
Characterization of Umami Dry-Cured Ham-Derived Dipeptide Interaction with Metabotropic Glutamate Receptor (mGluR) by Molecular Docking Simulation



Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Rattus Norvegicus and Homo sapiens mGluR1s Shared Homology
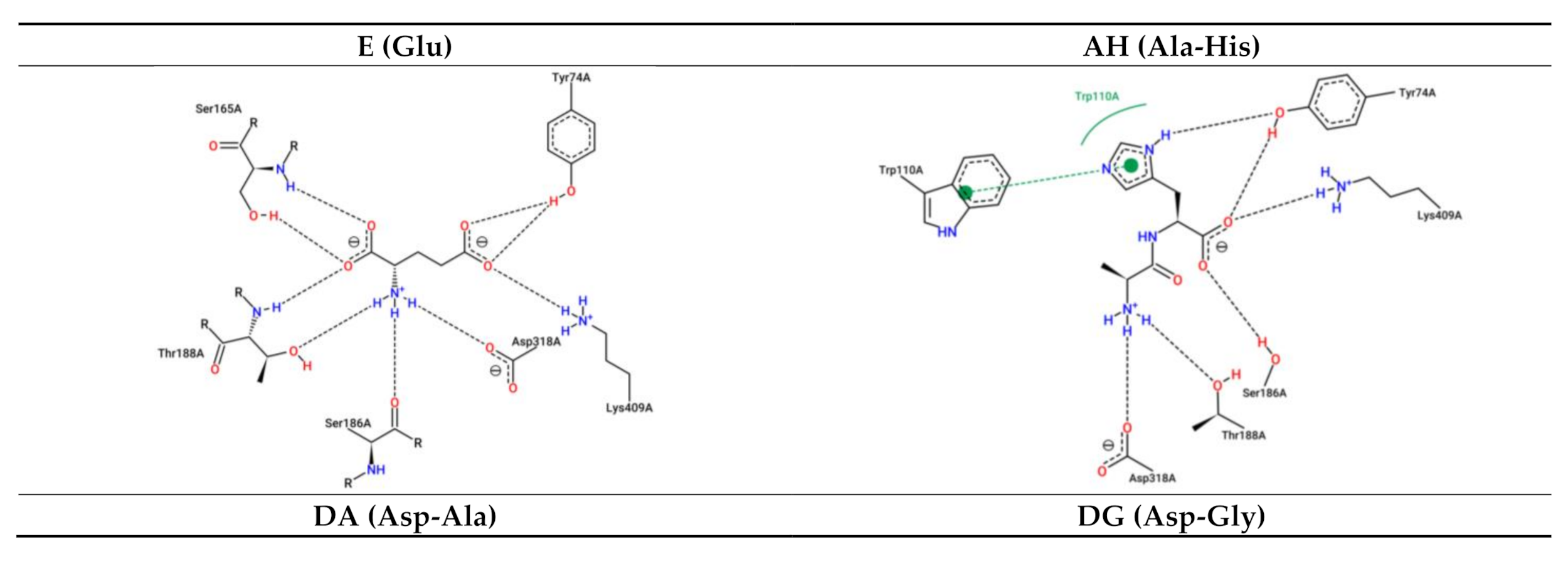
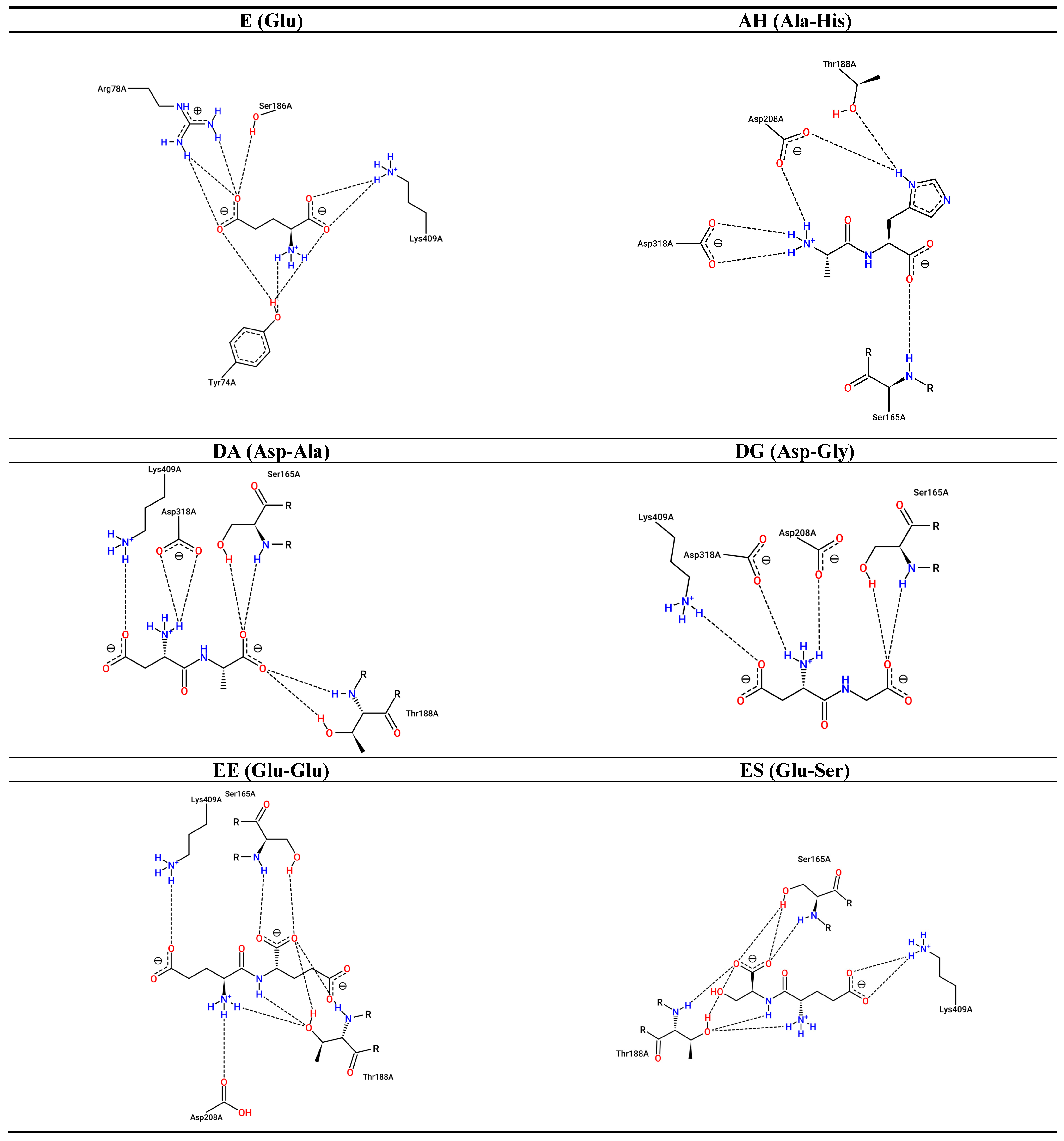
3.2. Interaction between Umami Dipeptides and Rattus Norvegicus mGluR1 Closed–Open Conformation
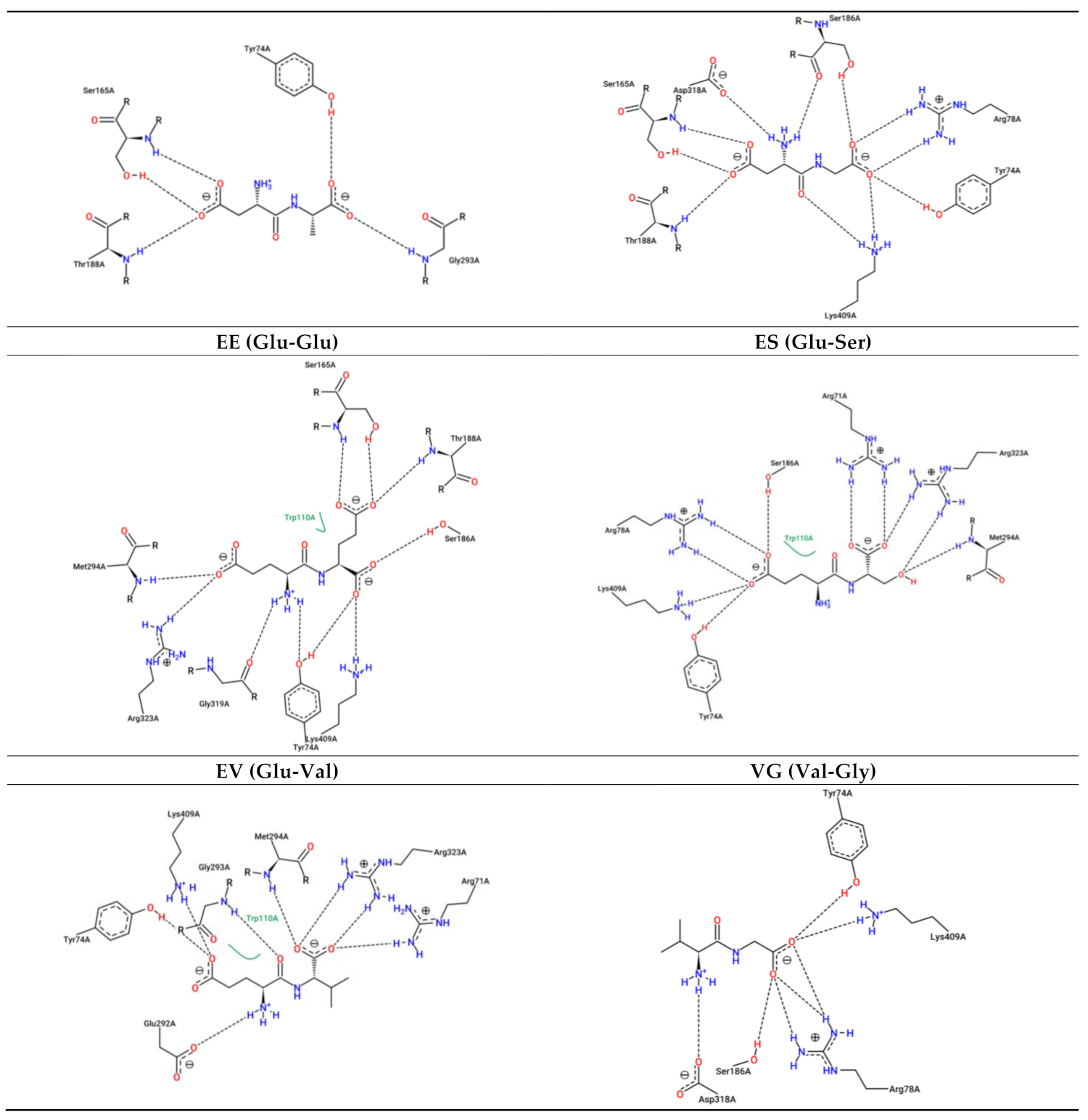
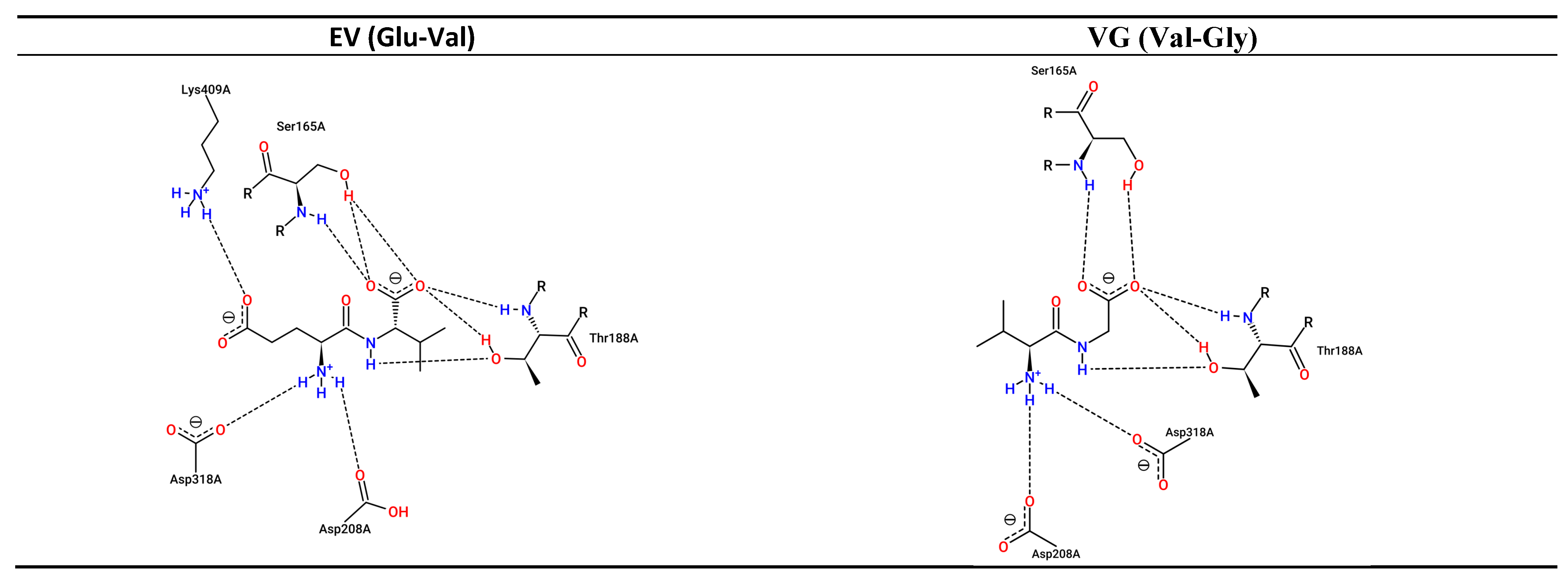
3.3. Interaction between Umami Dipeptides and Rattus Norvegicus mGluR1 Open–Open Conformation
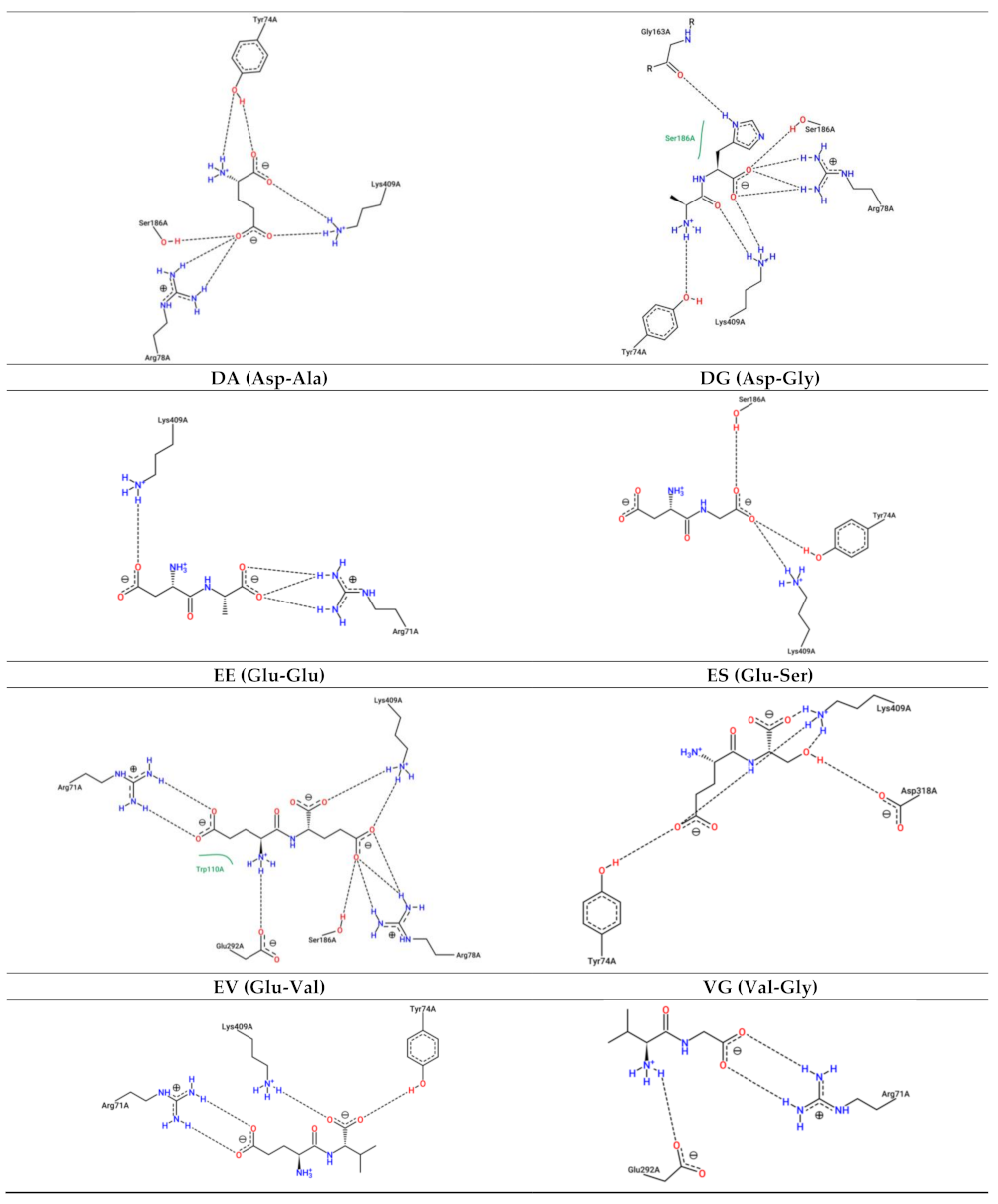
3.4. Interaction between Umami Dipeptides and Homo sapiens mGluR1 Open–Open Conformation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Santaescolástica, C.; Fraeye, I.; Barba, F.J.; Gómez, B.; Tomasevic, I.; Romero, A.; Moreno, A.; Toldrá, F.; Lorenzo, J.M. Trends in Food Science & Technology Application of Non-Invasive Technologies in Dry-Cured Ham: An Overview. Trends Food Sci. Technol. 2019, 86, 360–374. [Google Scholar] [CrossRef]
- Toldrá, F.; Aristoy, M.C. Dry-Cured Ham. In Handbook of Meat Processing; John Wiley & Sons: Hoboken, NJ, USA, 2010; pp. 351–362. [Google Scholar] [CrossRef]
- Carcò, G.; Schiavon, S.; Casiraghi, E.; Grassi, S.; Sturaro, E.; Bona, M.D.; Novelli, E.; L, G. Influence of Dietary Protein Content on the Chemico-Physical Profile of Dry-Cured Hams Produced by Pigs of Two Breeds. Sci. Rep. 2019, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bermúdez, R.; Franco, D.; Carballo, J.; Lorenzo, J.M. Physicochemical Changes during Manufacture and Final Sensory Characteristics of Dry-Cured Celta Ham. Effect of Muscle Type. Food Control 2014, 43, 263–269. [Google Scholar] [CrossRef]
- Tomažin, U.; Škrlep, M.; Prevolnik, M.; Batorek, N.; Karolyi, D. The Effect of Salting Time and Sex on Chemical and Textural Properties of Dry Cured Ham. Meat Sci. 2020, 161, 107990. [Google Scholar] [CrossRef] [PubMed]
- Čandek-Potokar, M.; Škrlep, M. Factors in Pig Production That Impact the Quality of Dry-Cured Ham: A Review. Animal 2012, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Mora, L.; Gallego, M.; Escudero, E.; Reig, M.; Aristoy, M.C.; Toldrá, F. Small Peptides Hydrolysis in Dry-Cured Meats. Int. J. Food Microbiol. 2015, 212, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Mora, L.; Bolumar, T.; Heres, A.; Toldrá, F. Effect of Cooking and Simulated Gastrointestinal Digestion on the Activity of Generated Bioactive Peptides in Aged Beef Meat. Food Funct. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F.; Flores, M.; Sanz, Y. Dry-Cured Ham Flavour: Enzymatic Generation and Process Influence. Food Chem. 1997, 59, 523–530. [Google Scholar] [CrossRef]
- Toldrá, F.; Flores, M. The Role of Muscle Proteases and Lipases in Flavor Development during the Processing of Dry-Cured Ham. Crit. Rev. Food Sci. Nutr. 1998, 38, 331–352. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F.; Gallego, M.; Reig, M.; Aristoy, M.-C.; Mora, L. Bioactive Peptides Generated in the Processing of Dry-Cured Ham. Food Chem. 2020, 321, 126689. [Google Scholar] [CrossRef]
- Aristoy, M.-C.; Toldrá, F. Isolation of Flavor Peptides from Raw Pork Meat and Dry-Cured Ham. Dev. Food Sci. 1995, 37, 1323–1344. [Google Scholar] [CrossRef]
- Sakurai, T.; Misaka, T.; Nagai, T.; Ishimaru, Y.; Matsuo, S.; Asakura, T.; Abe, K. PH-Dependent Inhibition of the Human Bitter Taste Receptor HTAS2R16 by a Variety of Acidic Substances. J. Agric. Food Chem. 2009, 57, 2508–2514. [Google Scholar] [CrossRef]
