
Micro-Landscape Dependent Changes in Arbuscular Mycorrhizal Fungal Community Structure

1
Institut für Biologie, Freie Universität Berlin, Altensteinstr. 6, D-14195 Berlin, Germany
2
Berlin-Brandenburg Institute of Advanced Biodiversity Research, D-14195 Berlin, Germany
*
Author to whom correspondence should be addressed.
†
These two authors contributed equally.
Appl. Sci. 2021, 11(11), 5297; https://0-doi-org.brum.beds.ac.uk/10.3390/app11115297
Submission received: 20 April 2021
/
Revised: 31 May 2021
/
Accepted: 1 June 2021
/
Published: 7 June 2021
(This article belongs to the Special Issue Hide and Seek of Soil Microbes—Who Is Where with Whom and Why?)
{kind=link}
{kind=link}
{kind=link}
Abstract
:The roots of most plants host diverse assemblages of arbuscular mycorrhizal fungi (AMF), which benefit the plant hosts in diverse ways. Even though we understand that such AMF assemblages are non-random, we do not fully appreciate whether and how environmental settings can make them more or less predictable in time and space. Here we present results from three controlled experiments, where we manipulated two environmental parameters, habitat connectance and habitat quality, to address the degree to which plant roots in archipelagos of high connectance and invariable habitats are colonized with (i) less diverse and (ii) easier to predict AMF assemblages. We observed no differences in diversity across our manipulations. We show, however, that mixing habitats and varying connectance render AMF assemblages less predictable, which we could only detect within and not between our experimental units. We also demonstrate that none of our manipulations favoured any specific AMF taxa. We present here evidence that the community structure of AMF is less responsive to spatio-temporal manipulations than root colonization rates which is a facet of the symbiosis which we currently poorly understand.
1. Introduction
Arbuscular mycorrhizal (AM) associations form direct nutritional symbioses between the roots of most terrestrial plants and a monophyletic group of soil-borne fungi belonging to the phylum Glomeromycota [1]. AM associations have attracted a lot of attention because they can promote net primary productivity (NPP) and agricultural production [2,3]. NPP gains can partially determine how AM fungal communities in plant roots are structured [4,5,6]. As a result, a lot of the literature addresses practices that likely select for more beneficial communities of Glomeromycota in plant roots (e.g., [7,8]) and environmental parameters and practices that determine AM fungal community structure (e.g., [7,9,10]). An alternative way to ask this question is via questioning how AM fungal diversity varies in space and time (i.e., which entails addressing the fraction of variance which is often classified in models as “unexplained”; [11,12]).
Our general understanding so far is that AM fungal assemblages in the roots are non-random. This has been shown both in relation to null-model analyses [13,14], which assess the degree to which chance exclusively could have generated the observed community table (i.e., the occurrences of AM fungal species across root samples) of the study, and models exploring species-abundance distributions [15,16], which essentially test whether particular groups of species have been more abundant than expected by chance. Many studies observing preferential establishment of AMF taxa in specific habitats also hint towards this direction (e.g., [9,17]). Specific biotic and abiotic parameters of the habitat (besides exerting selectivity to specific AM fungal taxa), however, might also alter our ability to predict (i.e., modify the predictability of) mycorrhizal community structure in nature, but this point remains underexplored. Two syntheses which addressed this question found that anthropogenic disturbances, environmental heterogeneity and a plant host identity (i.e., being a monocotyledon) render AM fungal communities less predictable (i.e., more divergent) than they would have been expected to be by chance alone [18,19]. More recently, Deveautour et al. [20] assayed AM fungal communities in the field to determine the degree to which AM fungal communities diverge with spatial distance but also when sampling from the root systems of the same or from a different plant-host individual. Deveautour et al. [20] observed small differences in AMF community turnover between adjacent neighbouring plants (as compared to sampling from the same individual) but also that AMF community turnover increased for plant individuals further away from each other.
A particular feature of AM fungi is that they are obligate symbionts, meaning that they cannot fulfil their life cycle in the absence of a suitable host. This limits their ability to colonize soil in some environments because their vegetative growth ceases at distances of about 50 cm from the closest colonized root [21]. There is a large body of literature addressing how dispersal constraints modify the community structure of organisms addressing variable types of landscape which can also occur at a micro level such as in soil in which case we can refer to them as micro-landscapes or meta-communities. There is a consensus that meta-communities simultaneously reduce local (α-) diversity and increase global (γ-) diversity because they make local community structure less predictable (e.g., [22]) which potentially allows persistence of less competitive species [23]. This point remains underexplored in relation to AM-associations [24]. Here we present a synthesis from three controlled studies with an overall aim to address how spatial structure in plant mesocosms alters predictability in AM fungal communities. Based on the points we made (e.g., [13,15]), we expected that in all experiments AM fungal communities were non-random (Hypothesis One) and that we would observe the highest γ-diversity in those cases in which the connectance of the patches in the archipelago is lowest (Hypothesis Two). Finally, we expected that lowering the connectance of plant and fungal mycorrhizal communities would increase segregation (i.e., the community table becomes more evenly dispersed via weakening pairwise interactions in agreement with the results from Hein et al. [25] showing that strong pairwise interactions promote species aggregation) in Glomeromycota (Hypothesis Three). To the best of our understanding, the point that segregation in AM fungal communities could depend on the structure of the micro-landscape has never been addressed in the past for any fungal group and showcases the high potential (because they have an obligate symbiotic lifestyle and are ubiquitous in nature) of using Glomeromycota as model systems in fungal ecology.
