
In Vitro Characterization of Periodontal Ligament Stem Cells Derived from Supernumerary Teeth in Three-Dimensional Culture Method

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. 2D and 3D Culture of Human sPDLSCs
2.2. Morphology
2.3. Assays of Cell Viability
2.4. Osteogenic Differentiation
2.5. Gene Expression
2.5.1. RNA Isolation
2.5.2. Library Preparation and Sequencing
2.5.3. Data Processing for the Identification of Differentially Expressed Genes
2.6. Statistical Analyses
3. Results
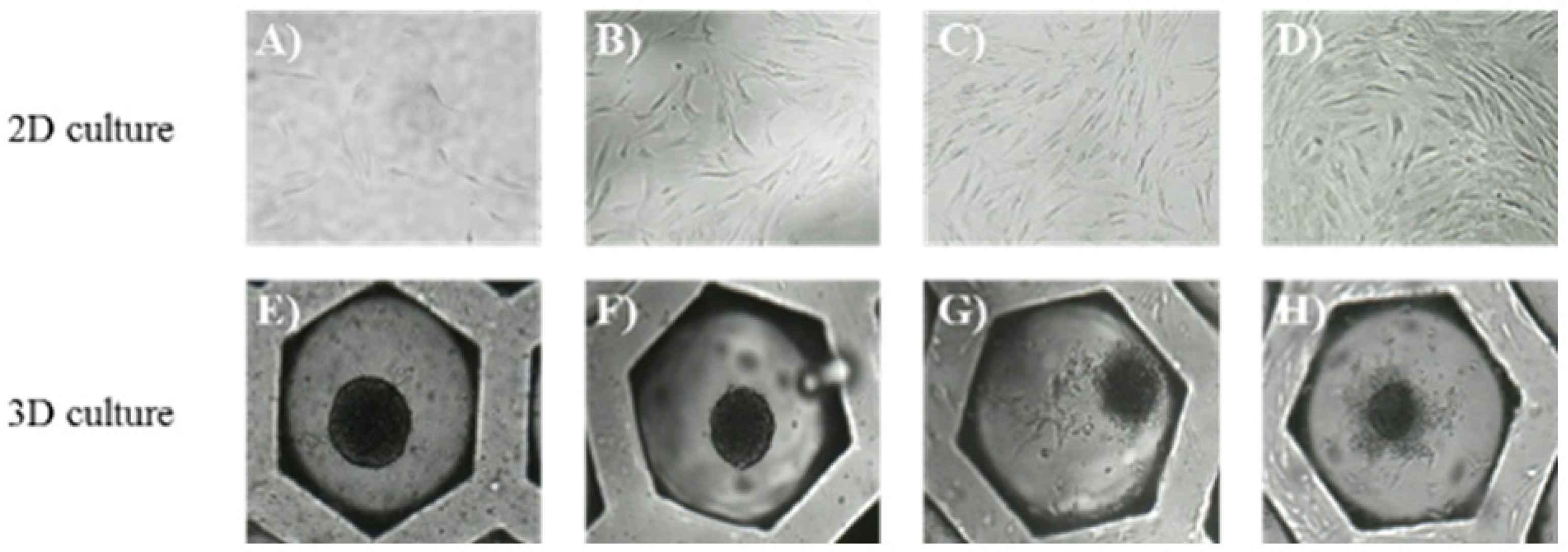
3.1. Morphology
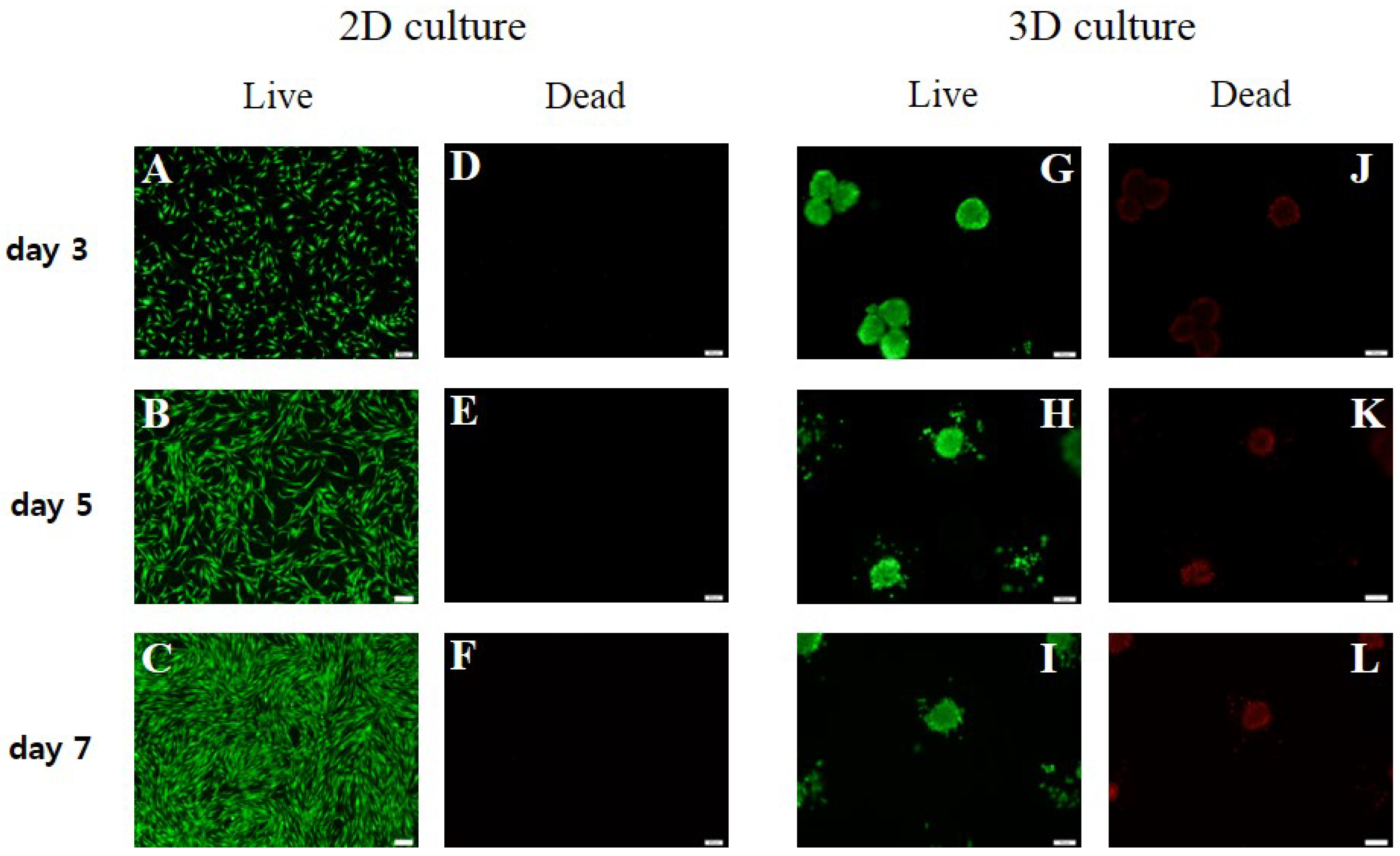
3.2. Cell Viability
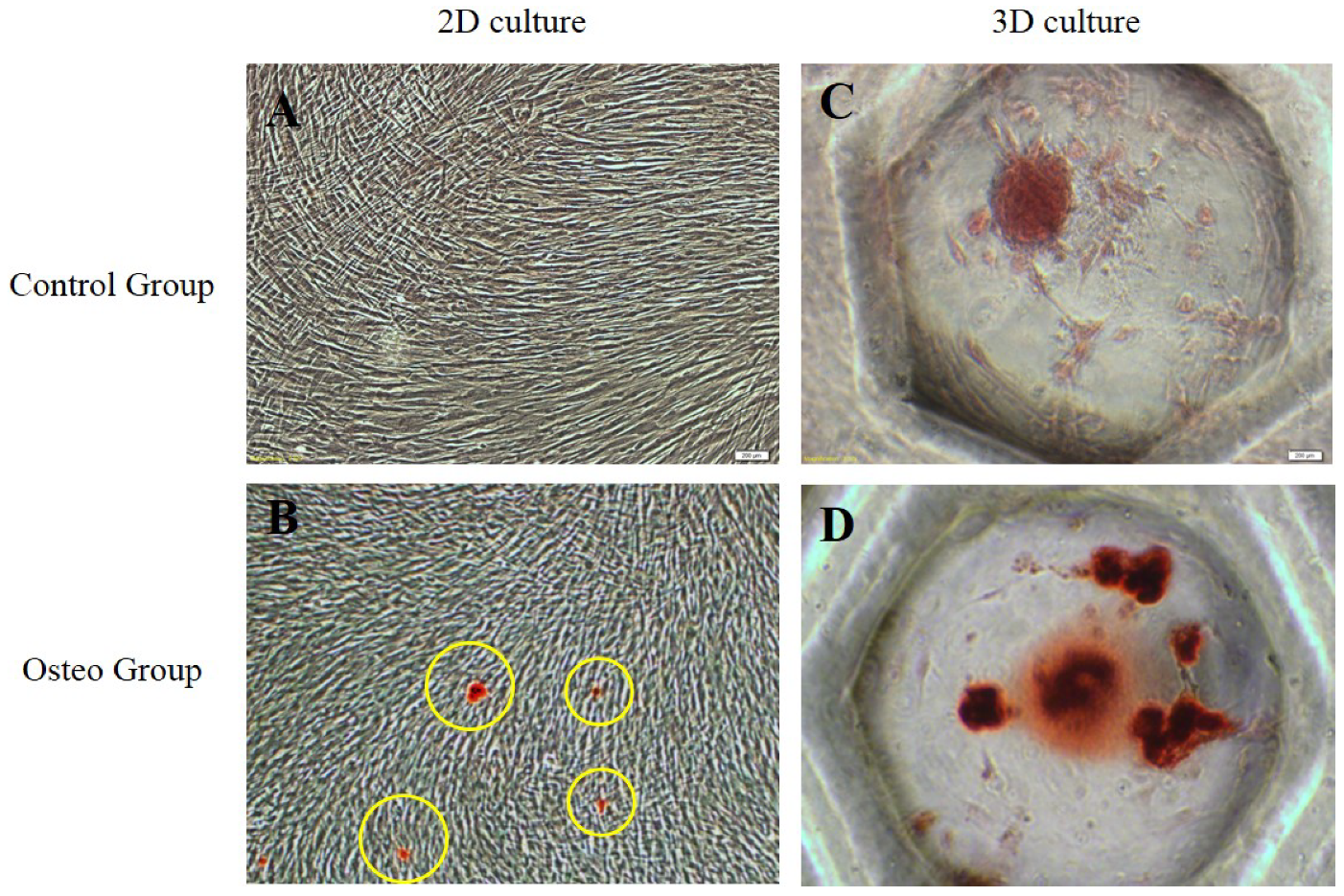
3.3. Stemness and Osteogenic Differentiation
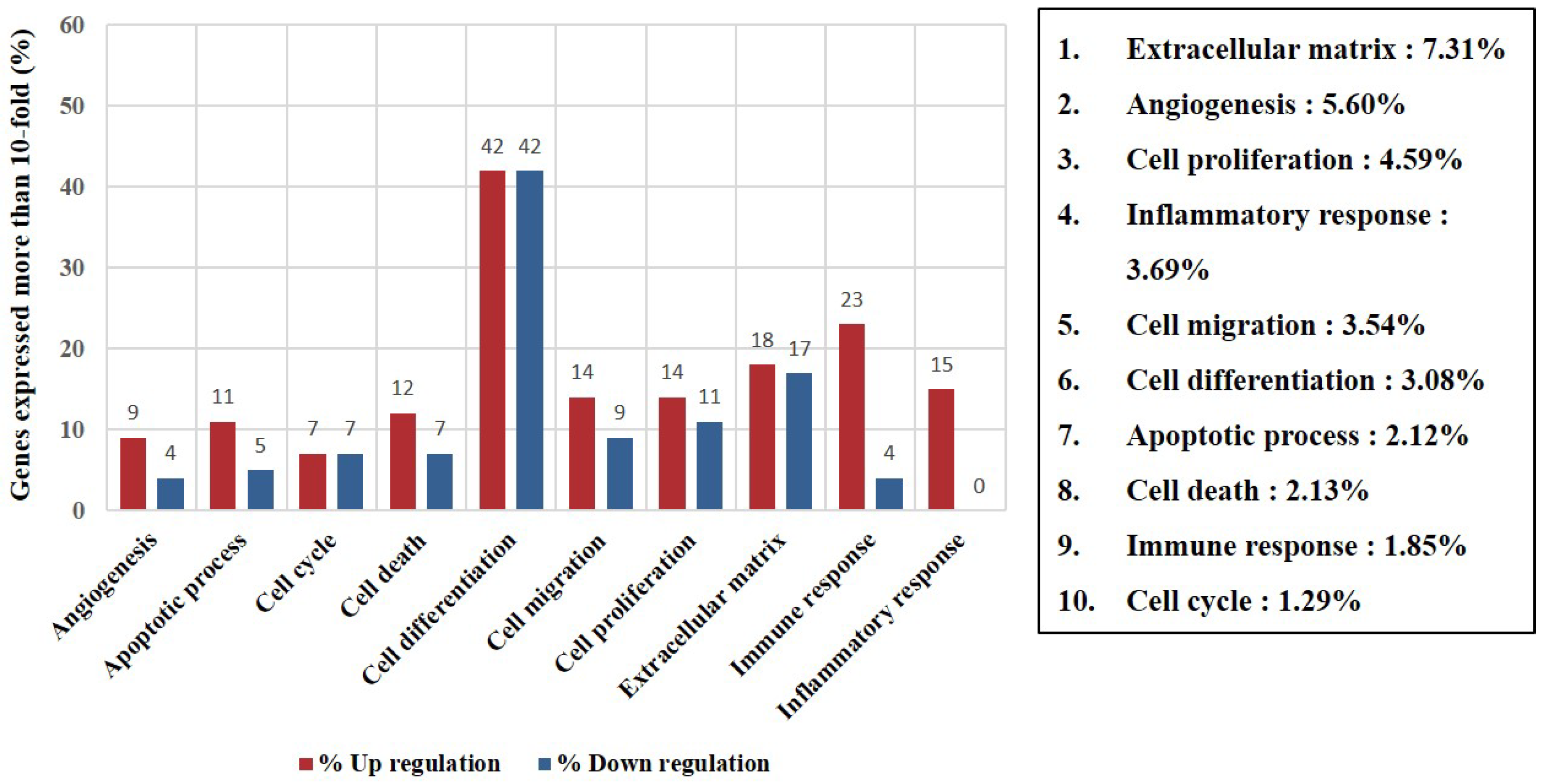
3.4. Identification of Differentially Expressed Genes and Gene Ontology Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Summary of Abbreviations
| Abbreviations | Definition |
| 2D | Two-dimensional |
| 3D | Three-dimensional |
| ARS | Alizarin red S |
| ECM | Extracellular matrix |
| HIF | Hypoxia inducible factor |
| MSCs | Mesenchymal stem cells |
| NADP | Nicotinamide adenine dinucleotide phosphate |
| NADPH | Nicotinamide adenine dinucleotide phosphate hydrogen |
| PDL | Periodontal ligament |
| PDLSCs | Periodontal ligament stem cells |
| sPDLSCs | periodontal ligament stem cells derived from supernumerary teeth |
References
- Jin, H.J.; Bae, Y.K.; Kim, M.; Kwon, S.J.; Jeon, H.B.; Choi, S.J.; Kim, S.W.; Yang, Y.S.; Oh, W.; Chang, J.W. Comparative analysis of human mesenchymal stem cells from bone marrow, adipose tissue, and umbilical cord blood as sources of cell therapy. Int. J. Mol. Sci. 2013, 14, 17986–18001. [Google Scholar] [CrossRef]
- Wong, S.F.; da No, Y.; Choi, Y.Y.; Kim, D.S.; Chung, B.G.; Lee, S.H. Concave microwell based size-controllable hepatosphere as a three-dimensional liver tissue model. Biomaterials 2011, 32, 8087–8096. [Google Scholar] [CrossRef]
- Lee, S.I.; Ko, Y.; Park, J.B. Evaluation of the maintenance of stemness, viability, and differentiation potential of gingiva-derived stem-cell spheroids. Exp. Ther. Med. 2017, 13, 1757–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Baipaywad, P.; Jeong, Y.; Park, H. Incorporation of gelatin microparticles on the formation of adipose-derived stem cell spheroids. Int. J. Biol. Macromol. 2018, 110, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Di Vito, A.; Giudice, A.; Chiarella, E.; Malara, N.; Bennardo, F.; Fortunato, L. In Vitro Long-Term Expansion and High Osteogenic Potential of Periodontal Ligament Stem Cells: More Than a Mirage. Cell Transpl. 2019, 28, 129–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.I.; Ko, Y.; Park, J.B. Evaluation of the shape, viability, stemness and osteogenic differentiation of cell spheroids formed from human gingiva-derived stem cells and osteoprecursor cells. Exp. Ther. Med. 2017, 13, 3467–3473. [Google Scholar] [CrossRef]