- Kim, M.J.; Son, H.J.; Kim, Y.; Misaka, T.; Rhyu, M.-R. Umami–Bitter Interactions: The Suppression of Bitterness by Umami Peptides via Human Bitter Taste Receptor. Biochem. Biophys. Res. Commun. 2015, 456, 586–590. [Google Scholar] [CrossRef]
- Sentandreu, M.A.A.; Stoeva, S.; Aristoy, M.C.C.; Laib, K.; Voelter, W.; Toldra, E.; Toldrá, F.; Toldra, E. Identification of Small Peptides Generated in Spanish Dry-Cured Ham. J. Food Sci. 2003, 68, 64–69. [Google Scholar] [CrossRef]
- Jamkhande, P.G.; Ghante, M.H.; Ajgunde, B.R. Software Based Approaches for Drug Designing and Development: A Systematic Review on Commonly Used Software and Its Applications. Bull. Fac. Pharmacy, Cairo Univ. 2017, 55, 203–210. [Google Scholar] [CrossRef]
- Pak, V.V.; Koo, M.; Kwon, D.Y.; Yun, L. Design of a Highly Potent Inhibitory Peptide Acting as a Competitive Inhibitor of HMG-CoA Reductase. Amino Acids 2012, 43, 2015–2025. [Google Scholar] [CrossRef]
- Pant, S.; Singh, M.; Ravichandiran, V.; Murty, U.S.N.; Srivastava, H.K. Peptide-like and Small-Molecule Inhibitors against Covid-19. J. Biomol. Struct. Dyn. 2021, 39, 2904–2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Qiao, L.; Chen, Y.; Zhao, B.; Gu, Y.; Huo, X.; Zhang, Y.; Li, G. In Silico Analysis of the Association Relationship between Neuroprotection and Flavors of Traditional Chinese Medicine Based on the MGluRs. Int. J. Mol. Sci. 2018, 19, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M.; Protasiewicz, M.; Mogut, D. Chemometrics and Cheminformatics in the Analysis of Biologically Active Peptides from Food Sources. J. Funct. Foods 2015, 16, 334–351. [Google Scholar] [CrossRef]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M.; Hrynkiewicz, M. Food Protein-Originating Peptides as Tastants - Physiological, Technological, Sensory, and Bioinformatic Approaches. Food Res. Int. 2016, 89, 27–38. [Google Scholar] [CrossRef]
- Finger, T.E. ATP Signaling Is Crucial for Communication from Taste Buds to Gustatory Nerves. Science 2005, 310, 1495–1499. [Google Scholar] [CrossRef]
- Yoshida, R.; Shigemura, N.; Sanematsu, K.; Yasumatsu, K.; Ishizuka, S.; Ninomiya, Y. Taste Responsiveness of Fungiform Taste Cells With Action Potentials. J. Neurophysiol. 2006, 96, 3088–3095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirazi-Beechey, S.P.; Daly, K.; Al-Rammahi, M.; Moran, A.W.; Bravo, D. Role of Nutrient-Sensing Taste 1 Receptor (T1R) Family Members in Gastrointestinal Chemosensing. Br. J. Nutr. 2014, 111, S8–S15. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, N.; Landin, A.M.; Roper, S.D. A Metabotropic Glutamate Receptor Variant Functions as a Taste Receptor. Nat. Neurosci. 2000, 3, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, A.S.; Uneyama, H.S.Y.; Torii, K. Cloning and Characterization of a Novel MGluR1 Variant from Vallate Papillae That Functions as a Receptor for L-Glutamate Stimuli. Chem. Senses 2005, 30 (Suppl. 1), i25–i26. [Google Scholar] [CrossRef] [PubMed]
- San Gabriel, A.M. Taste Receptors in the Gastrointestinal System. Flavour 2015, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Nelson, G.; Chandrashekar, J.; Hoon, M.A.; Feng, L.; Zhao, G.; Ryba, N.J.P.; Zuker, C.S. An Amino-Acid Taste Receptor. Nature 2002, 416, 199–202. [Google Scholar] [CrossRef]
- Kusuhara, Y.; Yoshida, R.; Ohkuri, T.; Yasumatsu, K.; Voigt, A.; Hübner, S.; Maeda, K.; Boehm, U.; Meyerhof, W.; Ninomiya, Y. Taste Responses in Mice Lacking Taste Receptor Subunit T1R1. J. Physiol. 2013, 591, 1967–1985. [Google Scholar] [CrossRef]
