The Use of Microalgae and Cyanobacteria in the Improvement of Agricultural Practices: A Review on Their Biofertilising, Biostimulating and Biopesticide Roles


Abstract
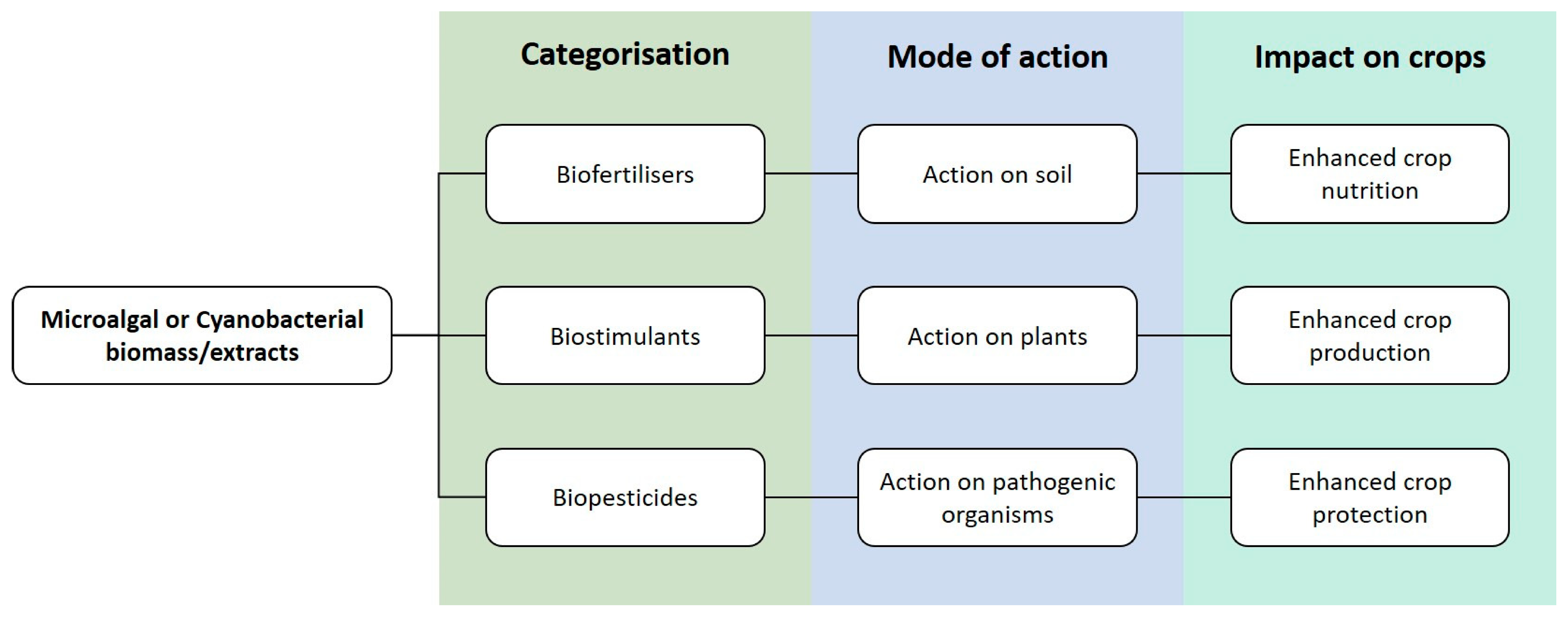
:1. Introduction
2. Microalgae as a Source of Biofertilisers, Biostimulants and Biopesticides
3. Microalgal/Cyanobacterial Metabolites and Phytohormones with Potential Interest for Agriculture
3.1. Phenolic Compounds
3.2. Terpenoids
3.3. Free Fatty Acids
3.4. Polysaccharides
3.5. Carotenoids
3.6. Phytohormones
4. The Impacts of Microalgae/Cyanobacteria and Their Extracts on Crops’ Production
4.1. Soil Improvement
4.1.1. Nitrogen Fixation
4.1.2. Nutrients’ Availability in Soils
4.1.3. Soil Physical and Chemical Amendments
4.2. Crops’ Protection
4.2.1. Protection against Biotic Factors
4.2.2. Protection against Abiotic Factors
4.3. Direct Growth Stimulation
5. Major Limitations to the Use of Microalgae and Cyanobacteria in Agriculture
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pathak, J.; Rajneesh; Maurya, P.K.; Singh, S.P.; Häder, D.-P.; Sinha, R.P. Cyanobacterial Farming for Environment Friendly Sustainable Agriculture Practices: Innovations and Perspectives. Front. Environ. Sci. 2018, 6, 7–19. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, M.; Bajguz, A.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Uncovering Potential Applications of Cyanobacteria and Algal Metabolites in Biology, Agriculture and Medicine: Current Status and Future Prospects. Front. Microbiol. 2017, 8, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiaiese, P.; Corrado, G.; Colla, G.; Kyriacou, M.C.; Rouphael, Y. Renewable Sources of Plant Biostimulation: Microalgae as a Sustainable Means to Improve Crop Performance. Front. Plant Sci. 2018, 9, 1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal Biostimulants and Biofertilisers in Crop Productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Dmytryk, A.; Chojnacka, K. Algae as fertilizers, biostimulants, and regulators of plant growth. In Algae Biomass: Characteristics and Applications; Chojnacka, K., Wieczorek, P.P., Schroeder, G., Michalak, I., Eds.; Springer: Cham, Switzerland, 2018; pp. 115–122. [Google Scholar]
- Baweja, P.; Kumar, S.; Kumar, G. Organic fertilizer from algae: A novel approach towards sustainable agriculture. In Biofertilizers for Sustainable Agriculture and Environment; Giri, B., Prasad, R., Wu, Q.-S., Varma, A., Eds.; Springer: Cham, Switzerland, 2019; pp. 353–370. [Google Scholar]
- Pan, S.; Jeevanandam, J.; Danquah, M.K. Benefits of Algal Extracts in Sustainable Agriculture. In Grand Challenges in Marine Biotechnology; Springer Science and Business Media LLC: Berlin, Germany, 2019; pp. 501–534. [Google Scholar]
- Win, T.T.; Barone, G.D.; Secundo, F.; Fu, P. Algal Biofertilizers and Plant Growth Stimulants for Sustainable Agriculture. Ind. Biotechnol. 2018, 14, 203–211. [Google Scholar] [CrossRef]
- Górka, B.; Korzeniowska, K.; Lipok, J.; Wieczorek, P.P. The Biomass of Algae and Algal Extracts in Agricultural Production. In Algae Biomass: Characteristics and Applications; Springer Science and Business Media LLC: Berlin, Germany, 2018; pp. 103–114. [Google Scholar]