2. Materials and Methods
2.1. Rationale of the Experiments
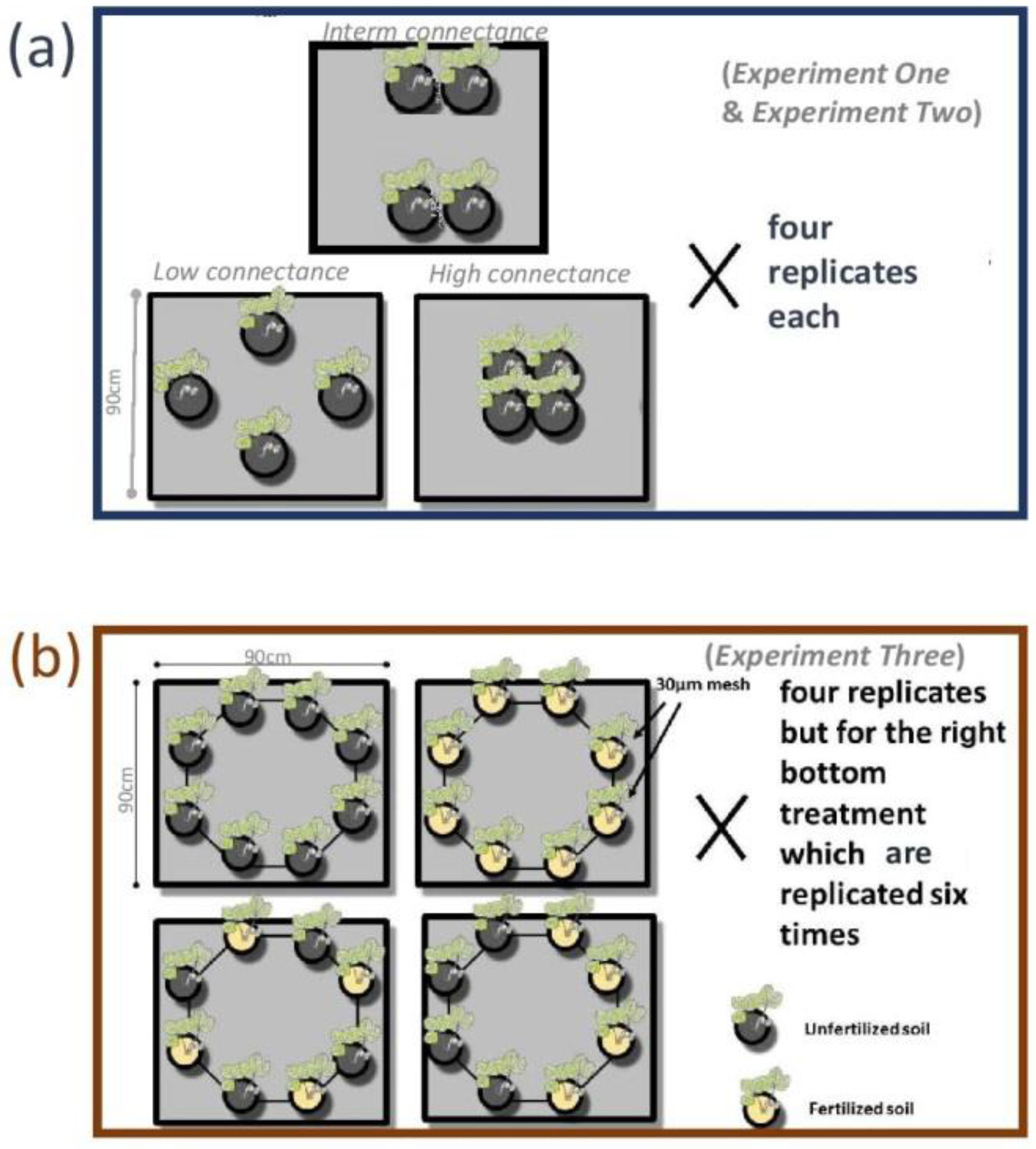
We worked with large mesocosms (i.e., 90 × 90 × 20 cm) as experimental units to which, for consistency with the meta-community literature, we refer to as archipelagos (Figure 1). Within the mesocosms we established patches (i.e., patches in the form of 8 cm diameter × 20 cm height cylindrical inserts containing 30 μm mesh-covered windows to block root growth but allow growth of fungal hyphae) of vegetated habitat and manipulated the connectance of the patches either by means of distances across patches of the “meta-community” (Experiment One and Experiment Two) or the fertility of the patches within each mesocosm (Experiment Three). At the same time via manipulating the distances of the patches we altered the spatial availability of nutrients in the mesocosms and likely also that (i.e., the spatial distribution) of AMF propagules which were contained in those inserts (and were thus influenced by their spatial arrangement). We anticipated that the lack of prospective hosts between inserts (i.e., patches), over distances of up to 70 cm, hindered dispersal of AMF and would induce meta-community dynamics in our experimental units. The idea of using meta-community theory to model symbiotic systems has been developed and explained in larger detail by Mihaljevic [26] (but see Veresoglou et al. [24] for some likely limitations of the approach in the particular case of AMF communities).
2.2. Experimental Work
The experimental work on Experiment Two and Experiment Three has been described in detail in Grünfeld et al. ([27]; the two experiments are described there as Experiment One and Experiment Two, respectively; Figure 1). In brief, we carried out three controlled experiments with rectangular mesocosms sized 90 × 90 × 20 cm (width × length × height; Figure 1). Experiment One and Experiment Two used identical experimental designs consisting of four inserts per mesocosm positioned at different distances (three different levels each replicated four times generating archipelagos of low, intermediate and high connectance) from each other but were carried out with different hosts (Plantago lanceolata and Medicago lupulina; Figure 1a). In Experiment Three we experimented with two different habitats (unfertilized soil and soil fertilized with 1.8 g superphosphate per insert) and the spatial structure of mixtures of them (i.e., aggregated vs. overdispersed spatial structure). In Experiment One some of the P. lanceolata roots penetrated the 30 μm mesh barriers and explored the unvegetated compartment. In Experiment Two and Experiment Three we observed differences in AMF colonization across the treatments which we presented in detail in Grünfeld et al. [27].
The soil that was used for the three experiments was collected from a location in northwest Berlin (52.51° N, 13.14° E), had a pH of 6.7 and contained on average 1.75% organic C and 1.3 g kg−1 N. The freshly collected soil used for the experiments was stored at room temperature for less than two weeks before setting up the experiments. The soil used to fill the patches was unsterilized providing natural microbiota. The soil used to fill the main compartment of the experimental units was mixed 1:1 with sand and steam-sterilized (99 °C for 2 h) in order to destroy AMF propagules. To each of the inserts we added 200–250 seeds (B&T World Seeds, Aigues-Vives, France) of either P. laneolata (Experiment One) or M. lupulina (Experiments Two and Three) to approximate a plant density of one seedling per square cm (e.g., [28]).
In the three experiments we used a fully randomized design. Because of the size of the mesocosms it was impossible to re-randomize the experimental units over the duration of the experiment. The temperature in the air-conditioned glasshouse was maintained close to 20 °C. In all three experiments, two weeks after germination of the seedlings, we set up an automatic irrigation system so that the plants were watered daily (over the first two weeks of the experiments watering was carried out manually). We further controlled growth conditions with five soil moisture sensors (ECH20 EC-5 soil moisture sensors and an Em50 data logger, METERs) positioned in three experimental units: in each experimental unit one of the sensors was in the unvegetated compartment and one in one of the inserts. Watering was adjusted so that soil moisture ranged between 60 and 75 % of the water holding capacity. We inspected plant growth daily and removed any unwanted seedlings.