- Delay, E.R.; Hernandez, N.P.; Bromley, K.; Margolskee, R.F. Sucrose and Monosodium Glutamate Taste Thresholds and Discrimination Ability of T1R3 Knockout Mice. Chem. Senses 2006, 31, 351–357. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 Update: Improved Access to Chemical Data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef] [Green Version]
- Natesh, R.; Schwager, S.L.U.; Sturrock, E.D.; Acharya, K.R. Crystal Structure of the Human Angiotensin-Converting Enzyme–Lisinopril Complex. Nature 2003, 421, 551–554. [Google Scholar] [CrossRef] [Green Version]
- Dobrovetsky, E.; Khutoreskaya, G.; Seitova, A.; Cossar, D.; Edwards, A.M.; Arrowsmith, C.H.; Bountra, C.; Weigelt, J.; Bochkarev, A. Metabotropic Glutamate Receptor Mglur1 Complexed with LY341495 Antagonist. 2010. Unpublished. [Google Scholar]
- Berman, H.M. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A Programming Language for Software Integration and Development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar]
- Fährrolfes, R.; Bietz, S.; Flachsenberg, F.; Meyder, A.; Nittinger, E.; Otto, T.; Volkamer, A.; Rarey, M. ProteinsPlus: A Web Portal for Structure Analysis of Macromolecules. Nucleic Acids Res. 2017, 45, W337–W343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully Automated Protein–Ligand Interaction Profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef] [PubMed]
- Volkamer, A.; Kuhn, D.; Grombacher, T.; Rippmann, F.; Rarey, M. Combining Global and Local Measures for Structure-Based Druggability Predictions. J. Chem. Inf. Model. 2012, 52, 360–372. [Google Scholar] [CrossRef]
- Muchtaridi, M.; Amir, S.F.B.; Indriyati, W.; Musfiroh, I. Interaction of Aspartyl-Dipeptides Derivatives with Metabotropic Glutamate Receptor (MGluR) Using Molecular Docking Simulation. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 478–485. [Google Scholar]
- Belenikin, M.S.; Baskin, I.I.; Costantino, G.; Palyulin, V.A.; Pellicciari, R.; Zefirov, N.S. Comparative Analysis of the Ligand-Binding Sites of the Metabotropic Glutamate Receptors MGluR1–MGluR8. Dokl. Biochem. Biophys. 2002, 386, 251–256. [Google Scholar] [CrossRef]
- Bupesh, G.; Meenakumari, K.; Prabhu, J.; Prabhu, K.; Kalaiselvi, V.S.; Manikandan, E.; Krishnarao, M.R.; Sathyarajeswaran, P. Molecular Properties and Insilico Neuroprotective Activity of Eugenol Against Glutamate Metabotrophic Receptors. Int. J. Pharm. Sci. Rev. Res. 2016, 40, 318–323. [Google Scholar]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the Scope of the Protein–Ligand Interaction Profiler to DNA and RNA. Nucleic Acids Res. 2021. [Google Scholar] [CrossRef]
- Schöning-Stierand, K.; Diedrich, K.; Fährrolfes, R.; Flachsenberg, F.; Meyder, A.; Nittinger, E.; Steinegger, R.; Rarey, M. ProteinsPlus: Interactive Analysis of Protein–Ligand Binding Interfaces. Nucleic Acids Res. 2020, 48, W48–W53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toldrá, F.; Gallego, M.; Reig, M.; Aristoy, M.-C.; Mora, L. Recent Progress in Enzymatic Release of Peptides in Foods of Animal Origin and Assessment of Bioactivity. J. Agric. Food Chem. 2020, 68, 12842–12855. [Google Scholar] [CrossRef] [PubMed]
- Sentandreu, M.; Toldrá, F. Evaluation of ACE Inhibitory Activity of Dipeptides Generated by the Action of Porcine Muscle Dipeptidyl Peptidases. Food Chem. 2007, 102, 511–515. [Google Scholar] [CrossRef]
- Panchaud, A.; Affolter, M.; Kussmann, M. Mass Spectrometry for Nutritional Peptidomics: How to Analyze Food Bioactives and Their Health Effects. J. Proteomics 2012, 75, 3546–3559. [Google Scholar] [CrossRef]