- Lee, S.I.; Yeo, S.I.; Kim, B.B.; Ko, Y.; Park, J.B. Formation of size-controllable spheroids using gingiva-derived stem cells and concave microwells: Morphology and viability tests. Biomed. Rep. 2016, 4, 97–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Pjesivac-Grbovic, J.; Cantrell, C.; Freyer, J.P. A multiscale model for avascular tumor growth. Biophys. J. 2005, 89, 3884–3894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.Y.; Kim, M.S.; Lee, K.E.; Nam, O.H.; Jang, J.-H.; Choi, S.-C.; Lee, H.-S. Comparison of 2- and 3-Dimensional Cultured Periodontal Ligament Stem Cells. Pilot Study 2021, 11, 1083. [Google Scholar]
- Curcio, E.; Salerno, S.; Barbieri, G.; De Bartolo, L.; Drioli, E.; Bader, A. Mass transfer and metabolic reactions in hepatocyte spheroids cultured in rotating wall gas-permeable membrane system. Biomaterials 2007, 28, 5487–5497. [Google Scholar] [CrossRef]
- Mueller-Klieser, W. Method for the determination of oxygen consumption rates and diffusion coefficients in multicellular spheroids. Biophys. J. 1984, 46, 343–348. [Google Scholar] [CrossRef]
- Cesarz, Z.; Tamama, K. Spheroid Culture of Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 9176357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.Z.; Chang, H.Y. Recent advances in three-dimensional multicellular spheroid culture for biomedical research. Biotechnol. J. 2008, 3, 1172–1184. [Google Scholar] [CrossRef]
- Hildebrandt, C.; Buth, H.; Thielecke, H. A scaffold-free in vitro model for osteogenesis of human mesenchymal stem cells. Tissue Cell 2011, 43, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kawashima, N.; Takashino, N.; Koizumi, Y.; Takimoto, K.; Suzuki, N.; Saito, M.; Suda, H. Three-dimensional spheroid culture promotes odonto/osteoblastic differentiation of dental pulp cells. Arch. Oral Biol. 2014, 59, 310–317. [Google Scholar] [CrossRef]
- Yeh, H.Y.; Liu, B.H.; Sieber, M.; Hsu, S.H. Substrate-dependent gene regulation of self-assembled human MSC spheroids on chitosan membranes. BMC Genom. 2014, 15, 10. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, H.; Suzuki, K.; Sakai, N.; Yamada, S. Odontoblasts induced from mesenchymal cells of murine dental papillae in three-dimensional cell culture. Cell Tissue Res. 2004, 317, 173–185. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Ohno, J.; Sato, A.; Kido, H.; Fukushima, T. Mesenchymal stem cell spheroids exhibit enhanced in-vitro and in-vivo osteoregenerative potential. BMC Biotechnol. 2014, 14, 105. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Inaba, A.; Mohindroo, N.; Ganesh, D.; Martin, C.E.; Chugal, N.; Kim, R.H.; Kang, M.K.; Park, N.H.; Shin, K.H. Three-dimensional Sphere-forming Cells Are Unique Multipotent Cell Population in Dental Pulp Cells. J. Endod. 2017, 43, 1302–1308. [Google Scholar] [CrossRef]