All three experiments were harvested 12 weeks after sowing, respectively, and cleaned root samples (50 mL core) were frozen at −20 °C before DNA extraction. Plant biomass was dried at 60 °C for three days and weighted. Root material from each insert was used to assess root colonization [29]. Soil cores (five per experimental unit with more details in Grünfeld et al. [27]) were used to assay extraradical hyphae in soil. These results are described in Grünfeld et al. [27].
2.3. Molecular Analyses and Bioinformatics
Roots from each individual insert per experiment were treated as one sample.Root samples were freeze-dried and homogenized with a Retsch Mixer Mill MM 400 and DNA was extracted from 30 mg ground root material per sample with the DNeasy® PowerPlant® Pro Kit (Qiagen). DNA was amplified with a proofreading polymerase (Kapa HiFi; Kapa Biosystems) and the primer pair NS31-AML2 targeting Glomeromycota (Lee et al., 2008). Thermocycling conditions were as follows: Initial denaturation at 95 °C for 2 min, 35 cycles with first 98 °C for 45 s, then 65 °C for 45 s and 72 °C for 45 s and final elongation at 72 °C for 10 min. The PCR master mix for indexing consisted of 1 µL of the purified polymerase chain reaction (PCR) template, 2.4 µL of the primer mix, 0.25 µL polymerase, 0.5 µL dNTPs (10 µM), 5 µL PCR buffer and 15.85 µL nuclease-free water per 25 µL reaction. Amplicons were purified with the NucleoSpin® gel and PCR clean-up kit (Macherey-Nagel, Düren, Germany) and indexed for MiSeq sequencing by means of an additional PCR with the same conditions as described earlier but with only 15 cycles. Amplicons were purified with magnetic beads (GC Biotech, Alphen aan den Rijn, The Netherlands), and were pooled at equimolar quantities. Sequencing was carried out at the Berlin Center for Genomics in Biodiversity Research (BeGenDiv, Berlin, Germany).
Raw sequences were processed with the UPARSE pipeline [30] with USEARCH v 10.0.240 and default settings and were clustered into phylotypes (i.e., Operational Taxonomic Units - OTUs) at a threshold of 97% sequence similarity. Representative OTU sequences were blasted against MaarjAM [31] and non-specific to Glomeromycota (i.e., >97.5 % similarity or >99 % coverage) OTUs were excluded from further analyses. We then rarefied these to 2200 reads which filtered out two samples from further analyses (i.e., analysis was carried out on the remaining 240 samples).
2.4. Statistical Analyses
To address Hypothesis One, stating that AM fungal local communities were non-random, we compared C score (i.e., checkerboards) occurrences in our presence-absence community tables with 1000 randomizations in which we maintained the total number of row sums fixed and the column sums proportional to those of the original community table. This was carried out through the sim4 algorithm (i.e., which is appropriate for assessing incomplete lists) [32] which we implemented through the R package EcoSimR [33]. We presented the results in the form of z-score standardized effect sizes (SES) which can be interpreted as (1) random community structure in the case of scores with absolute SES values below 1.96; (2) aggregation for negative SES values below −1.96; and (3) segregation for positive values above 1.96.
To address Hypothesis Two, stating that low connectance promoted a high γ-diversity in Glomeromycotan communities we used a fixed-effects linear models. We assayed how the experimental design (a categorical predictor with three levels: high connectance vs. intermediate vs. low connectance archipelagos; Figure 1) modified γ-diversity (response variable) in the experimental units. To further gauge the impact of connectance on α- and γ- diversity we calculated those indices (i.e., local to the inserts and global for the entire mesocosm richness estimates, describing essentially the observed in the resulting community table number of OTUs at each of the two hierarchical levels) for individual inserts and modelled them after a repeated-measures analysis of variance (ANOVA) approach in which we used as response variables the diversity indices and the type of meta-community as predictors with additional error terms to model the nesting of inserts within experimental units. To further address the possibility that the treatments induced differences at a community level we implemented redundancy analyses (RDA) with the Hellinger transformed community tables as responses and the treatments as predictors. Additionally, we carried out an indicator species analysis to assess the degree to which phylotypes preferably established in some spatial designs.
To address Hypothesis Three, stating that low connectance of plant communities increases segregation, we used the Jaccard index (i.e., Jaccard similarity coefficient), defined for any pairwise combination of habitats as the ratio of common species over total number of species, as a metric of similarity across communities. We calculated Jaccard similarities for all pairwise combinations of inserts within individual experimental units. To avoid inflating the degrees of freedom we averaged the similarity coefficients describing the similarity of any given insert across habitats of any particular class (i.e., short distance/long distance/(un)fertilized soil patches). To model similarity coefficients we used a repeated-measures ANOVA approach with the Jaccard coefficients as response variable and a structure identical to the models we used to model α- and γ- diversity.