- Han, X.; Zeng, H.; Bartocci, P.; Fantozzi, F.; Yan, Y. Phytohormones and Effects on Growth and Metabolites of Microalgae: A Review. Fermentation 2018, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Xu, J. Phytohormones in microalgae: A new opportunity for microalgal biotechnology? Trends Plant Sci. 2015, 20, 273–282. [Google Scholar] [CrossRef]
- Kusvuran, A.; Kusvuran, S. Using of microbial fertilizer as biostimulant alleviates damage from drought stress in guar (Cyamopsis Tetragonoloba (L.) Taub.) seedlings. Int. Lett. Nat. Sci. 2019, 76, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Reddy, C.A.; Saravanan, R.S. Polymicrobial multi-functional approach for enhancement of crop productivity. In Advances in Applied Microbiology; Sariaslani, S., Gadd, G.M., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 82, pp. 53–113. [Google Scholar]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Carvajal-Muñoz, J.; Carmona-Garcia, C. Benefits and limitations of biofertilization in agricultural practices. Livest. Res. Rural Dev. 2012, 24, 1–8. [Google Scholar]
- Esquivel-Hernández, D.A.; Ibarra-Garza, I.P.; Rodríguez-Rodríguez, J.; Cuéllar-Bermúdez, S.P.; Rostro-Alanis, M.D.J.; Alemán-Nava, G.S.; García-Pérez, J.S.; Parra-Saldívar, R. Green extraction technologies for high-value metabolites from algae: A review. Biofuels Bioprod. Biorefining 2017, 11, 215–231. [Google Scholar] [CrossRef]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Yau, S.K.; Khong, N.M.; Chan, K.W.; Ebrahimi, M. Antioxidant capacities of fucoxanthin-producing algae as influenced by their carotenoid and phenolic contents. J. Biotechnol. 2017, 241, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; De Brabanter, J.; De Cooman, L. Antioxidant potential of microalgae in relation to their phenolic and carotenoid content. Environ. Boil. Fishes 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Khoddami, A.; Wilkes, M.A.; Roberts, T.H. Techniques for Analysis of Plant Phenolic Compounds. Molecules 2013, 18, 2328–2375. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K.; Saeid, A. Plant Growth Biostimulants, Dietary Feed Supplements and Cosmetics Formulated with Supercritical CO2 Algal Extracts. Molecules 2017, 22, 66. [Google Scholar] [CrossRef]
- Oksana, S. Plant phenolic compounds for food, pharmaceutical and cosmetics production. J. Med. Plants Res. 2012, 6, 2526–2539. [Google Scholar] [CrossRef]
- Awasthi, M.; Upadhyay, A.K.; Singh, S.; Pandey, V.P.; Dwivedi, U.N. Terpenoids as promising therapeutic molecules against Alzheimer’s disease: Amyloid beta- and acetylcholinesterase-directed pharmacokinetic and molecular docking analyses. Mol. Simul. 2018, 44, 1–11. [Google Scholar] [CrossRef]
- Betterle, N.; Melis, A. Photosynthetic generation of heterologous terpenoids in cyanobacteria. Biotechnol. Bioeng. 2019, 116, 2041–2051. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef]
- Pattanaik, B.; Lindberg, P. Terpenoids and Their Biosynthesis in Cyanobacteria. Life 2015, 5, 269–293. [Google Scholar] [CrossRef] [Green Version]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-García, A.; Hosseini, S.; Martinez-Chapa, S.O.; Cordell, G.A. Multi-target Activities of Selected Alkaloids and Terpenoids. Mini-Reviews Org. Chem. 2017, 14, 272–279. [Google Scholar] [CrossRef]