- Bellotti, C.; Duchi, S.; Bevilacqua, A.; Lucarelli, E.; Piccinini, F. Long term morphological characterization of mesenchymal stromal cells 3D spheroids built with a rapid method based on entry-level equipment. Cytotechnology 2016, 68, 2479–2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sart, S.; Tsai, A.C.; Li, Y.; Ma, T. Three-dimensional aggregates of mesenchymal stem cells: Cellular mechanisms, biological properties, and applications. Tissue Eng. Part B Rev. 2014, 20, 365–380. [Google Scholar] [CrossRef] [Green Version]
- Ishiyama, M.; Miyazono, Y.; Sasamoto, K.; Ohkura, Y.; Ueno, K. A highly water-soluble disulfonated tetrazolium salt as a chromogenic indicator for NADH as well as cell viability. Talanta 1997, 44, 1299–1305. [Google Scholar] [CrossRef]
- Amirikia, M.; Ali Jorsaraei, S.G.; Ali Shariatzadeh, S.M.; Mehranjani, M.S. Differentiation of stem cells from the apical papilla into osteoblasts by the elastic modulus of porous silk fibroin scaffolds. Biologicals 2019, 57, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fennema, E.; Rivron, N.; Rouwkema, J.; van Blitterswijk, C.; de Boer, J. Spheroid culture as a tool for creating 3D complex tissues. Trends Biotechnol. 2013, 31, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.Z.; van den Beucken, J.; Yuan, C.; Jansen, J.A.; Yang, F. Spheroid formation and stemness preservation of human periodontal ligament cells on chitosan films. Oral. Dis. 2018, 24, 1083–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okumura, K.; Nakamura, K.; Hisatomi, Y.; Nagano, K.; Tanaka, Y.; Terada, K.; Sugiyama, T.; Umeyama, K.; Matsumoto, K.; Yamamoto, T.; et al. Salivary gland progenitor cells induced by duct ligation differentiate into hepatic and pancreatic lineages. Hepatology 2003, 38, 104–113. [Google Scholar] [CrossRef]
- Zhao, S.; Fung-Leung, W.P.; Bittner, A.; Ngo, K.; Liu, X. Comparison of RNA-Seq and microarray in transcriptome profiling of activated T cells. PLoS ONE 2014, 9, e78644. [Google Scholar] [CrossRef]
- Laschke, M.W.; Schank, T.E.; Scheuer, C.; Kleer, S.; Schuler, S.; Metzger, W.; Eglin, D.; Alini, M.; Menger, M.D. Three-dimensional spheroids of adipose-derived mesenchymal stem cells are potent initiators of blood vessel formation in porous polyurethane scaffolds. Acta Biomater. 2013, 9, 6876–6884. [Google Scholar] [CrossRef] [PubMed]
- Tsai, A.C.; Liu, Y.; Yuan, X.; Ma, T. Compaction, fusion, and functional activation of three-dimensional human mesenchymal stem cell aggregate. Tissue Eng. Part A 2015, 21, 1705–1719. [Google Scholar] [CrossRef] [Green Version]