3. Results
3.1. Overall Statistics
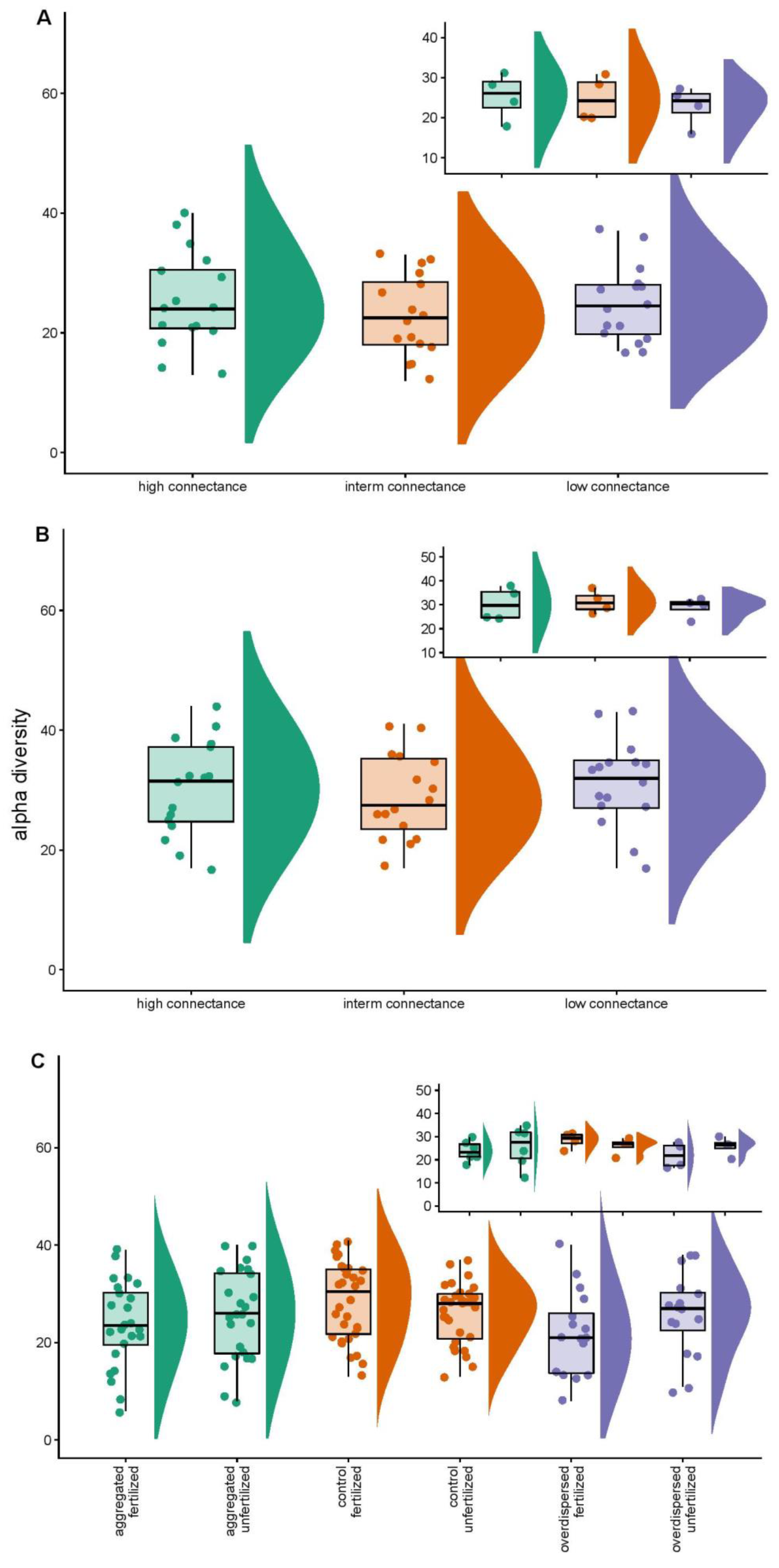
Alpha diversity varied in the experiments between 6 and 44 phylotypes (i.e., 12–40 in Experiment One; 17 to 44 in Experiment Two and 6 to 41 in Experiment Three; Figure 2). Gamma diversity varied between 30 and 53 phylotypes (i.e., 30–48 in Experiment One; 39 to 52 in Experiment Two; 35 to 53 in Experiment Three). In none of the three experiments could we explain alpha (F values varied between 0.38 and 2.1 with respective p values larger than 0.11) or gamma diversity (F values varied between 0.2 and 1.3 with respective p values larger than 0.3) based on the experimental treatments.
Community differences across the treatments were not significant in any of the three experiment specific RDAs (F values varied between 0.96 and 1.22 with adjective R2 values were in all cases below 0.005). Indicator species analysis yielded inconsistent results and a low occurrence frequency of indicators: there were no indicator OTU in Experiment One, there was a single indicator OTU in Experiment Two specific to low connectance archipelagos (with p = 0.03) and there were two OTUs specific to the unfertilized control and one to the fertilized control but not to any of the mixes of them in Experiment Three. Such low frequencies of indicators could have been explained, at least in the case of the two first experiments, by chance.
3.2. Null Model Analyses
All standardized effect size statistics differed from zero and ranged between −9.6 and −20.6 (Experiment One: −9.6; Experiment Two: −10.99 and Experiment Three: −20.66), suggesting community aggregation.
3.3. Comparative Analysis of Jaccard Similarities across Experiments
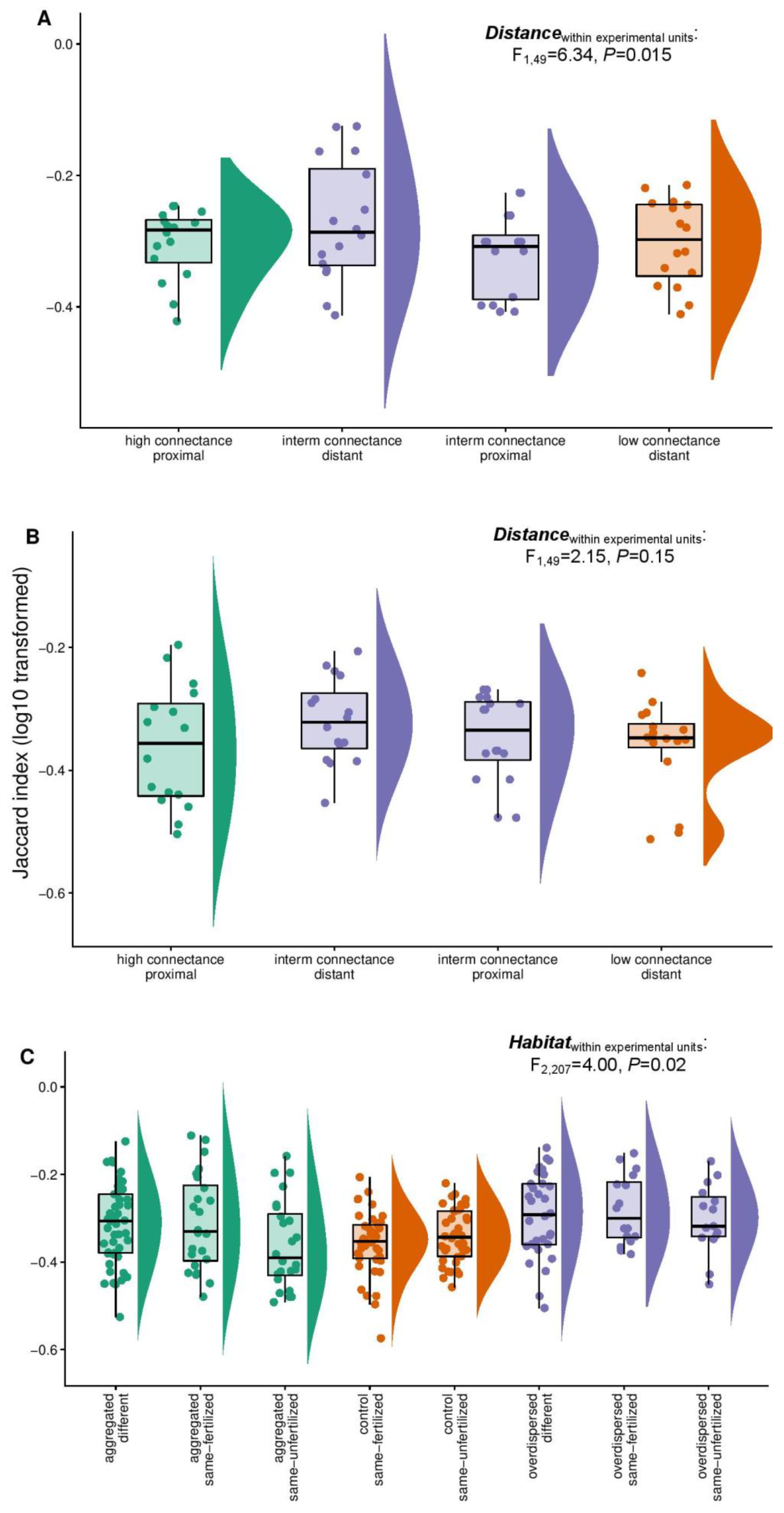
Jaccard similarities did not differ across treatments but within experimental units between short-distance and long-distance inserts in the intermediate connectance treatment of Experiment One (F1, 49 = 6.3, p = 0.015; Figure 3a; Test 1.1 in the supplementary matterials). There was a comparable trend with Jaccard similarities (F1, 49= 2.12, p = 0.15) in Experiment Two (Figure 3b; Test 2.1 in the supplementary materials). In Experiment Three, there were differences in Jaccard similarities only between observations within experimental units which differed in their habitat type (i.e., unfertilized vs. fertilized; unfertilized vs. unfertilized; fertilized vs. fertilized; F2, 207= 4.0, p = 0.02; Figure 3c; Test 3.1 in the supplementary materials). Jaccard similarities were on average larger in the overdispersed treatment compared to the aggregated treatment in Experiment Three (t = −2.03, p = 0.044; Test 3.2 in the supplementary materials).
