseguer, J.; Esteban, M.A. Enrichment of gilthead seabream (Sparusaurata L. ) diet with microalgae: Effects on the immune system. Fish. Physiol. Biochem. 2012, 38, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- Yadav, G.; Meena, D.K.; Sahoo, A.K.; Das, B.K.; Sen, R. Effective valorization of microalgal biomass for the production of nutritional fish-feed supplements. J. Clean. Prod. 2020, 243, 118697. [Google Scholar] [CrossRef]
- Al-Quraishi, D.O.; Al-Mayaly, I.K. Biosorption of Ccadmium, lead and nickel in their aqueous solution by Nitzschia palea and Navicula incerta. J. Eng. Appl. Sci. 2019, 14, 9114–9120. [Google Scholar]
- Cherifi, O.; Sbihi, K.; Bertrand, M.; Cherifi, K. The removal of metals (Cd, Cu and Zn) from the Tensift river using the diatom Navicula subminuscula Manguin: A laboratory study. Int. J. Adv. Res. Biol. Sci. 2016, 3, 177–187. [Google Scholar] [CrossRef]
- Kumar, K.S.; Dahms, H.U.; Won, E.J.; Lee, J.S.; Shin, K.H. Microalgae—A promising tool for heavy metal remediation. Ecotoxicol. Environ. Saf. 2015, 113, 329–352. [Google Scholar] [CrossRef]
- Mera, R.; Torres, E.; Abalde, J. Sulphate, more than a nutrient, protects the microalga Chlamydomonas moewusii from cadmium toxicity. Aquat. Toxicol. 2014, 148, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Adhiya, J.; Cai, X.; Sayre, R.T.; Traina, S.J. Binding of aqueous cadmium by the lyophilized biomass of Chlamydomonas reinhardtii. Colloid Surf. A-Physicochem. Eng. Asp. 2002, 210, 1–11. [Google Scholar] [CrossRef]
- Tüzün, İ.; Bayramoğlu, G.; Yalçin, E.; Başaran, G.; Çelik, G.; Arica, M.Y. Equilibrium and kinetic studies on biosorption of Hg(II), Cd(II) and Pb(II) ions onto microalgae Chlamydomonas reinhardtii. J. Environ. Manag. 2005, 77, 85–92. [Google Scholar] [CrossRef]
- Maeda, S.; Sakaguchi, T. Introduction to Applied Phycology; Academic Publishing: Amsterdam, The Netherlands, 1990; pp. 109–136. [Google Scholar]
- Rangsayatorn, N.; Upatham, E.S.; Kruatrachue, M.; Pokethitiyook, P.; Lanza, G.R. Phytoremediation potential of Spirulina (Arthrospira) platensis: Biosorption and toxicity studies of cadmium. Environ. Pollut. 2002, 119, 45–53. [Google Scholar] [CrossRef]
- Solisio, C.; Lodi, A.; Soletto, D.; Converti, A. Cadmium biosorption on Spirulina platensis biomass. Bioresour. Technol. 2008, 99, 5933–5937. [Google Scholar] [CrossRef]
- Elleuch, J.; Ben Amor, F.; Chaaben, Z.; Frikha, F.; Michaud, P.; Fendri, I.; Abdelkafi, S. Zinc biosorption by Dunaliella sp. AL-1: Mechanism and effects on cell metabolism. Sci. Total Environ. 2021, 773, 145024. [Google Scholar] [CrossRef]