- Mora, L.; Gallego, M.; Reig, M.; Toldrá, F. Challenges in the Quantitation of Naturally Generated Bioactive Peptides in Processed Meats. Trends Food Sci. Technol. 2017, 69, 306–314. [Google Scholar] [CrossRef]
- Takahashi, K.; Tokuoka, M.; Kohno, H.; Sawamura, N.; Myoken, Y.; Mizuno, A. Comprehensive Analysis of Dipeptides in Alcoholic Beverages by Tag-Based Separation and Determination Using Liquid Chromatography/Electrospray Ionization Tandem Mass Spectrometry and Quadrupole-Time-of-Flight Mass Spectrometry. J. Chromatogr. A 2012, 1242, 17–25. [Google Scholar] [CrossRef]
- Tang, Y.; Li, R.; Lin, G.; Li, L. PEP Search in MyCompoundID: Detection and Identification of Dipeptides and Tripeptides Using Dimethyl Labeling and Hydrophilic Interaction Liquid Chromatography Tandem Mass Spectrometry. Anal. Chem. 2014, 86, 3568–3574. [Google Scholar] [CrossRef]
- Dang, Y.; Gao, X.; Ma, F.; Wu, X. Comparison of Umami Taste Peptides in Water-Soluble Extractions of Jinhua and Parma Hams. LWT - Food Sci. Technol. 2015, 60, 1179–1186. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, G.M.; Zhang, J.W.; Liu, Y.X.; Li, M.Y.; Hu, D.H. Separation, Purification and Structural Identification of Small Peptides from Jinhua Ham. Food Sci. 2012, 33, 16–20. [Google Scholar] [CrossRef]
- Zhu, C.-Z.; Tian, W.; Li, M.-Y.; Liu, Y.-X.; Zhao, G.-M. Separation and Identification of Peptides from Dry-Cured Jinhua Ham. Int. J. Food Prop. 2017, 20 (Suppl. 3), S2980–S2989. [Google Scholar] [CrossRef]
- Zhou, C.-Y.; Tang, C.-B.; Wang, C.; Dai, C.; Bai, Y.; Yu, X.-B.; Li, C.-B.; Xu, X.-L.; Zhou, G.-H.; Cao, J.-X. Insights into the Evolution of Myosin Light Chain Isoforms and Its Effect on Sensory Defects of Dry-Cured Ham. Food Chem. 2020, 315, 126318. [Google Scholar] [CrossRef] [PubMed]
- Gallego, M.; Mora, L.; Aristoy, M.C.; Toldrá, F. Titin-Derived Peptides as Processing Time Markers in Dry-Cured Ham. Food Chem. 2015, 167, 326–339. [Google Scholar] [CrossRef]
- Gupta, S.; Kapoor, P.; Chaudhary, K.; Gautam, A.; Kumar, R.; Raghava, G.P.S. In Silico Approach for Predicting Toxicity of Peptides and Proteins. PLoS One 2013, 8, e73957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tharmalingam, S.; Burns, A.R.; Roy, P.J.; Hampson, D.R. Orthosteric and Allosteric Drug Binding Sites in the Caenorhabditis Elegans Mgl-2 Metabotropic Glutamate Receptor. Neuropharmacology 2012, 63, 667–674. [Google Scholar] [CrossRef]
- Crupi, R.; Impellizzeri, D.; Cuzzocrea, S. Role of Metabotropic Glutamate Receptors in Neurological Disorders. Front. Mol. Neurosci. 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- San Gabriel, A.; Maekawa, T.; Uneyama, H.; Torii, K. Metabotropic Glutamate Receptor Type 1 in Taste Tissue. Am. J. Clin. Nutr. 2009, 90, 743S–746S. [Google Scholar] [CrossRef]
- Nango, E.; Akiyama, S.; Maki-Yonekura, S.; Ashikawa, Y.; Kusakabe, Y.; Krayukhina, E.; Maruno, T.; Uchiyama, S.; Nuemket, N.; Yonekura, K.; et al. Taste Substance Binding Elicits Conformational Change of Taste Receptor T1r Heterodimer Extracellular Domains. Sci. Rep. 2016, 6, 25745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunishima, N.; Shimada, Y.; Tsuji, Y.; Sato, T.; Yamamoto, M.; Kumasaka, T.; Nakanishi, S.; Jingami, H.; Morikawa, K. Structural Basis of Glutamate Recognition by a Dimeric Metabotropic Glutamate Receptor. Nature 2000, 407, 971–977. [Google Scholar] [CrossRef]
- Stierand, K.; Maass, P.C.; Rarey, M. Molecular Complexes at a Glance: Automated Generation of Two-Dimensional Complex Diagrams. Bioinformatics 2006, 22, 1710–1716. [Google Scholar] [CrossRef]