- Bartosh, T.J.; Ylostalo, J.H.; Bazhanov, N.; Kuhlman, J.; Prockop, D.J. Dynamic compaction of human mesenchymal stem/precursor cells into spheres self-activates caspase-dependent IL1 signaling to enhance secretion of modulators of inflammation and immunity (PGE2, TSG6, and STC1). Stem Cells 2013, 31, 2443–2456. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Fold Change | Description | Related Function |
|---|---|---|---|
| AREG | 1172.7 | Amphiregulin | Cell differentiation, cell proliferation |
| SMOC1 | 1161.3 | SPARC-related modular calcium binding 1 | Extracellular matrix |
| MMP1 | 829.2 | Matrix metallopeptidase 1 | Cell migration, extracellular matrix |
| STC1 | 791.3 | Stanniocalcin 1 | Cell differentiation, cell proliferation |
| MMP10 | 349.8 | Matrix metallopeptidase 10 | Extracellular matrix |
| PGF | 341.9 | Placental growth factor | Angiogenesis |
| PTGS2 | 195.6 | Prostaglandin-endoperoxide synthase 2 | Angiogenesis, cell differentiation, inflammatory response |
| COL5A3 | 173.4 | Collagen type V alpha 3 | Extracellular matrix |
| IL24 | 156.0 | Interleukin 24 | Apoptotic process, cell death |
| FRZB | 119.4 | Frizzled-related protein | Cell differentiation |
| BMP2 | 112.8 | Bone morphogenetic protein 2 | Cell differentiation, cell proliferation, inflammatory response |
| SFRP2 | 104.5 | Secreted frizzled-related protein 2 | Angiogenesis, apoptotic process, cell death, cell differentiation, cell proliferation, extracellular matrix |
| FGL2 | 98.6 | Fibrinogen-like 2 | Immune response |
| IER3 | 97.3 | Immediate early response 3 | Apoptotic process, cell death |
| TFPI2 | 94.9 | Tissue factor pathway inhibitor 2 | Extracellular matrix |
| DPT | 83.6 | Dermatopontin | Extracellular matrix |
| NR4A2 | 82.5 | Nuclear receptor subfamily 4 group A member 2 | Cell differentiation, cell migration |
| TNC | 82.1 | Tenascin C | Cell differentiation, extracellular matrix |
| NPNT | 73.8 | Nephronectin | Cell differentiation, extracellular matrix |
| NDP | 64.7 | Norrie disease (pseudoglioma) | Cell proliferation, extracellular matrix |
| ACKR4 | 64.5 | Atypical chemokine receptor 4 | Immune response |
| IL18R1 | 63.5 | Interleukin 18 receptor 1 | Cell differentiation, immune response |
| MMP8 | 62.5 | Matrix metallopeptidase 8 | Extracellular matrix |
| SPRY1 | 61.6 | Sprouty RTK signaling antagonist 1 | Cell cycle, cell differentiation |
| RGS2 | 61.4 | Regulator of G-protein signaling 2 | Cell cycle, cell differentiation |
| IL1R1 | 60.3 | Interleukin 1 receptor type 1 | Immune response |
| PTGS1 | 60.3 | Prostaglandin-endoperoxide synthase 1 | Inflammatory response |
| SPON1 | 53.6 | Spondin 1 | Extracellular matrix |
| EDNRB | 52.4 | Endothelin receptor type B | Cell differentiation, cell migration |
| PECAM1 | 49.3 | Platelet and endothelial cell adhesion molecule 1 | Angiogenesis, cell differentiation, cell migration |