We thus observed that within experimental units there were differences in AMF community turnover (assayed with the Jaccard index) which peaked for pairs of distantly placed patches (Experiment One and Experiment Two; as compared to closely placed patches) and pairs of patches containing different habitats (Experiment Three). However, we observed no comparable differences between treatments only containing distantly placed vs. only closely placed patches or high fertility vs. low fertility patches (and this is why the predictor treatment was not significant). A high community turnover, in the absence of diversity differences, is evidence of a lower predictability.
4. Discussion
We present evidence from three controlled experiments supporting that small scale micro-landscape heterogeneity (i.e., here describing either experimental units with a low patch-connectance or experimental units containing habitats of different quality) hardly alters diversity patterns in AMF communities. AMF community structure, however, remains non-random. At the same time, we observed that archipelagos combining habitats of both low- and high-connectance (which implies that AMF might have needed to combine traits of long-distance and short-distance dispersal), as well as overdispersed micro-landscapes displayed differences in community turnover (and thus predictability; Test 3.2 in the supplement) across their patches, with pairs of highly connected patches and patches sharing comparable habitats being the most similar to each other. Some conceptual models predict that AMF communities become less random at small (local) spatial scales (i.e., manifested in the form of a low community turnover [34]). With this study we provide experimental evidence that even at such small spatial scales, micro-landscape variability continues to structure AMF communities and can alter their stochasticity (i.e., used here as an opposite to predict).
Our Hypothesis Two stated that we would observe the highest γ-diversity in the cases in which they were most fragmented, but we observed that the differences across treatments in our experiments were unrelated to AMF diversity. Evidence suggests that AMF richness (either in the form of alpha, here defined as OTUs observed per insert, or gamma diversity, here describing the number of OTUs per mesocosm) stays relatively constant across a range of environmental gradients in AMF systems (e.g., [35,36,37]) albeit this is not the case with nutrient availability gradients as has been for example shown in Camenzind et al. [38]. It has actually been proposed that plants impose a strong filter on the number of partners they simultaneously associate with [6,39,40], which could determine AMF richness in plant roots. In our experimental set ups, manipulations of the spatial design altered AMF root colonization [27]. The exact reasons why in mixed micro-landscapes we observed a higher root colonization and variable AMF community turnover (which was masked when comparing across less diverse micro-landscapes) are not clear. We suspect that the underlying mechanism relates to alternative growth strategies across AMF taxa. AMF have been proposed to contain two types of hyphae, absorptive and explorative, which differ in their functions (e.g., [41]). Mixed micro-landscapes might necessitate both types of hyphae to be present at high densities which likely weakens interspecific pairwise co-occurrence interactions across AMF species (i.e., pairs of species found together more frequently than expected by chance and pairs of species co-occurring less frequently than expected by chance; e.g., [42]). High densities of both types of hyphae should theoretically result in a higher diversity of pairwise interactions, including many combinations of short-distance dispersers and long-distance dispersers. Also, mixed micro-landscapes could render the benefits that plants acquire from the different AMF species more variable with long-distance dispersers being favoured in some parts of the micro-landscape whereas short-distance ones in others and thus generate conditions with unclear investment optima. In doing so, mixed microlandscapes favour a more diverse set of AMF [6]. An alternative explanation is that within experimental units we could better control for idiosyncratic parameters that can sometimes determine AMF community structure in the early stages, such as the quality and quantity of the AMF propagules and soil moisture settings throughout the experiment. We think that through controlling those idiosyncratic parameters in our within experimental units comparisons, we might had a higher statistical power to detect differences in community turnover (and thus predictability) than across experimental units.
We found support for Hypothesis One that AM fungal communities were non-random which, however, was not surprising. A large body of the mycorrhizal literature supports the idea as we reviewed in the introduction (e.g., [13,14]). What makes our study novel is that across three controlled experiments we found consistent results on a parameter that determines how random AM fungal communities might be micro-landscape structure. We observed differences in community turnover in mixed micro-landscapes (that were masked in their homogenized counterparts) which was higher for distant patches and patches differing in their habitat quality. This observation aligns well with expectations based on meta-community theory [22]. There have only been a few studies so far quantitatively (i.e., assessing effect sizes on the degree of predictability, rather than simply obtaining a qualitative result such as whether the community is segregated) assessing how predictable synthetic microbial systems can be. A recent meta-analysis on the topic examining 21 datasets showed that organic additions make microbial communities less predictable (i.e., more stochastic; [43]) which was later further supported by an additional study [44]. In another study, Fodelianakis et al. [45] showed that evolutionary drift in synthetic bacterial communities rendered them less predictable than in their original cultures. We used here an important for the functioning of ecosystems, system, arbuscular mycorrhizae, to show that also spatial structure can induce less predictable microbial communities and that this happens when we mix different micro-landscape features.