- Chen, Q.; Ho, J.D.; Ashok, S.; Vargas, M.C.; Wang, J.; Atwell, S.; Bures, M.; Schkeryantz, J.M.; Monn, J.A.; Hao, J. Structural Basis for (S)-3,4-Dicarboxyphenylglycine (DCPG) As a Potent and Subtype Selective Agonist of the MGlu 8 Receptor. J. Med. Chem. 2018, 61, 10040–10052. [Google Scholar] [CrossRef]
- López Cascales, J.J.; Oliveira Costa, S.D.; de Groot, B.L.; Walters, D.E. Binding of Glutamate to the Umami Receptor. Biophys. Chem. 2010, 152, 139–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Da, L.-T.; Liu, Y. Understanding the Molecular Mechanism of Umami Recognition by T1R1-T1R3 Using Molecular Dynamics Simulations. Biochem. Biophys. Res. Commun. 2019, 514, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Klebansky, B.; Fine, R.M.; Liu, H.; Xu, H.; Servant, G.; Zoller, M.; Tachdjian, C.; Li, X. Molecular Mechanism of the Sweet Taste Enhancers. Proc. Natl. Acad. Sci. 2010, 107, 4752–4757. [Google Scholar] [CrossRef] [Green Version]
- Raliou, M.; Grauso, M.; Hoffmann, B.; Schlegel-Le-Poupon, C.; Nespoulous, C.; Debat, H.; Belloir, C.; Wiencis, A.; Sigoillot, M.; Preet Bano, S.; et al. Human Genetic Polymorphisms in T1R1 and T1R3 Taste Receptor Subunits Affect Their Function. Chem. Senses 2011, 36, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Kang, L.; Zhao, W.; Wu, S.; Ding, L.; Zheng, F.; Liu, J.; Li, J. Identification of Novel Umami Peptides from Myosin via Homology Modeling and Molecular Docking. Food Chem. 2021, 344, 128728. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Hao, L.; Cao, J.; Sun, Y.; Zeng, X.; Wu, Z.; Pan, D. Molecular Docking and Simulation of the Synergistic Effect between Umami Peptides, Monosodium Glutamate and Taste Receptor T1R1/T1R3. Food Chem. 2019, 271, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heres, A.; Mora, L.; Toldrá, F. Inhibition of 3-Hydroxy-3-Methyl-Glutaryl-Coenzyme A Reductase Enzyme by Dipeptides Identified in Dry-Cured Ham. Food Prod. Process. Nutr. 2021, 3, 18. [Google Scholar] [CrossRef]
- San Gabriel, A.M.; Maekawa, T.; Uneyama, H.; Yoshie, S.; Torii, K. MGluR1 in the Fundic Glands of Rat Stomach. FEBS Lett. 2007, 581, 1119–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Yin, Y.; Shu, X.G.; Li, T.; Li, F.; Tan, B.; Wu, Z.; Wu, G. Oral Administration of MSG Increases Expression of Glutamate Receptors and Transporters in the Gastrointestinal Tract of Young Piglets. Amino Acids 2013, 45, 1169–1177. [Google Scholar] [CrossRef]
- Torii, K.; Uneyama, H.; Nakamura, E. Physiological Roles of Dietary Glutamate Signaling via Gut–Brain Axis Due to Efficient Digestion and Absorption. J. Gastroenterol. 2013, 48, 442–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondoh, T.; Mallick, H.N.; Torii, K. Activation of the Gut-Brain Axis by Dietary Glutamate and Physiologic Significance in Energy Homeostasis. Am. J. Clin. Nutr. 2009, 90, 832S–837S. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dipeptide a | Dry-Cured Ham Parental Protein b | Net Charge (Value (pI)) c | Hydrophobicity d | Steric Hindrance d | Main Residue Attribute (N-residue, C-residue) c |
|---|---|---|---|---|---|
AH | Unknown | 0.1 (7.88) | −0.08 | 0.26 | aliphatic, basic |
DA | MLC1 [54] | −1 (0.69) | -0.23 | 0.64 | acidic, aliphatic |
DG | Unknown | −1 (0.68) | −0.28 | 0.72 | acidic, aliphatic |
EE | TTN [55] MLC1 [54] | −2 (0.85) | −0.62 | 0.68 | acidic, acidic |
ES | Unknown | −1 (1.01) | −0.44 | 0.60 | acidic, polar |