| IL1RN | 46.1 | Interleukin 1 receptor antagonist | Immune response, inflammatory response |
| PRDM1 | 44.6 | PR domain 1 | Cell differentiation, cell proliferation, immune response |
| CHST2 | 42.1 | Carbohydrate sulfotransferase 2 | Inflammatory response |
| APOE | 40.0 | Apolipoprotein E | Cell differentiation, extracellular matrix |
| NOX4 | 38.8 | NADPH oxidase 4 | Cell differentiation, inflammatory response |
| SNAP25 | 38.2 | Synaptosome-associated protein 25 kDa | Cell differentiation, immune response |
| TNN | 36.8 | Tenascin N | Cell differentiation, extracellular matrix |
| VAV3 | 34.8 | VAV guanine nucleotide exchange factor 3 | Angiogenesis, cell migration |
| BCL2A1 | 31.5 | BCL2-related protein A1 | Apoptotic process, cell death |
| JUP | 30.3 | Junction plakoglobin | Cell death, cell differentiation, cell migration, extracellular matrix, immune response |
| AKR1C1 | 30.3 | Aldo-keto reductase family 1, member C1 | Cell differentiation |
| NRN1 | 29.8 | Neuritin 1 | Cell differentiation |
| ITGA2 | 28.6 | Integrin subunit alpha 2 | Cell differentiation, cell migration, cell proliferation |
| SLC1A3 | 28.3 | Solute carrier family 1 member 3 | Cell differentiation |
| SECTM1 | 27.7 | Secreted and transmembrane 1 | Immune response |
| CD7 | 27.2 | CD7 molecule | Immune response |
| AKR1C2 | 27.1 | Aldo-keto reductase family 1, member C2 | Cell differentiation |
| AIM2 | 26.7 | Absent in melanoma 2 | Apoptotic process, cell death, immune response, inflammatory response |
| GPM6B | 26.5 | Glycoprotein M6B | Cell differentiation |
| ETV1 | 26.0 | ETS Variant 1 | Cell differentiation |
| CXCL8 | 24.7 | C-X-C motif chemokine ligand 8 | Angiogenesis, cell cycle, cell migration, immune response, inflammatory response |
| NKX2-5 | 24.5 | NK2 homeobox 5 | Apoptotic process, cell death, cell differentiation, cell proliferation |
| TNFAIP6 | 24.5 | TNF-alpha-induced protein 6 | Immune response, inflammatory response |
| NEFM | 23.6 | Neurofilament, medium polypeptide | Cell differentiation |
| CCL2 | 23.3 | C-C motif chemokine ligand 2 | Cell migration, immune response |
| KYNU | 22.7 | Kynureninase | Immune response |
| RANBP3L | 22.5 | RAN-binding protein 3-like | Cell cycle, cell differentiation |
| NTN1 | 21.6 | Netrin 1 | Apoptotic process, cell death, cell differentiation, cell migration, extracellular matrix |
| SEPT3 | 21.0 | Septin 3 | Cell cycle |
| GABRB2 | 20.4 | Gamma-aminobutyric acid type A receptor beta2 subunit | Cell differentiation |
| EDNRA | 20.3 | Endothelin receptor type A | Angiogenesis, cell differentiation, cell proliferation |
| CHST1 | 19.5 | Carbohydrate sulfotransferase 1 | Inflammatory response |
| AFAP1L2 | 19.5 | Actin filament-associated protein 1-like 2 | Inflammatory response |