- Salam, K.A. Towards sustainable development of microalgal biosorption for treating effluents containing heavy metals. Biofuel Res. J. 2019, 6, 948–961. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chlamydomonas sp. MAa1 | Navicula sp. MAn1 | |
|---|---|---|
| Proteins (%) | 47.22 ± 0.19 | 34.16 ± 0.05 |
| Lipids (%) | 25.55 ± 0.2 | 15.00 ± 0.1 |
| Carbohydrates (%) | 27.23 ± 0.05 | 50.83 ± 0.2 |
| Chlorophyll (mg/L) | 1.5 ± 0.3 | 0.95 ± 0.05 |
| Phenols (mg/mL) | 40.5 ± 0.15 | 11.32 ± 0.05 |
| Carotenoid (mg/L) | 3.81 ± 0.3 | 4.72 ± 0.15 |
| Fatty Acid (Relative Percentage) | ||
|---|---|---|
| Chlamydomonas Sp. MAa1 | Navicula Sp. MAn1 | |
| C14:0 Myristic acid | 0.69 | 4.96 |
| C15:0Pentadecylicacid | 0.13 | 1.62 |
| C16:0 Palmitic acid | 23.87 | 10.80 |
| C18:0 Stearic acid | 4.98 | 0.45 |
| C20:0 Arachidic acid | 0.27 | 0.00 |
| C22:0 Behenic acid | 0.41 | 0.15 |
| Total saturated (%) | 30.35 | 17.98 |
| C16:1n-7 Palmitoleic acid | 1.14 | 43.48 |
| C18:1n-9 Oleic acid | 12.50 | 5.04 |
| C18:1n-7 Vaccenic acid | 10.45 | 2.07 |
| C20:1n-9 Gondoic acid | 1.00 | 0.14 |
| C22:1n-9 Erucic acid | 0.00 | 0.08 |
| C24:1n-9 Nervonic acid | 0.21 | 0.07 |
| Total monounsaturated (%) | 25.30 | 50.88 |
| C18:2n-6 Linoleic acid | 5.82 | 2.91 |
| C18:3n-6 γ-linoleic acid | 0.33 | 1.46 |
| C20:3n-6 Dihomo-γ-linoleic acid | 0.00 | 0.56 |
| C20:4n-6 Arachidonic acid | 1.41 | 5.64 |
| C22:2n-6 Docosadonic acid | 0.36 | 0.14 |
| C22:4n-6 Adrenic acid | 0.00 | 0.14 |
| Total n-6 PUFA (%) | 7.92 | 10.85 |
| C18:3n-3 α-linolenic acid | 30.08 | 0.00 |
| C18:4n-3 Stearidonic acid | 0.00 | 0.05 |
| C20:3n-3 Dihomo-α-linolenic acid | 0.20 | 0.00 |
| C20:5n-3 Eicosapentaenoic acid | 6.15 | 19.99 |
| C22:6n-3 Docosahexanoic acid | 0.00 | 0.25 |
| Total n-3 PUFA (%) | 36.43 | 20.29 |
| Total PUFAs (%) | 44.35 | 31.14 |
| n-3/n-6 | 4.59 | 1.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, J.B.; Elleuch, J.; Drira, M.; Esteban, M.Á.; Michaud, P.; Abdelkafi, S.; Fendri, I. Characterization and Biotechnological Potential of Two Native Marine Microalgae Isolated from the Tunisian Coast. Appl. Sci. 2021, 11, 5295. https://0-doi-org.brum.beds.ac.uk/10.3390/app11115295
Mohamed JB, Elleuch J, Drira M, Esteban MÁ, Michaud P, Abdelkafi S, Fendri I. Characterization and Biotechnological Potential of Two Native Marine Microalgae Isolated from the Tunisian Coast. Applied Sciences. 2021; 11(11):5295. https://0-doi-org.brum.beds.ac.uk/10.3390/app11115295
Chicago/Turabian StyleMohamed, Jamila Ben, Jihen Elleuch, Marwa Drira, Maria Ángeles Esteban, Philippe Michaud, Slim Abdelkafi, and Imen Fendri. 2021. "Characterization and Biotechnological Potential of Two Native Marine Microalgae Isolated from the Tunisian Coast" Applied Sciences 11, no. 11: 5295. https://0-doi-org.brum.beds.ac.uk/10.3390/app11115295