
Effects of a Dual-Purpose Inoculant on the Quality and Aerobic Stability of Corn Silage at the Laboratory and Field Scales

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Silage Production
2.2. Chemical Composition
2.3. Microbial Counts and Aerobic Stability
2.4. Statistical Analysis
3. Results
3.1. Laboratory-Scale Silage
3.2. Baled Silage
3.3. Bunker Silage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, D.; Ni, K.; Zhang, Y.; Lin, Y.; Yang, F. Fermentation characteristics, chemical composition and microbial community of tropical forage silage under different temperatures. Asian Australas. J. Anim. 2019, 32, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, C.; Zhou, W.; Yang, F.-Y.; Chen, X.-Y.; Zhang, Q. Effects of wilting and Lactobacillus plantarum addition on the fermentation quality and microbial community of Moringa oleifera leaf silage. Front. Microbiol. 2018, 9, 1817–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Thévenot-Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Anim. Feed Sci. Technol. 2013, 182, 1–15. [Google Scholar] [CrossRef]
- Wilkinson, J.M.; Davies, D.R. The aerobic stability of silage: Key findings and recent developments. Grass Forage Sci. 2013, 68, 1–19. [Google Scholar] [CrossRef]
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [Green Version]
- Boudra, H.; Rouillé, B.; Lyan, B.; Morgavi, D.P. Presence of mycotoxins in sugar beet pulp silage collected in France. Anim. Feed Sci. Technol. 2015, 205, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Magaji, U.; Hussin, G.; Ramli, A.; Miah, G. Fermentation quality and additives: A case of rice straw silage. BioMed Res. Int. 2016, 2016, 7985167. [Google Scholar] [CrossRef] [Green Version]
- Kung, L., Jr.; Stokes, M.R.; Lin, C. Silage additives. In Silage Science and Technology, 1st ed.; Buxton, D.R., Muck, R.E., Harrison, J.H., Eds.; Crop Science of America, Inc.: Madison, WI, USA, 2003; Volume 42, pp. 305–360. [Google Scholar]
- Kandler, O. Carbohydrate metabolism in lactic acid bacteria. Anton. Leeuw. 1983, 49, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Filya, I.; Sucu, E.; Karabulut, A. The effects of Propionibacterium acidipropionici and Lactobacillus plantarum, applied at ensiling, on the fermentation and aerobic stability of low dry matter corn and sorghum silages. J. Ind. Microbiol. Biot. 2006, 33, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, A.; Härter, C.J.; Silva, A.W.L.; Reis, R.A.; Rabelo, C.H.S. A meta-analysis examining lactic acid bacteria inoculants for maize silage: Effects on fermentation, aerobic stability, nutritive value and livestock production. Grass Forage Sci. 2019, 74, 596–612. [Google Scholar] [CrossRef]
- Comino, L.; Tabacco, E.; Righi, F.; Revello-Chion, A.; Quarantelli, A.; Borreani, G. Effects of an inoculant containing a Lactobacillus buchneri that produces ferulate-esterase on fermentation products, aerobic stability, and fiber digestibility of maize silage harvested at different stages of maturity. Anim. Feed Sci. Technol. 2014, 198, 94–106. [Google Scholar] [CrossRef]