Arbuscular mycorrhizal fungi are most likely to experience dispersal constraints in urban and agricultural landscapes as well as woody habitats [27]. In the case of agricultural landscapes, the growth settings most likely select for short-distance dispersal traits (i.e., there are uniform distances across crop individuals, which ease the proliferation of AMF species from close by patches of AMF diversity). In contrast, in woody habitats the growth settings most likely select for a combination of long- and short-distance dispersal (i.e., distances between AMF-associating plants most likely vary in time and space). Based on the results of our study, plant hosts in woody habitats could, therefore, experience a higher stochasticity in relation to harbouring AMF community structure than other hosts. This might actually benefit AMF-associating plants in forests, in the longer term. Woody plants, in particular, experience a high mortality at early life-stages. If plant fitness to a certain degree depends on the benefits they acquire from associating with AMF (as we suggest in Veresoglou et al. [46] and Grünfeld et al. [27]), stochasticity in AMF community structure could render plant fitness more variable in both time and space and ensure that the surviving individuals are those that associate with strongly mutualistic AMF (e.g., [47]). Further studying parameters that determine stochasticity in mycorrhizal fungal communities, could be key to explaining why and how plant–soil feedback varies in time and space (e.g., [48,49]).
5. Conclusions
In conclusion we present evidence that mixing micro-habitat features, such as distances across hosts and fertility levels, makes AMF communities more stochastic (i.e., less predictable). This observation presents a range of opportunities to increase AMF diversity (via facilitating establishment of less competitive species) and hopefully productivity in silviculture and agriculture. Glomeromycota, clearly, present a special case of fungi because of their obligate symbiotic lifestyle, meaning that it is possible to control their spatial structure through manipulating the location of their plant hosts. A follow up question revolves around assessing the degree to which there are comparable patterns in other systems of fungi and the overall consequences for ecosystem functioning.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/app11115297/s1, Overview (and detailed statistics) of the six tests that had Jaccard coefficients as response variables.
Author Contributions
Conceived the study, did the statistics/bioinformatics and prepared the first draft of the paper: S.D.V.; Carried out the experimental work and contributed to the writing of the manuscript: L.G.; Carried out the molecular work described in the study: L.G., M.M.; All authors have read and agreed to the published version of the manuscript.
Funding
The authors acknowledge funding from the Deutsche Forschungsgemeinschaft Project Metacorrhiza (VE 736/2-1) awarded to S.D.V.
Institutional Review Statement
The study received no institutional reviews.
Informed Consent Statement
The study used no humans or animals.
Data Availability Statement
The data will be made available in an online repository upon acceptance of the article.
Acknowledgments
We want to thank Matthias Rillig and Maraike Probst for constructive comments. The publication of this article was funded by Freie Universität Berlin.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Hoeksema, J.D.; Chaudhary, V.B.; Gehring, C.A.; Johnson, N.C.; Karst, J.; Koide, R.T.; Pringle, A.; Zabinski, C.; Bever, J.D.; Moore, J.C.; et al. A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecol. Lett. 2010, 13, 394–407. [Google Scholar] [CrossRef]
- Zhang, S.; Lehmann, A.; Zheng, W.; You, Z.; Rillig, M.C. Arbuscular mycorrhizal fungi increase grain yields: A meta-analysis. New Phytol. 2019, 222, 543–555. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Maherali, H.; Klironomos, J.N. Influence of phylogeny on fungal community assembly and ecosystem functioning. Science 2007, 316, 1746–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A.; et al. Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science 2011, 333, 880–883. [Google Scholar] [CrossRef] [Green Version]
- Rillig, M.C.; Sosa-Hernández, M.A.; Roy, J.; Aguilar-Trigueros, C.A.; Vályi, K.; Lehmann, A. Towards an integrated mycorrhizal technology: Harnessing mycorrhiza for sustainable intensification in agriculture. Front. Plant Sci. 2016, 7, 1625. [Google Scholar] [CrossRef] [Green Version]
- Pánková, H.; Lepinay, C.; Rydlová, J.; Voková, A.; Janoušková, M.; Dostálek, T.; Münzbergová, Z. Arbuscular mycorrhizal fungi and associated microbial communities from dry grassland do not improve plant growth on abandoned field soil. Oecologia 2018, 186, 677–689. [Google Scholar] [CrossRef]
- Egerton-Warburton, L.M.; Johnson, N.C.; Allen, E.B. Mycorrhizal community dynamics following nitrogen fertilization: A cross-site test in five grasslands. Ecol. Monogr. 2007, 77, 527–544. [Google Scholar] [CrossRef] [Green Version]
- Veresoglou, S.D.; Caruso, T.; Rillig, M.C. Modelling the environmental and soil factors that shape the niches of two common arbuscular mycorrhizal fungal families. Plant Soil 2013, 368, 507–518. [Google Scholar] [CrossRef]
- Dumbrell, A.J.; Ashton, P.D.; Aziz, N.; Feng, G.; Nelson, M.; Dytham, C.; Fitter, A.H.; Helgason, T. Distinct seasonal assemblages of arbuscular mycorrhizal fungi revealed by massively parallel pyrosequencing. New Phytol. 2011, 190, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Davison, J.; Öpik, M.; Zobel, M.; Vasar, M.; Metsis, M.; Moora, M. Communities of arbuscular mycorrhizal fungi detected in forest soil are spatially heterogeneous but do not vary throughout the growing season. PLoS ONE 2012, 7, e41938. [Google Scholar] [CrossRef] [Green Version]
- Davison, J.; Öpik, M.; Daniell, T.J.; Moora, M.; Zobel, M. Arbuscular mycorrhizal fungal communities in plant roots are not random assemblages. FEMS Microbiol. Ecol. 2011, 78, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, S.; Hempel, S.; Verbruggen, E.; Rillig, M.C.; Caruso, T. Linking the community structure of arbuscular mycorrhizal fungi and plants: A story of interdependence? ISME J. 2017, 11, 1400–1411. [Google Scholar] [CrossRef] [Green Version]