| CRYGD | 19.4 | Crystallin, gamma D | Cell differentiation |
| CMKLR1 | 18.6 | Chemerin chemokine-like receptor 1 | Immune response |
| WNT7B | 17.4 | Wnt family member 7B | Cell differentiation, cell proliferation, extracellular matrix |
| PTPRU | 16.5 | Protein tyrosine phosphatase, receptor type U | Cell differentiation |
| ITGB3 | 16.4 | Integrin subunit beta 3 | Angiogenesis, cell differentiation, cell migration |
| TNFRSF21 | 15.5 | Tumor necrosis factor receptor superfamily member 21 | Apoptotic process, cell death, cell differentiation, immune response |
| NYAP1 | 14.4 | Neuronal tyrosine phosphorylated phosphoinositide-3-kinase adaptor 1 | Cell differentiation |
| GRIN2A | 13.3 | Glutamate ionotropic receptor NMDA type subunit 2A | Cell differentiation |
| PHOX2B | 12.8 | Paired-like homeobox 2b | Cell differentiation, cell migration |
| GPR68 | 12.8 | G-protein-coupled receptor 68 | Inflammatory response |
| FPR1 | 12.7 | Formyl peptide receptor 1 | Cell migration, immune response, inflammatory response |
| EREG | 12.6 | Epiregulin | Angiogenesis, cell cycle, cell differentiation, cell proliferation |
| FGF7 | 12.5 | Fibroblast growth factor 7 | Cell proliferation |
| TNFSF10 | 12.1 | Tumor necrosis factor superfamily member 10 | Apoptotic process, cell death, immune response |
| LRP4 | 11.5 | LDL receptor-related protein 4 | Cell differentiation |
| PTPN22 | 11.3 | Protein tyrosine phosphatase, non-receptor type 22 | Cell differentiation |
| RNF152 | 11.3 | Ring finger protein 152 | Apoptotic process, cell death |
| SOCS2 | 11.3 | Suppressor of cytokine signaling 2 | Cell differentiation |
| CXCL3 | 11.2 | CXC motif chemokine ligand 3 | Cell migration, immune response, inflammatory response |
| GEM | 11.1 | GTP-binding protein overexpressed in skeletal muscle | Cell cycle, immune response |
| ZNF443 | 10.8 | Zinc finger protein 443 | Apoptotic process, cell death |
| MGP | 10.7 | Matrix Gla protein | Extracellular matrix |
| TNIK | 10.5 | TRAF2 and NCK interacting kinase | Cell differentiation |
| NR4A3 | 10.3 | Nuclear receptor subfamily 4 group A member 3 | Cell differentiation, cell proliferation, immune response |
| CSGALNACT1 | 10.3 | Chondroitin sulfate N-acetylgalactosaminyltransferase 1 | Cell proliferation |
| CTSS | 10.3 | Cathepsin S | Immune response |
| Gene Name | Fold Change | Description | Related Function |
|---|---|---|---|
| ACAN | 177.4 | Aggrecan | Cell differentiation, extracellular matrix |
| NGF | 136.7 | Nerve growth factor | Apoptotic process, cell death, cell differentiation |
| HAPLN1 | 106.0 | Hyaluronan and proteoglycan link protein 1 | Extracellular matrix |
| NPR3 | 102.8 | Natriuretic peptide receptor 3 | Cell proliferation |
| CTNND2 | 86.6 | Catenin delta 2 | Cell differentiation |
| ADM2 | 81.4 | Adrenomedullin 2 | Angiogenesis |