- Blajman, J.E.; Paez, R.B.; Vinderola, C.G.; Lingua, M.S.; Signorini, M.L. A meta-analysis on the effectiveness of homofermentative and heterofermentative lactic acid bacteria for corn silage. J. Appl. Microbiol. 2018, 125, 1655–1669. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, Z.G.; Ashbell, G.; Hen, Y.; Azrieli, A.; Szakacs, G.; Filya, I. Ensiling whole-crop wheat and corn in large containers with Lactobacillus plantarum and Lactobacillus buchneri. J. Ind. Microbiol. Biot. 2002, 28, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Huisden, C.M.; Adesogan, A.T.; Kim, S.C.; Ososanya, T. Effect of applying molasses or inoculants containing homofermentative or heterofermentative bacteria at two rates on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 2009, 92, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Puntillo, M.; Gaggiotti, M.; Oteiza, J.M.; Binetti, A.; Massera, A.; Vinderola, G. Potential of Lactic Acid Bacteria Isolated From Different Forages as Silage Inoculants for Improving Fermentation Quality and Aerobic Stability. Front. Microbiol. 2020, 11, 3091. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, F.; Tabacco, E.; Piano, S.; Casale, M.; Borreani, G. Temperature during conservation in laboratory silos affects fermentation profile and aerobic stability of corn silage treated with Lactobacillus buchneri, Lactobacillus hilgardii, and their combination. J. Dairy Sci. 2021, 104, 1696–1713. [Google Scholar] [CrossRef] [PubMed]
- Mari, L.J.; Schmidt, R.J.; Nussio, L.G.; Hallada, C.M.; Kung, L., Jr. Short communication: An evaluation of the effectiveness of Lactobacillus buchneri 40788 to alter fermentation and improve the aerobic stability of corn silage in farm silos. J. Dairy Sci. 2009, 92, 1174–1176. [Google Scholar] [CrossRef] [Green Version]
- Borreani, G.; Tabacco, E. Temperature measurements of large scale silo face to assess aerobic deterioration of corn silage on farm. In Proceedings of the International Symposium on Forage Quality and Conservation, Sao Pedro, SP, Brazil, 28–30 November 2009; FEALQ: Piracicaba, SP, Brazil, 2009. [Google Scholar]
- Borreani, G.; Tabacco, E. The relationship of silage temperature with the microbiological status of the face of corn silage bunkers. J. Dairy Sci. 2010, 93, 2620–2629. [Google Scholar] [CrossRef]
- Queiroz, O.C.M.; Adesogan, A.T.; Arriola, K.G.; Queiroz, M.F.S. Effect of a dual-purpose inoculant on the quality and nutrient losses from corn silage produced in farm-scale silos. J. Dairy Sci. 2012, 95, 3354–3362. [Google Scholar] [CrossRef]
- Gardner, N.J.; Savard, T.; Obermeier, P.; Caldwell, G.; Champagne, C.P. Selection and characterization of mixed starter cultures for lactic acid fermentation of carrot, cabbage, beet and onion vegetable mixtures. Int. J. Food Microbiol. 2001, 64, 261–275. [Google Scholar] [CrossRef]
- Schnürer, J.; Magnusson, J. Antifungal lactic acid bacteria as biopreservatives. Trends Food Sci. Technol. 2005, 16, 70–78. [Google Scholar] [CrossRef]