- Dumbrell, A.J.; Nelson, M.; Helgason, T.; Dytham, C.; Fitter, A.H. Idiosyncrasy and overdominance in the structure of natural communities of arbuscular mycorrhizal fungi: Is there a role for stochastic processes? J. Ecol. 2010, 98, 419–428. [Google Scholar] [CrossRef]
- Unterseher, M.; Jumpponen, A.; Öpik, M.; Tedersoo, L.; Moora, M.; Dormann, C.F.; Schnittler, M. Species abundance distributions and richness estimations in fungal metagenomics—Lessons learned from community ecology. Mol. Ecol. 2011, 20, 275–285. [Google Scholar] [CrossRef]
- Chen, Y.L.; Xu, Z.W.; Xu, T.L.; Veresoglou, S.D.; Yang, G.W.; Chen, B.D. Nitrogen deposition and precipitation induced phylogenetic clustering of arbuscular mycorrhizal fungal communities. Soil Biol. Biochem. 2017, 115, 233–242. [Google Scholar] [CrossRef]
- Caruso, T.; Powell, J.R.; Rillig, M.C. Compositional divergence and convergence in local communities and spatially structured landscapes. PLoS ONE 2012, 7, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R.; Bennett, A.E. Unpredictable assembly of arbuscular mycorrhizal fungal communities. Pedobiologia 2016, 59, 11–15. [Google Scholar] [CrossRef]
- Deveautour, C.; Donn, S.; Bennett, A.E.; Power, S.; Powell, J.R. Variability of arbuscular mycorrhizal fungal communities within the root systems of individual plants is high and influenced by host species and root phosphorus. Pedobiologia 2021, 84, 150691. [Google Scholar] [CrossRef]
- Klironomos, J.N.; Moutoglis, P. Colonization of nonmycorrhizal plants by mycorrhizal neighbours as influenced by the collembolan, Folsomia candida. Biol. Fertil. Soils 1999, 29, 277–281. [Google Scholar] [CrossRef]
- Hubbell, S.P. The Unified Neutral Theory of Biodiversity and Biogeography; Monographs of Population Biology; Princeton University Press: Princeton, NJ, USA, 2001; Volume 32. [Google Scholar]
- Cadotte, M.W. Competition-colonization trade-offs and disturbance effects at multiple scales. Ecology 2007, 88, 823–829. [Google Scholar] [CrossRef] [Green Version]
- Veresoglou, S.D.; Caruso, T.; Rillig, M.C. Metacommunities and symbiosis: Hosts of challenges. Trends Ecol. Evol. 2012, 27, 588–589. [Google Scholar] [CrossRef]
- Hein, C.L.; Öhlund, G.; Englund, G. Fish introductions reveal the temperature dependence of species interactions. Proc. R. Soc. B Biol. Sci. 2013, 281. [Google Scholar] [CrossRef]
- Mihaljevic, J.R. Linking metacommunity theory and symbiont evolutionary ecology. Trends Ecol. Evol. 2012, 27, 323–329. [Google Scholar] [CrossRef]
- Grünfeld, L.; Wulf, M.; Rillig, M.C.; Manntschke, A.; Veresoglou, S.D. Neighbours of arbuscular-mycorrhiza associating trees are colonized more extensively by arbuscular mycorrhizal fungi than their conspecifics in ectomycorrhiza dominated stands. New Phytol. 2019, 227, 10–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scotton, M. Mountain grassland restoration: Effects of sowing rate, climate and soil on plant density and cover. Sci. Total Environ. 2019, 651, 3090–3098. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular- Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Öpik, M.; Vanatoa, A.; Vanatoa, E.; Moora, M.; Davison, J.; Kalwij, J.M.; Reier, Ü.; Zobel, M. The online database MaarjAM reveals global and ecosystemic distribution patterns in arbuscular mycorrhizal fungi (Glomeromycota). New Phytol. 2010, 188, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Gotelli, N.J. Null model analysis of species co-occurrence patterns. Ecology 2000, 81, 2606–2621. [Google Scholar] [CrossRef]
- Gotelli, N.; Hart, E.; Ellison, A.; Hart, M.E. Package ‘EcoSimR’—Null Model Analysis for Ecological Data. R Packag. 2015.
- Vályi, K.; Mardhiah, U.; Rillig, M.C.; Hempel, S. Community assembly and coexistence in communities of arbuscular mycorrhizal fungi. ISME J. 2016, 10, 2341–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lekberg, Y.; Schnoor, T.; Kjøller, R.; Gibbons, S.M.; Hansen, L.H.; Al-Soud, W.A.; Sørensen, S.J.; Rosendahl, S. 454-Sequencing Reveals Stochastic Local Reassembly and High Disturbance Tolerance Within Arbuscular Mycorrhizal Fungal Communities. J. Ecol. 2012, 100, 151–160. [Google Scholar] [CrossRef]
- Kivlin, S.N.; Hawkes, C.V. Tree species, spatial heterogeneity, and seasonality drive soil fungal abundance, richness, and composition in Neotropical rainforests. Environ. Microbiol. 2016, 18, 4662–4673. [Google Scholar] [CrossRef] [PubMed]
- Maitra, P.; Zheng, Y.; Chen, L.; Wang, Y.L.; Ji, N.N.; Lü, P.P.; Gan, H.Y.; Li, X.C.; Sun, X.; Zhou, X.H.; et al. Effect of drought and season on arbuscular mycorrhizal fungi in a subtropical secondary forest. Fungal Ecol. 2019, 41, 107–115. [Google Scholar] [CrossRef]
- Camenzind, T.; Hempel, S.; Homeier, J.; Horn, S.; Velescu, A.; Wilcke, W.; Rillig, M.C. Nitrogen and phosphorus additions impact arbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest. Glob. Chang. Biol. 2014, 20, 3646–3659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veresoglou, S.D.; Halley, J.M. A model that explains diversity patterns of arbuscular mycorrhizas. Ecol. Model. 2012, 231, 146–152. [Google Scholar] [CrossRef]
- Hammer, E.C.; Pallon, J.; Wallander, H.; Olsson, P.A. Tit for tat? A mycorrhizal fungus accumulates phosphorus under low plant carbon availability. FEMS Microbiol. Ecol. 2011, 76, 236–244. [Google Scholar] [CrossRef] [Green Version]
- Staddon, P.L.; Ramsey, C.B.; Ostle, N.; Ineson, P.; Fitter, A.H. Rapid turnover of hyphae of mycorrhizal fungi determined by AMS microanalysis of 14C. Science 2003, 300, 1138–1140. [Google Scholar] [CrossRef] [Green Version]
- Bar-Massada, A. Complex relationships between species niches and environmental heterogeneity affect species co-occurrence patterns in modelled and real communities. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150927. [Google Scholar] [CrossRef] [Green Version]
- Ning, D.; Deng, Y.; Tiedje, J.M.; Zhou, J. A general framework for quantitatively assessing ecological stochasticity. Proc. Natl. Acad. Sci. USA 2019, 116, 16892–16898. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.O.D.; Pernthaler, J. Biomass addition alters community assembly in ultrafiltration membrane biofilms. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fodelianakis, S.; Valenzuela-Cuevas, A.; Barozzi, A.; Daffonchio, D. Direct quantification of ecological drift at the population level in synthetic bacterial communities. ISME J. 2021, 15, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Veresoglou, S.D.; Wulf, M.; Rillig, M.C. Facilitation between woody and herbaceous plants that associate with arbuscular mycorrhizal fungi in temperate European forests. Ecol. Evol. 2017, 7, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.W.; Grorud-Colvert, K.; Sponaugle, S.; Semmens, B.X. Phenotypic variation and selective mortality as major drivers of recruitment variability in fishes. Ecol. Lett. 2014, 17, 743–755. [Google Scholar] [CrossRef]