| TRIM67 | 58.4 | Tripartite motif containing 67 | Cell differentiation |
| ADAMTS14 | 55.6 | ADAM metallopeptidase with thrombospondin type 1 motif 14 | Extracellular matrix |
| DLGAP5 | 52.7 | Discs large homolog-associated protein 5 | Cell cycle, cell proliferation |
| ADRA1B | 45.1 | Adrenoceptor alpha 1B | Cell proliferation |
| LGR5 | 45.1 | Leucine-rich repeat-containing G-protein-coupled receptor 5 | Cell differentiation, cell proliferation |
| TENM2 | 41.3 | Teneurin transmembrane protein 2 | Cell differentiation |
| SLITRK1 | 41.2 | SLIT and NTRK like family member 1 | Cell differentiation |
| MAP2 | 38.3 | Microtubule associated protein 2 | Cell differentiation |
| IL7R | 35.5 | Interleukin 7 receptor | Cell differentiation, cell proliferation, immune response |
| GLI1 | 33.3 | GLI family zinc finger 1 | Cell differentiation |
| KRT33B | 32.6 | Keratin 33B | Cell death, cell differentiation |
| COL1A1 | 27.8 | Collagen type I alpha 1 | Cell differentiation, cell migration, extracellular matrix |
| HILS1 | 25.5 | Histone linker H1 domain, spermatid-specific 1 (pseudogene) | Cell differentiation |
| ANK2 | 25.5 | Ankyrin 2, neuronal | Cell differentiation |
| PRELP | 25.0 | Proline/arginine-rich end leucine-rich repeat protein | Cell differentiation, extracellular matrix |
| KRT34 | 23.2 | Keratin 34 | Cell death, cell differentiation |
| TTK | 19.6 | TTK protein kinase | Cell cycle |
| ASB2 | 18.7 | Ankyrin repeat and SOCS box-containing 2 | Cell differentiation |
| PSG1 | 17.8 | Pregnancy-specific beta-1-glycoprotein 1 | Cell migration |
| ASPN | 16.9 | Asporin | Extracellular matrix |
| RSPO2 | 16.5 | R-Spondin 2 | Cell differentiation |
| TAGLN | 15.7 | Transgelin | Cell differentiation |
| FBN2 | 15.3 | Fibrillin 2 | Extracellular matrix |
| LOX | 15.1 | Lysyl oxidase | Extracellular matrix |
| SEMA3D | 14.4 | Semaphorin 3D | Cell differentiation |
| LEP | 14.0 | Leptin | Cell differentiation, cell migration |
| COL6A6 | 13.6 | Collagen type VI alpha 6 | Extracellular matrix |
| MMP15 | 13.6 | Matrix metallopeptidase 15 | Extracellular matrix |
| CREB3L1 | 13.5 | cAMP-responsive element-binding protein 3-like 1 | Cell differentiation |
| EFHD1 | 13.5 | EF-hand domain family member D1 | Cell differentiation |
| KRTAP1-5 | 13.4 | Keratin-associated protein 1-5 | Cell differentiation |
| ANLN | 13.3 | Anillin actin-binding protein | Cell cycle, cell differentiation, cell migration |
| HLA-DMA | 13.1 | Major histocompatibility complex, class II, DM alpha | Immune response |
| MKI67 | 13.0 | Marker of proliferation Ki-67 | Cell cycle, cell proliferation |
| GDF6 | 12.9 | Growth differentiation factor 6 | Cell death, cell differentiation |
| ANKRD1 | 12.7 | Ankyrin repeat domain 1 | Cell differentiation |