- Giraffa, G.; Chanishvili, N.; Widyastuti, Y. Importance of lactobacilli in food and feed biotechnology. Res. Microbiol. 2010, 161, 480–487. [Google Scholar] [CrossRef]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef] [PubMed]
- Mickan, F.J.; Piltz, J.W. Silage Storage. In Successful Silage (TopFodder Silage Manual), 2nd ed.; Kaiser, A., Piltz, J.W., Burns, H., Griffths, N., Eds.; Dairy Australia and New South Wales Department of Primary Industry: New South Wales, Australia, 2003; pp. 217–252. [Google Scholar]
- Junga, P.; Trávníček, P. Surface temperature of the exposed silo face as quick indicator of the decomposition process of maize silage. J. Cent. Eur. Agric. 2015, 16, 76–91. [Google Scholar] [CrossRef]
- Kleinschmit, D.H.; Kung, L., Jr. A meta-analysis of the effects of Lactobacillus buchneri on the fermentation and aerobic stability of corn and grass and small-grain silages. J. Dairy Sci. 2006, 89, 4005–4013. [Google Scholar] [CrossRef]
- Kung, L., Jr.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Elferink, S.J.O.; Krooneman, J.; Gottschal, J.C.; Spoelstra, S.F.; Faber, F.; Driehuis, F. Anaerobic conversion of lactic acid to acetic acid and 1,2-propanediol by Lactobacillus buchneri. Appl. Environ. Microb. 2001, 67, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Reich, L.J.; Kung, L., Jr. Effects of combining Lactobacillus buchneri 40788 with various lactic acid bacteria on the fermentation and aerobic stability of corn silage. Anim. Feed Sci. Technol. 2010, 159, 105–109. [Google Scholar] [CrossRef]
- Krooneman, J.; Faber, F.; Alderkamp, A.C.; Elferink, S.J.H.W.O.; Driehuis, F.; Cleenwerck, I.; Swings, J.; Gottschal, J.C.; Vancanneyt, M. Lactobacillus diolivorans sp. nov., a 1,2-propanediol-degrading bacterium isolated from aerobically stable maize silage. Int. J. Syst. Evol. Microbiol. 2002, 52, 639–646. [Google Scholar] [CrossRef]
- D’Amours, L.; Savoie, P. Density profile of corn silage in bunker silos. Can. Biosyst. Eng. 2005, 47, 2–21. [Google Scholar] [CrossRef]
- Whiter, A.G.; Kung, L., Jr. The effect of a dry or liquid application of Lactobacillus plantarum MTD1 on the fermentation of alfalfa silage. J. Dairy Sci. 2001, 84, 2195–2202. [Google Scholar] [CrossRef]
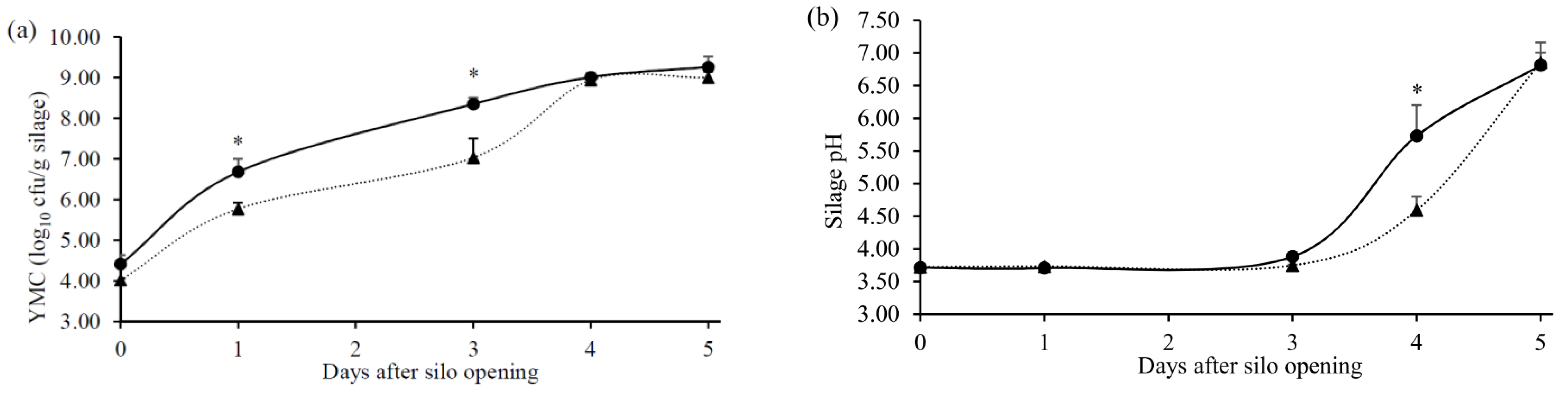
 , n = 3) or with (
, n = 3) or with (  , n = 3) DPI. Values with asterisk (*) within the same aerobic exposure time differed significantly (p < 0.05). Data were presented as mean ± SEM.
, n = 3) or with ( , n = 3) DPI. Values with asterisk (*) within the same aerobic exposure time differed significantly (p < 0.05). Data were presented as mean ± SEM.
, n = 3) DPI. Values with asterisk (*) within the same aerobic exposure time differed significantly (p < 0.05). Data were presented as mean ± SEM.
, n = 3) or with ( , n = 3) DPI. Values with asterisk (*) within the same aerobic exposure time differed significantly (p < 0.05). Data were presented as mean ± SEM. , n=3) or with ( , n=3) DPI. Data were presented as mean ± SEM.