- Kadowaki, K.; Yamamoto, S.; Sato, H.; Tanabe, A.S.; Hidaka, A.; Toju, H. Mycorrhizal fungi mediate the direction and strength of plant–soil feedbacks differently between arbuscular mycorrhizal and ectomycorrhizal communities. Commun. Biol. 2018, 1, 196. [Google Scholar] [CrossRef]
- Liang, M.; Johnson, D.; Bursem, F.R.P.; Yu, S.; Fang, M.; Taylor, J.D.; Taylor, A.F.S.; Helgason, T.; Liu, X. Soil fungal networks maintain local dominance of ectomycorrhizal trees. Nat. Commun. 2020, 11, 2636. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the experimental design of the three experiments. (a) In Experiment One and Experiment Two we manipulated the connectance (low; intermediate and high) of four vegetated inserts (dark grey) over an unvegetated soil (sterilized and diluted with sand) matrix (in light grey). In Experiment One we used Plantago lanceolata as a host whereas in Experiment Two Medicago lupulina. (b) In Experiment Three we manipulated the diversity (i.e., only one habitat type; either fertilized or unfertilized or both habitat types) and spatial structure (overdispersed vs. aggregated in the bottom two subpanels) of the vegetated inserts which we describe earlier (Top and bottom left archipelagos/treatments: 4 replicates/were each replicated four times; bottom right archipelagos/treatment: six replicates/ was replicated six times).We used Medicago lupulina as a host and the matrix soil was (like in the other experiments) sterilized, mixed with sand and was kept unvegetated (light grey).
Figure 1.
Schematic representation of the experimental design of the three experiments. (a) In Experiment One and Experiment Two we manipulated the connectance (low; intermediate and high) of four vegetated inserts (dark grey) over an unvegetated soil (sterilized and diluted with sand) matrix (in light grey). In Experiment One we used Plantago lanceolata as a host whereas in Experiment Two Medicago lupulina. (b) In Experiment Three we manipulated the diversity (i.e., only one habitat type; either fertilized or unfertilized or both habitat types) and spatial structure (overdispersed vs. aggregated in the bottom two subpanels) of the vegetated inserts which we describe earlier (Top and bottom left archipelagos/treatments: 4 replicates/were each replicated four times; bottom right archipelagos/treatment: six replicates/ was replicated six times).We used Medicago lupulina as a host and the matrix soil was (like in the other experiments) sterilized, mixed with sand and was kept unvegetated (light grey).

Figure 2.
Alpha diversity statistics across treatments and habitats in (A) Experiment One; (B) Experiment Two; (C) Experiment Three. Each experimental unit contained several inserts and we assayed the mycorrhizal community independently for each insert. We observed no differences in alpha diversity in all three experiments. Main panels depict alpha diversity across individual samples whereas the panel inserts show the results after averaging the four (Experiment One and Experiment Two) or eight (Experiment Three) estimates of alpha diversity per experimental unit. Note the lack of differences in relation to alpha diversity. We observed comparable trends for gamma diversity.
Figure 2.
Alpha diversity statistics across treatments and habitats in (A) Experiment One; (B) Experiment Two; (C) Experiment Three. Each experimental unit contained several inserts and we assayed the mycorrhizal community independently for each insert. We observed no differences in alpha diversity in all three experiments. Main panels depict alpha diversity across individual samples whereas the panel inserts show the results after averaging the four (Experiment One and Experiment Two) or eight (Experiment Three) estimates of alpha diversity per experimental unit. Note the lack of differences in relation to alpha diversity. We observed comparable trends for gamma diversity.

Figure 3.
Jaccard index statistics from pairwise comparisons of samples within experimental units in (A) Experiment One; (B) Experiment Two; (C) Experiment Three. Note in panels (A,B) that in intermediate connected spatial arrangements we observe higher Jaccard distances between long-distance compared to short-distance “patches” (in the case of Experiment Two a trend) within-subjects distance effect; F1,49 = 6.34, p = 0.015 in Experiment One; F1,49 = 2.12, p = 0.15 in Experiment Two) and in panel (C) that Jaccard distances differed (F2, 207= 4, p = 0.02) within experimental units in the overdispersed and aggregated treatment.
Figure 3.
Jaccard index statistics from pairwise comparisons of samples within experimental units in (A) Experiment One; (B) Experiment Two; (C) Experiment Three. Note in panels (A,B) that in intermediate connected spatial arrangements we observe higher Jaccard distances between long-distance compared to short-distance “patches” (in the case of Experiment Two a trend) within-subjects distance effect; F1,49 = 6.34, p = 0.015 in Experiment One; F1,49 = 2.12, p = 0.15 in Experiment Two) and in panel (C) that Jaccard distances differed (F2, 207= 4, p = 0.02) within experimental units in the overdispersed and aggregated treatment.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Veresoglou, S.D.; Grünfeld, L.; Mola, M. Micro-Landscape Dependent Changes in Arbuscular Mycorrhizal Fungal Community Structure. Appl. Sci. 2021, 11, 5297. https://0-doi-org.brum.beds.ac.uk/10.3390/app11115297
AMA Style
Veresoglou SD, Grünfeld L, Mola M. Micro-Landscape Dependent Changes in Arbuscular Mycorrhizal Fungal Community Structure. Applied Sciences. 2021; 11(11):5297. https://0-doi-org.brum.beds.ac.uk/10.3390/app11115297
Chicago/Turabian StyleVeresoglou, Stavros D, Leonie Grünfeld, and Magkdi Mola. 2021. "Micro-Landscape Dependent Changes in Arbuscular Mycorrhizal Fungal Community Structure" Applied Sciences 11, no. 11: 5297. https://0-doi-org.brum.beds.ac.uk/10.3390/app11115297
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.