| BCL2 | 12.6 | B-cell CLL/lymphoma 2 | Cell death, cell differentiation, cell proliferation |
| RIMS1 | 12.5 | Regulating synaptic membrane exocytosis 1 | Cell differentiation |
| ECM2 | 12.4 | Extracellular matrix protein 2 | Cell differentiation, extracellular matrix |
| TUBB2B | 12.2 | Tubulin beta 2B class IIb | Cell differentiation, cell migration |
| CLDN1 | 12.0 | Claudin 1 | Cell differentiation, immune response |
| AURKB | 11.7 | Aurora kinase B | Cell cycle |
| RTN1 | 11.6 | Reticulon 1 | Cell differentiation |
| BCAT1 | 11.6 | Branched chain amino acid transaminase 1 | Cell cycle, cell proliferation |
| KIRREL3 | 11.5 | Kin of IRRE-like 3 (Drosophila) | Cell differentiation, cell migration |
| SLC4A5 | 11.2 | Solute carrier family 4 member 5 | Cell differentiation |
| MYC | 11.1 | v-Myc avian myelocytomatosis viral oncogene homolog | Cell cycle, cell differentiation, cell proliferation, immune response |
| CYR61 | 11.0 | Cysteine-rich angiogenic inducer 61 | Angiogenesis, apoptotic process, cell death, cell differentiation, cell migration, cell proliferation, extracellular matrix |
| CCBE1 | 11.0 | Collagen and calcium-binding EGF domains 1 | Angiogenesis, extracellular matrix |
| TPM1 | 10.6 | Tropomyosin 1 (alpha) | Cell differentiation |
| CHAC1 | 10.5 | ChaC glutathione-specific gamma-glutamylcyclotransferase 1 | Apoptotic process, cell death, cell differentiation |
| CTGF | 10.4 | Connective tissue growth factor | Angiogenesis, cell differentiation, cell migration, cell proliferation, extracellular matrix |
| NAV2 | 10.3 | Neuron navigator 2 | Extracellular matrix |
| ETV7 | 10.2 | ETS Variant 7 | Cell differentiation |
| CLEC3B | 10.1 | C-type lectin domain family 3 member B | Extracellular matrix |
| MATN2 | 10.1 | Matrilin 2 | Cell differentiation, cell migration, extracellular matrix |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, Y.Y.; Kim, M.S.; Lee, K.E.; Nam, O.H.; Jang, J.-H.; Choi, S.-C.; Lee, H.-S. In Vitro Characterization of Periodontal Ligament Stem Cells Derived from Supernumerary Teeth in Three-Dimensional Culture Method. Appl. Sci. 2021, 11, 6040. https://0-doi-org.brum.beds.ac.uk/10.3390/app11136040
Jeong YY, Kim MS, Lee KE, Nam OH, Jang J-H, Choi S-C, Lee H-S. In Vitro Characterization of Periodontal Ligament Stem Cells Derived from Supernumerary Teeth in Three-Dimensional Culture Method. Applied Sciences. 2021; 11(13):6040. https://0-doi-org.brum.beds.ac.uk/10.3390/app11136040
Chicago/Turabian StyleJeong, Yun Yeong, Mi Sun Kim, Ko Eun Lee, Ok Hyung Nam, Ji-Hyun Jang, Sung-Chul Choi, and Hyo-Seol Lee. 2021. "In Vitro Characterization of Periodontal Ligament Stem Cells Derived from Supernumerary Teeth in Three-Dimensional Culture Method" Applied Sciences 11, no. 13: 6040. https://0-doi-org.brum.beds.ac.uk/10.3390/app11136040