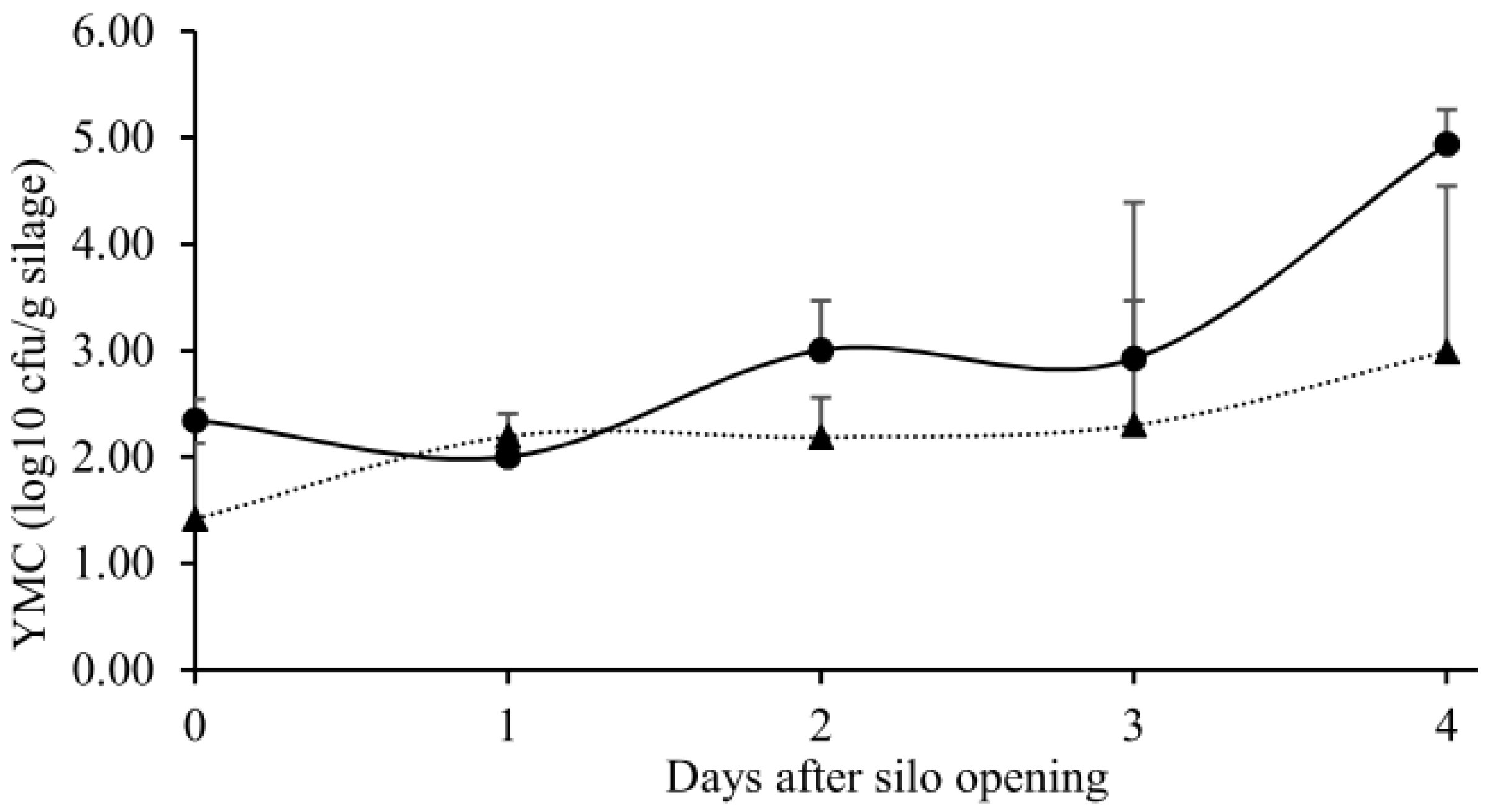
, n=3) or with ( , n=3) DPI. Data were presented as mean ± SEM.
, n=3) or with ( , n=3) DPI. Data were presented as mean ± SEM.