EV | Unknown | −1 (0.94) | −0.04 | 0.69 | acidic, aliphatic |
VG | Unknown | 0 (3.59) | 0.35 | 0.69 | aliphatic, aliphatic |
| Ligand | PubChem ID | Binding Energy (kcal/mol) | Inhibition Constant (µM) | Protein Residues Involved in H-Bond Interactions [Chain:Residue (Distance btw Donor-Acceptor) (Protein Donor/Acceptor, Residue from Side Chain)] | No. of H-Bonds | Protein Residues Involved in Hydrophobic Interactions[Chain:Residue (Distance btw Carbon Atoms)] | Protein Residues Involved in Salt Bridges [Chain:Residue (Distance btw Centers of Charge) (Ligand Functional Group Providing the Charge)] |
|---|---|---|---|---|---|---|---|
| E | 33,032 | −6.56 | 15.58 | A:Tyr74 (2.80 Å) (Donor,sd) A:Tyr74 (2.89 Å) (Acceptor,sd) A:Ser165 (3.03 Å) (Donor) A:Ser165 (2.86 Å) (Acceptor,sd) A:Ser186 (4.08 Å) (Donor,sd) A:Thr188 (2.80 Å) (Donor) A:Asp318 (3.47 Å) (Acceptor,sd) | 7 | Absent | A:Arg323 (5.39 Å) (Carboxilate) A:Lys409 (3.25 Å) (Carboxilate) |
| AH | 9,837,455 | −6.93 | 8.26 | A:Tyr74 (2.84 Å) (Donor,sd) A:Tyr74 (2.84 Å) (Acceptor,sd) A:Ser186 (2.82 Å) (Donor,sd) A:Ser186 (3.88 Å) (Acceptor,sd) A:Tyr236 (3.74 Å) (Acceptor,sd) A:Glu292 (4.02 Å) (Acceptor,sd) A:Gly293 (3.57 Å) (Donor) A:Arg323 (2.79 Å) (Donor,sd) | 8 | A:Trp110 (3.86 Å) A:Tyr236 (3.27 Å) | A:Arg78 (4.47 Å) (Carboxilate) A:Lys409 (3.49 Å) (Carboxilate) |
| DA | 5,491,963 | −6.23 | 27.24 | A:Tyr74 (2.68 Å) (Donor,sd) A:Ser165 (3.12 Å) (Donor) A:Ser165 (2.73 Å) (Acceptor,sd) A:Thr188 (3.16 Å) (Donor) A:Gly293 (3.06 Å) (Donor) A:Asp318 (3.44 Å) (Acceptor,sd) A:Gly319 (3.08 Å) (Acceptor) A:Lys409 (3.69 Å) (Donor,sd) | 8 | A:Tyr236 (3.13 Å) A:Glu292 (3.17 Å) | A:Arg323 (4.24 Å) (Carboxilate) A:Lys409 (5.38 Å) (Carboxilate) |
| DG | 151,148 | −7.12 | 6.00 | A:Tyr74 (3.13 Å) (Acceptor,sd) A:Gly163 (3.73 Å) (Acceptor) A:Ser165 (2.72 Å) (Donor) A:Ser165 (2.77 Å) (Acceptor,sd) A:Ser186 (2.82 Å) (Donor,sd) A:Thr188 (3.05 Å) (Donor) A:Asp318 (3.46 Å) (Acceptor,sd) A:Lys409 (3.90 Å) (Donor,sd) | 8 | A:Tyr236 (3.48 Å) | A:Arg78 (3.56 Å) (Carboxilate) A:Lys409 (3.97 Å) (Carboxilate) |
| EE | 439,500 | 5.77 | 59.42 | A:Tyr74 (2.97 Å) (Acceptor,sd) A:Tyr74 (3.22 Å) (Donor,sd) A:Tyr74 (3.22 Å) (Acceptor,sd) A:Ser165 (2.73 Å) (Donor) A:Ser186 (2.76 Å) (Donor,sd) A:Ser186 (2.61 Å) (Acceptor) A:Thr188 (2.99 Å) (Donor) A:Gly293 (3.41 Å) (Donor) A:Met294 (3.14 Å) (Donor) A:Gly319 (2.55 Å) (Acceptor) A:Arg323 (2.44 Å) (Donor,sd) | 11 | A:Tyr74 (3.33 Å) A:Trp110 (3.10 Å) A:Trp110 (3.51 Å) A:Glu292 (3.54 Å) | A:Arg71 (4.74 Å) (Carboxilate) A:Arg78 (4.28 Å) (Carboxilate) A:Arg323 (4.21 Å) (Carboxilate) A:Lys409 (3.60 Å) (Carboxilate) |
| ES | 6,995,653 | −6.87 | 9.17 | A:Tyr74 (3.46 Å) (Donor,sd) A:Tyr74 (3.46 Å) (Acceptor,sd) A:Tyr74 (2.80 Å) (Acceptor,sd) A:Ser166 (2.97 Å) (Donor,sd) A:Gly293 (3.68 Å) (Donor) A:Met294 (3.03 Å) (Donor) A:Asp318 (3.57 Å) (Acceptor,sd) A:Arg323 (2.94 Å) (Donor,sd) A:Arg323 (3.86 Å) (Donor,sd) | 9 | A:Trp110 (3.12 Å) A:Lys409 (3.84 Å) | A:Arg71 (3.44 Å) (Carboxilate) A:Arg78 (3.72 Å) (Carboxilate) A:Arg323 (4.04 Å) (Carboxilate) A:Lys409 (3.99 Å) (Carboxilate) |
| EV | 6,992,567 | −6.96 | 7.87 | A:Tyr74 (3.77 Å) (Acceptor,sd) A:Tyr74 (3.83 Å) (Donor,sd) A:Glu292 (3.65 Å) (Acceptor,sd) A:Glu292 (3.28 Å) (Acceptor,sd) A:Gly293 (2.85 Å) (Donor) A:Met294 (3.11 Å) (Donor) A:Asp318 (2.91 Å) (Acceptor,sd) A:Arg323 (2.55 Å) (Donor,sd) | 8 | A:Trp110 (3.48 Å) A:Glu292 (3.03 Å) A:Met294 (3.19 Å) | A:Arg71 (4.49 Å) (Carboxilate) A:Arg323 (3.19 Å) (Carboxilate) A:Lys409 (2.69 Å) (Carboxilate) |