, n=3) or with ( , n=3) DPI. Data were presented as mean ± SEM. , n = 3) or with ( , n = 3) DPI. Asterisk (*) indicated that the slopes of linear regression lines differed significantly (p < 0.05). Data were presented as mean ± SEM. SEYX: standard error of regression coefficient.
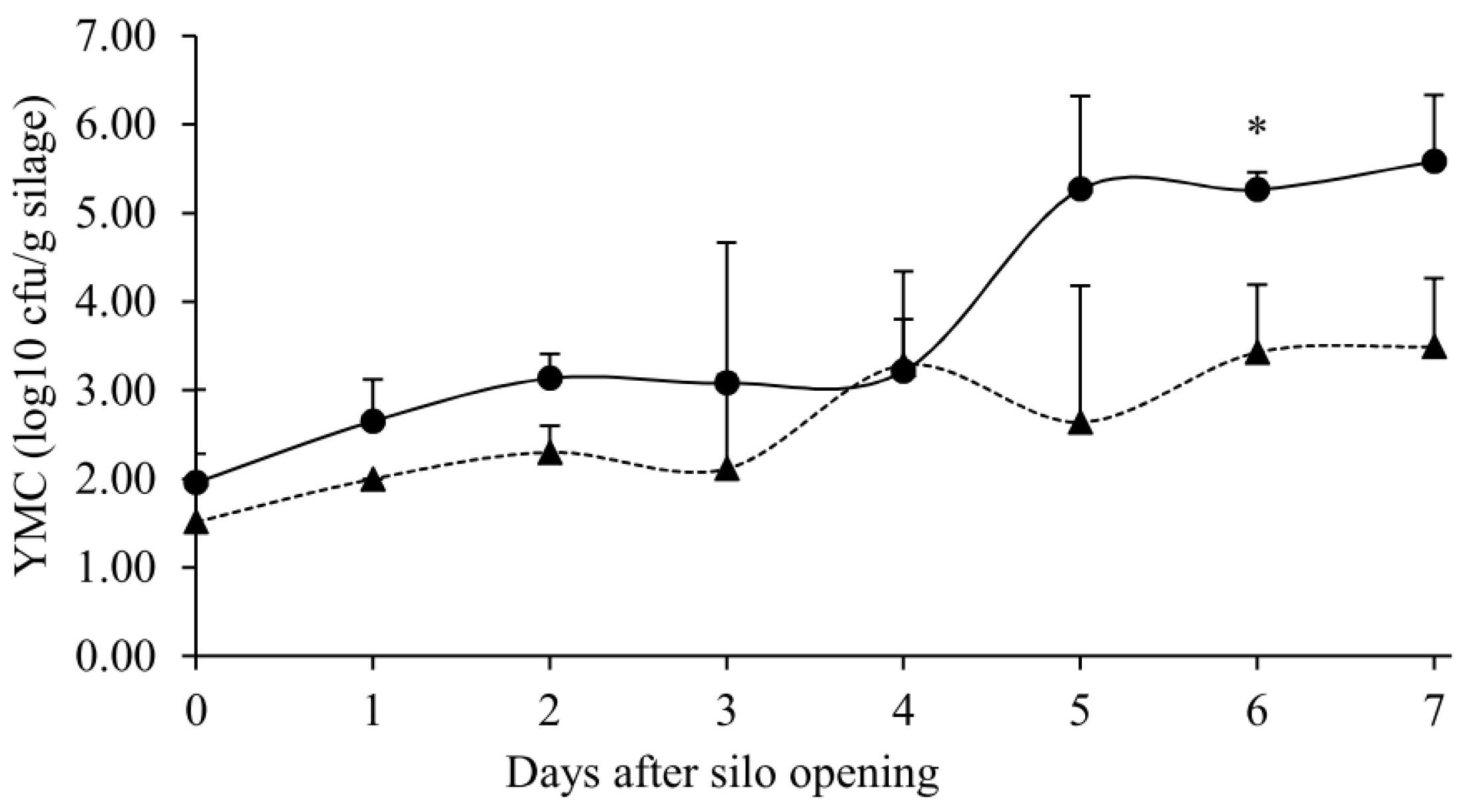
, n = 3) or with ( , n = 3) DPI. Asterisk (*) indicated that the slopes of linear regression lines differed significantly (p < 0.05). Data were presented as mean ± SEM. SEYX: standard error of regression coefficient.
, n = 3) or with ( , n = 3) DPI. Asterisk (*) indicated that the slopes of linear regression lines differed significantly (p < 0.05). Data were presented as mean ± SEM. SEYX: standard error of regression coefficient.
, n = 3) or with ( , n = 3) DPI. Asterisk (*) indicated that the slopes of linear regression lines differed significantly (p < 0.05). Data were presented as mean ± SEM. SEYX: standard error of regression coefficient. , n = 3) or with ( , n = 3) DPI. Values with asterisk (*) within the same aerobic exposure time differed significantly (p < 0.05). Data were presented as mean ± SEM.
, n = 3) or with ( , n = 3) DPI. Values with asterisk (*) within the same aerobic exposure time differed significantly (p < 0.05). Data were presented as mean ± SEM.
, n = 3) or with ( , n = 3) DPI. Values with asterisk (*) within the same aerobic exposure time differed significantly (p < 0.05). Data were presented as mean ± SEM.
, n = 3) or with ( , n = 3) DPI. Values with asterisk (*) within the same aerobic exposure time differed significantly (p < 0.05). Data were presented as mean ± SEM.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Composition 2 | 45 d | 105 d | ||||
|---|---|---|---|---|---|---|
| Control | DPI | p Value | Control | DPI | p Value | |
| DM (%) | 29.9 ± 3.2 | 24.9 ± 0.3 | 0.102 | 23.9 ± 0.3 | 24.9 ± 0.5 | 0.092 |
| CP (%) | 6.54 ± 0.10 | 6.80 ± 0.08 | 0.054 | 8.19 ± 0.16 | 9.81 ± 1.51 | 0.17 |
| ADF (%) | 23.1 ± 1.1 b | 28.7 ± 0.6 a | 0.006 | 25.9 ± 1.1 | 26.7 ± 0.9 | 0.31 |
| NDF (%) | 42.1 ± 3.0 b | 50.7 ± 1.2 a | 0.028 | 45.7 ± 1.1 | 47.4 ± 0.7 | 0.11 |
| WSC (%) | 1.7 ± 0.3 | 1.6 ± 0.2 | 0.336 | 2.23 ± 0.4 | 3.8 ± 1.4 | 0.18 |
| Starch (%) | 34.69 ± 2.77 a | 25.55 ± 1.81 b | 0.025 | 28.59 ± 1.49 | 25.88 ± 0.96 | 0.10 |
| Ammonia nitrogen (% of DM) | 3.08 ± 1.00 | 2.67 ± 0.74 | 0.381 | 2.54 ± 0.08 | 2.30 ± 0.09 | 0.057 |
| pH | 3.71 ± 0.02 | 3.72 ± 0.01 | 0.388 | 3.72 ± 0.02 | 3.72 ± 0.01 | 0.37 |
| Lactic acid (% of DM) | 3.58 ± 0.36 | 3.33 ± 0.43 | 0.337 | 3.49 ± 0.28 | 3.56 ± 0.41 | 0.45 |
| Acetic acid (% of DM) | 1.63 ± 0.15 | 1.73 ± 0.20 | 0.358 | 1.79 ± 0.25 | 2.05 ± 0.12 | 0.19 |
| LA-AA ratio | 2.19 ± 0.06 a | 1.92 ± 0.06 b | 0.014 | 2.00 ± 0.24 | 1.73 ± 0.13 | 0.18 |