| VG | 6,993,111 | −8.31 | 0.811 | A:Tyr74 (2.93 Å) (Acceptor,sd) A:Gly163 (3.57 Å) (Acceptor) A:Ser186 (2.72 Å) (Donor,sd) A:Thr188 (3.47 Å) (Acceptor,sd) A:Lys409 (3.96 Å) (Donor,sd) | 5 | A:Tyr236 (3.44 Å) A:Tyr236 (3.37 Å) | A:Arg78 (3.66 Å) (Carboxilate) A:Lys409 (3.75 Å) (Carboxilate) |
| Ligand | PubChem ID | Binding Energy (kcal/mol) | Inhibition Constant (µM) | Protein Residues Involved in H-Bond Interactions [Chain:Residue (Distance btw Donor-Acceptor) (Protein Donor/Acceptor, Residue from Side Chain)] | No. of H-Bonds | Protein Residues Involved in Hydrophobic Interactions [Chain:Residue (Distance btw Carbon Atoms)] | Protein Residues Involved in Salt Bridges [Chain:Residue (Distance btw Centers of Charge) (Ligand Functional Group Providing the Charge)] |
|---|---|---|---|---|---|---|---|
| E | 33,032 | −4.31 | 694.05 | A:Tyr74 (3.31 Å) (Acceptor,sd) A:Tyr74 (2.97 Å) (Acceptor,sd) A:Tyr74 (2.57 Å) (Acceptor,sd) A:Ser186 (3.00 Å) (Donor,sd) | 4 | A:Trp110 (3.30 Å) | A:Arg78 (3.91 Å) (Carboxilate) A:Lys409 (3.90 Å) (Carboxilate) A:Lys409 (2.49 Å) (Carboxilate) |
| AH | 9,837,455 | −6.70 | 12.32 | A:Tyr74 (3.40 Å) (Acceptor,sd) A:Tyr74 (2.59 Å) (Acceptor,sd) A:Gly163 (2.74 Å) (Acceptor) A:Ser165 (3.43 Å) (Donor) A:Ser186 (2.99 Å) (Donor,sd) A:Thr188 (3.46 Å) (Acceptor,sd) A:Lys409 (2.93 Å) (Donor,sd) | 7 | A:Trp110 (3.31 Å) A:Trp110 (3.48 Å) | A:Arg78 (3.85 Å) (Carboxilate) A:Lys409 (3.91 Å) (Carboxilate) |
| DA | 5,491,963 | −4.76 | 322.75 | A:Tyr74 (3.36 Å) (Acceptor,sd) A:Glu292 (3.65 Å) (Donor,sd) A:Glu292 (3.65 Å) (Acceptor,sd) | 3 | Absent | A:Arg71 (3.58 Å) (Carboxilate) A:Lys409 (2.77 Å) (Carboxilate) |
| DG | 151,148 | −4.60 | 424.75 | A:Tyr74 (2.79 Å) (Acceptor,sd) A:Tyr74 (2.38 Å) (Acceptor,sd) A:Tyr74 (3.00 Å) (Donor,sd) A:Ser186 (3.10 Å) (Donor,sd) A:Lys409 (3.76 Å) (Acceptor,sd) | 5 | Absent | A:Arg78 (4.39 Å) (Carboxilate) A:Arg323 (4.91 Å) (Carboxilate) A:Lys409 (3.28 Å) (Carboxilate) A:Lys409 (3.81 Å) (Carboxilate) |
| EE | 439,500 | −6.21 | 27.93 | A:Tyr74 (3.75 Å) (Acceptor,sd) A:Ser186 (3.02 Å) (Donor,sd) A:Glu292 (3.06 Å) (Donor,sd) A:Glu292 (3.06 Å) (Acceptor,sd) | 4 | A:Tyr74 (3.42 Å) A:Trp110 (3.16 Å) A:Trp110 (3.02 Å) | A:Arg71 (3.50 Å) (Carboxilate) A:Arg78 (3.65 Å) (Carboxilate) A:Lys409 (4.16 Å) (Carboxilate) A:Lys409 (3.40 Å) (Carboxilate) |
| ES | 6,995,653 | −4.17 | 875.75 | A:Tyr74 (2.82 Å) (Acceptor,sd) A:Tyr74 (2.64 Å) (Acceptor,sd) A:Ser186 (3.76 Å) (Donor,sd) A:Asp318 (2.83 Å) (Accpetor,sd) A:Ser408 (4.07 Å) (Donor,sd) A:Lys409 (3.00 Å) (Donor,sd) | 6 | A:Tyr74 (3.70 Å) A:Trp110 (3.30 Å) | A:Lys409 (2.99 Å) (Carboxilate) A:Lys409 (2.88 Å) (Carboxilate) |
| EV | 6,992,567 | −6.17 | 29.93 | A:Tyr74 (2.57 Å) (Acceptor,sd) A:Tyr74 (3.86 Å) (Acceptor,sd) A:Glu292 (2.53 Å) (Donor,sd) A:Glu292 (2.53 Å) (Acceptor,sd) | 4 | A:Trp110 (2.96 Å) A:Glu292 (3.64 Å) | A:Arg71 (3.44 Å) (Carboxilate) A:Lys409 (2.59 Å) (Carboxilate) |
| VG | 6,993,111 | −5.90 | 47.1 | A:Trp110 (2.96 Å) (Donor,sd) A:Glu292 (2.57 Å) (Donor,sd) A:Glu292 (2.57 Å) (Acceptor,sd) | 3 | A:Tyr236 (3.83 Å) A:Glu292 (3.92 Å) | A:Arg71 (3.50 Å) (Carboxilate) |
| Ligand | PubChem ID | Binding Energy (kcal/mol) | Inhibition Constant (µM) | Protein Residues Involved in H-Bond Interactions [Chain:Residue (Distance btw Donor-Acceptor) (Protein Donor/Acceptor, Residue from Side Chain)] | No. of H Bonds | Protein Residues Involved in Hydrophobic Interactions [Chain:Residue (Distance btw Carbon Atoms)] | Protein residues involved in Salt Bridges [Chain:Residue (Distance btw Centers of Charge) (Ligand Functional Group providing the Charge)] | π-Stacking [Chain:Residue (Distance in atm) (Stacking Type)] |
|---|---|---|---|---|---|---|---|---|
| E | 33,032 | −4.58 | 438.58 | A:Tyr74 (2.49 Å) (Acceptor,sd) A:Tyr74 (2.74 Å) (Acceptor,sd) A:Tyr74 (2.42 Å) (Acceptor,sd) A:Ser186 (3.17 Å) (Donor,sd) | 4 | A:Tyr74 (3.70 Å) A:Trp110 (3.28 Å) | A:Arg78 (3.60 Å) (Carboxilate) A:Lys409 (2.59 Å) (Carboxilate) A:Lys409 (4.74 Å) (Carboxilate) | Absent |
| AH | 9,837,455 | −6.83 | 9.93 | A:Ser165 (2.96 Å) (Donor) A:Ser186 (2.88 Å) (Acceptor) A:Thr188 (3.27 Å) (Donor,sd) A:Asn235 (2.78 Å) (Acceptor,sd) A:Tyr236 (3.53 Å) (Donor) A:Thr188 (3.27 Å) (Acceptor,sd) A:Asp208 (3.73 Å) (Acceptor,sd) A:Gln211 (3.37 Å) (Donor,sd) | 8 | A:Thr188 (3.51 Å) | Absent | A:Tyr236 (5.31 Å) (⊥) |
| DA | 5,491,963 | −5.50 | 92.95 | A:Ser165 (2.94 Å) (Donor) A:Ser186 (2.58 Å) (Acceptor) A:Ser186 (2.80 Å) (Acceptor) A:Thr188 (2.76 Å) (Donor) A:Thr188 (2.70 Å) (Acceptor,sd) A:Gln211 (3.44 Å) (Donor,sd) A:Asp318 (2.91 Å) (Acceptor,sd) | 7 | A:Thr188 (3.22 Å) | A:Lys409 (3.58 Å) (Carboxilate) | Absent |
| DG | 151,148 | −4.82 | 294.68 | A:Ser165 (2.93 Å) (Donor) A:Ser186 (3.99 Å) (Acceptor) A:Ser186 (2.70 Å) (Acceptor) A:Ser186 (2.98 Å) (Acceptor) A:Asp208 (3.83 Å) (Acceptor,sd) | 5 | A:Thr188 (3.25 Å) | A:Lys409 (3.25 Å) (Carboxilate) | Absent |
| EE | 439,500 | −4.92 | 248.83 | A:Ser165 (2.99 Å) (Donor) A:Ser186 (3.54 Å) (Donor,sd) A:Thr188 (2.68 Å) (Donor) A:Thr188 (2.87 Å) (Acceptor,sd) A:Thr188 (3.07 Å) (Acceptor,sd) A:Ser189 (3.87 Å) (Donor) A:Asp208 (3.25 Å) (Acceptor,sd) A:Asp318 (2.92 Å) (Acceptor,sd) | 8 | A:Thr188 (3.03 Å) A:Leu342 (3.10 Å) | A:Lys409 (3.10 Å) (Carboxilate) A:Lys409 (3.28 Å) (Carboxilate) | Absent |
| ES | 6,995,653 | −5.16 | 165.54 | A:Ser165 (2.91 Å) (Donor) A:Thr188 (2.78 Å) (Acceptor,sd) A:Thr188 (2.79 Å) (Donor) A:Thr188 (2.67 Å) (Acceptor,sd) A:Thr188 (2.89 Å) (Acceptor,sd) A:Ser189 (4.07 Å) (Donor) A:Gln211 (3.73 Å) (Donor,sd) A:Asp318 (3.88 Å) (Acceptor,sd) | 8 | A:Thr188 (3.40 Å) A:Leu342 (3.51 Å) | A:Lys409 (2.55 Å) (Carboxilate) | Absent |
| EV | 6,992,567 | −6.11 | 33.23 | A:Ser165 (3.02 Å) (Donor) A:Ser186 (2.89 Å) (Acceptor) A:Thr188 (2.62 Å) (Donor) A:Thr188 (2.68 Å) (Acceptor,sd) A:Thr188 (3.01 Å) (Acceptor,sd) A:Ser189 (3.86 Å) (Donor) A:Gln211 (3.87 Å) (Donor,sd) A:Asp318 (3.13 Å) (Acceptor,sd) | 8 | A:Thr188 (2.97 Å) A:Leu342 (3.52 Å) | A:Lys409 (3.04 Å) (Carboxilate) | Absent |
| VG | 6,993,111 | −6.49 | 17.44 | A:Ser165 (3.02 Å) (Donor) A:Thr188 (2.80 Å) (Donor) A:Thr188 (2.83 Å) (Acceptor,sd) A:Thr188 (2.65 Å) (Acceptor,sd) A:Gln211 (3.24 Å) (Donor,sd) A:Asp318 (2.86 Å) (Acceptor,sd) | 6 | A:Thr188 (3.28 Å) A:Leu342 (3.67 Å) | Absent | Absent |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heres, A.; Toldrá, F.; Mora, L. Characterization of Umami Dry-Cured Ham-Derived Dipeptide Interaction with Metabotropic Glutamate Receptor (mGluR) by Molecular Docking Simulation. Appl. Sci. 2021, 11, 8268. https://0-doi-org.brum.beds.ac.uk/10.3390/app11178268
Heres A, Toldrá F, Mora L. Characterization of Umami Dry-Cured Ham-Derived Dipeptide Interaction with Metabotropic Glutamate Receptor (mGluR) by Molecular Docking Simulation. Applied Sciences. 2021; 11(17):8268. https://0-doi-org.brum.beds.ac.uk/10.3390/app11178268
Chicago/Turabian StyleHeres, Alejandro, Fidel Toldrá, and Leticia Mora. 2021. "Characterization of Umami Dry-Cured Ham-Derived Dipeptide Interaction with Metabotropic Glutamate Receptor (mGluR) by Molecular Docking Simulation" Applied Sciences 11, no. 17: 8268. https://0-doi-org.brum.beds.ac.uk/10.3390/app11178268