| Propylene glycol (% of DM) | 0.39 ± 0.05 b | 0.51 ± 0.02 a | 0.04 | 0.58 ± 0.08 | 0.63 ± 0.04 | 0.31 |
| Butyrate (% of DM) | ND | ND | ND | ND | ||
| LAB (log10 cfu/g) | 6.1 ± 0.2 | 6.7 ± 0.4 | 0.088 | 6.4 ± 0.6 | 5.9 ± 0.5 | 0.26 |
| YMC (log10 cfu/g) | 2.94 ± 0.63 | 2.91 ± 0.13 | 0.487 | 4.42 ± 0.21 | 4.02 ± 0.03 | 0.069 |
| Aerobic stability (h) 3 | 30 | 51 | 30 | 47 | ||
| Chemical Composition 2 | 60 d | 90 d | ||||
|---|---|---|---|---|---|---|
| Control | DPI | p Value | Control | DPI | p Value | |
| DM (%) | 41.2 ± 1.2 | 39.8 ± 0.7 | 0.179 | 41.6 ± 1.3 | 42.6 ± 1.7 | 0.34 |
| CP (%) | 9.3 ± 0.2 | 9.4 ± 0.1 | 0.229 | 9.1 ± 0.2 | 8.9 ± 0.2 | 0.27 |
| ADF (%) | 25.6 ± 1.6 | 25.5 ± 1.4 | 0.473 | 24.0 ± 0.6 | 23.3 ± 0.6 | 0.24 |
| NDF (%) | 45.6 ± 1.0 | 44.8 ± 0.7 | 0.284 | 41.8 ± 1.2 | 43.0 ± 1.9 | 0.31 |
| WSC (%) | 2.54 ± 0.47 | 1.78 ± 0.06 | 0.091 | 2.19 ± 0.28 | 2.56 ± 0.55 | 0.29 |
| Starch (%) | 26.1 ± 2.6 | 26.8 ± 2.4 | 0.429 | 30.6 ± 2.0 | 31.7 ± 1.8 | 0.35 |
| pH | 3.78 ± 0.01 | 3.83 ± 0.06 | 0.215 | 3.65 ± 0.01 b | 3.72 ± 0.01 a | 0.004 |
| Lactic acid (% of DM) | 2.58 ± 0.13 b | 2.91 ± 0.06 a | 0.039 | 2.93 ± 0.06 | 2.61 ± 0.14 | 0.053 |
| Acetic acid (% of DM) | 1.43 ± 0.07 | 1.40 ± 0.10 | 0.389 | 1.46 ± 0.06 | 1.50 ± 0.14 | 0.39 |
| LA-AA ratio | 1.82 ± 0.16 | 2.10 ± 0.11 | 0.109 | 2.02 ± 0.05 | 1.75 ± 0.12 | 0.050 |
| Butyrate (% of DM) | ND | ND | ND | ND | ||
| Propylene glycol (% of DM) | 0.77 ± 0.10 | 0.61 ± 0.07 | 0.143 | 0.52 ± 0.09 | 0.69 ± 0.10 | 0.130 |
| LAB (log10 cfu/g) | 6.82 ± 0.27 | 6.43 ± 0.19 | 0.150 | 5.00 ± 0.00 | 5.23 ± 0.23 | 0.19 |
| YMC (log10 cfu/g) | 0.00 ± 0.00 | 0.62 ± 0.62 | 0.187 | 2.35 ± 0.20 | 1.42 ± 0.71 | 0.14 |
| Aerobic stability (h) | 185 ± 13 | 165 ± 40 | 0.327 | 137 ± 47 | 136 ± 57 | 0.49 |
| Chemical Composition 2 | 45 d | 105 d | ||||
|---|---|---|---|---|---|---|
| Control | DPI | p Value | Control | DPI | p Value | |
| DM (%) | 31.7 ± 1.2 | 31.7 ± 0.8 | 0.495 | 30.3 ± 0.9 | 30.4 ± 0.5 | 0.44 |
| CP (%) | 7.83 ± 0.20 | 7.96 ± 0.33 | 0.378 | 7.59 ± 0.03 | 7.47 ± 0.10 | 0.16 |
| ADF (%) | 28.3 ± 0.8 | 28.9 ± 0.5 | 0.289 | 29.5 ± 0.8 | 30.7 ± 0.8 | 0.18 |
| NDF (%) | 56.7 ± 0.9 | 58.0 ± 0.4 | 0.129 | 56.7 ± 0.5 | 58.8 ± 1.4 | 0.11 |
| WSC (%) | 1.86 ± 0.24 | 1.69 ± 0.17 | 0.301 | 1.02 ± 0.40 | 1.43 ± 0.40 | 0.26 |
| Starch (%) | 20.7 ± 0.9 | 20.4 ± 2.1 | 0.450 | 19.3 ± 0.7 | 17.1 ± 1.8 | 0.15 |
| pH | 3.76 ± 0.04 | 3.77 ± 0.05 | 0.423 | 3.79 ± 0.01 | 3.81 ± 0.06 | 0.42 |
| Lactic acid (% of DM) | 1.51 ± 0.23 | 1.41 ± 0.17 | 0.367 | 3.28 ± 0.14 | 2.90 ± 0.20 | 0.095 |
| Acetic acid (% of DM) | 3.79 ± 0.34 | 4.73 ± 0.36 | 0.065 | 3.43 ± 0.10 | 3.08 ± 0.22 | 0.11 |
| LA-AA ratio | 0.40 ± 0.05 | 0.30 ± 0.03 | 0.087 | 0.96 ± 0.02 | 0.96 ± 0.13 | 0.50 |
| Butyrate (% of DM) | ND | ND | ND | ND | ||
| Propylene glycol (% of DM) | 1.80 ± 0.64 | 2.83 ± 0.88 | 0.199 | 2.07 ± 0.05 | 1.62 ± 0.60 | 0.249 |
| LAB (log10 cfu/g) | 8.1 ± 0.1 | 8.2 ± 0.1 | 0.294 | 7.0 ± 0.3 | 7.6 ± 0.3 | 0.11 |
| YMC (log10 cfu/g) | 3.8 ± 1.8 | 3.8 ± 1.8 | 0.493 | 2.0 ± 1.1 | 1.5 ± 0.8 | 0.37 |
| Aerobic stability (h) | 53 ± 13 | 59 ± 14 | 0.389 | 53 ± 11 | 113 ± 29 | 0.062 |
| Hours of Aerobic Exposure | Control | DPI | SEM | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| Upper | Lower | Upper | Lower | Treatment | Layer | Treatment × Layer | ||
| 24h | 693 | 678 | 691 | 680 | 0.065 | 0.003 | <0.001 | <0.001 |
| 48h | 1352 | 1332 | 1346 | 1332 | 0.115 | <0.001 | <0.001 | <0.001 |
| 72h | 2021 | 1998 | 2010 | 1997 | 0.199 | <0.001 | <0.001 | <0.001 |
| 96h | 2721 | 2684 | 2702 | 2684 | 0.311 | <0.001 | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, H.-M.; Kao, L.-C.; Wang, S.-M.; Chen, C.-S.; Lee, T.-Y.; Chang, H.-T.; Young, S.-L.; Lin, J.-S. Effects of a Dual-Purpose Inoculant on the Quality and Aerobic Stability of Corn Silage at the Laboratory and Field Scales. Appl. Sci. 2021, 11, 8257. https://0-doi-org.brum.beds.ac.uk/10.3390/app11178257
Weng H-M, Kao L-C, Wang S-M, Chen C-S, Lee T-Y, Chang H-T, Young S-L, Lin J-S. Effects of a Dual-Purpose Inoculant on the Quality and Aerobic Stability of Corn Silage at the Laboratory and Field Scales. Applied Sciences. 2021; 11(17):8257. https://0-doi-org.brum.beds.ac.uk/10.3390/app11178257
Chicago/Turabian StyleWeng, Hsiu-Ming, Li-Chen Kao, Shu-Min Wang, Chia-Sheng Chen, Ting-Yu Lee, Hsiao-Tung Chang, San-Land Young, and Jin-Seng Lin. 2021. "Effects of a Dual-Purpose Inoculant on the Quality and Aerobic Stability of Corn Silage at the Laboratory and Field Scales" Applied Sciences 11, no. 17: 8257. https://0-doi-org.brum.beds.ac.uk/10.3390/app11178257