- Wei, G.; Jia, Q.; Chen, X.; Köllner, T.G.; Bhattacharya, D.; Wong, G.K.; Gershenzon, J.; Chen, F. Terpene Biosynthesis in Red Algae Is Catalyzed by Microbial Type But Not Typical Plant Terpene Synthases. Plant Physiol. 2019, 179, 382–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirbas, A.; Demirbas, M.F. Importance of algae oil as a source of biodiesel. Energy Convers. Manag. 2011, 52, 163–170. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- El-Baz, F.; El-Senousy, W.; El-Sayed, A.; Kamel, M. In vitro antiviral and antimicrobial activities of Spirulina platensis extract. J. Appl. Pharm. Sci. 2013, 3, 52–56. [Google Scholar] [CrossRef]
- Feller, R.; Matos, Â.P.; Mazzutti, S.; Moecke, E.H.; Tres, M.V.; Derner, R.B.; Oliveira, J.V.; Agenor, F., Jr. Polyunsaturated ω-3 and ω-6 fatty acids, total carotenoids and antioxidant activity of three marine microalgae extracts obtained by supercritical CO2 and subcritical n-butane. J. Supercrit. Fluids 2018, 133, 437–443. [Google Scholar] [CrossRef]
- Lam, M.K.; Lee, K.T. Microalgae biofuels: A critical review of issues, problems and the way forward. Biotechnol. Adv. 2012, 30, 673–690. [Google Scholar] [CrossRef]
- El Arroussi, H.; Benhima, R.; Elbaouchi, A.; Sijilmassi, B.; El Mernissi, N.; Aafsar, A.; Meftah-Kadmiri, I.; Bendaou, N.; Smouni, A. Dunaliella salina exopolysaccharides: A promising biostimulant for salt stress tolerance in tomato (Solanum lycopersicum). Environ. Boil. Fishes 2018, 30, 2929–2941. [Google Scholar] [CrossRef]
- Campos, E.V.R.; De Oliveira, J.L.; Fraceto, L.F.; Singh, B. Polysaccharides as safer release systems for agrochemicals. Agron. Sustain. Dev. 2015, 35, 47–66. [Google Scholar] [CrossRef]
- Chanda, M.-J.; Merghoub, N.; Hicham, E.A. Microalgae polysaccharides: The new sustainable bioactive products for the development of plant bio-stimulants? World J. Microbiol. Biotechnol. 2019, 35, 177. [Google Scholar] [CrossRef] [PubMed]
- Delattre, C.; Pierre, G.; Laroche, C.; Michaud, P. Production, extraction and characterization of microalgal and cyanobacterial exopolysaccharides. Biotechnol. Adv. 2016, 34, 1159–1179. [Google Scholar] [CrossRef] [PubMed]
- Dvir, I.; Stark, A.H.; Chayoth, R.; Madar, Z.; Arad, S. Hypocholesterolemic Effects of Nutraceuticals Produced from the Red Microalga Porphyridium sp. in Rats. Nutrients 2009, 1, 156–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elarroussia, H.; Elmernissia, N.; Benhimaa, R.; El Kadmiria, I.M.; Bendaou, N.; Smouni, A.; Wahbya, I. Microalgae polysac-charides a promising plant growth biostimulant. J. Algal Biomass Util. 2016, 7, 55–63. [Google Scholar]
- Farid, R.; Mutale-Joan, C.; Redouane, B.; Najib, E.M.; Abderahime, A.; Laila, S.; Hicham, E.A. Effect of Microalgae Polysaccharides on Biochemical and Metabolomics Pathways Related to Plant Defense in Solanum lycopersicum. Appl. Biochem. Biotechnol. 2018, 188, 225–240. [Google Scholar] [CrossRef]
- González, A.; Castro, J.; Vera, J.; Moenne, A. Seaweed Oligosaccharides Stimulate Plant Growth by Enhancing Carbon and Nitrogen Assimilation, Basal Metabolism, and Cell Division. J. Plant Growth Regul. 2012, 32, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Guilherme, M.R.; Aouada, F.A.; Fajardo, A.R.; Martins, A.F.; Paulino, A.T.; Davi, M.F.; Rubira, A.F.; Muniz, E.C. Superabsorbent hydrogels based on polysaccharides for application in agriculture as soil conditioner and nutrient carrier: A review. Eur. Polym. J. 2015, 72, 365–385. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, S.; Gato, A.; Lamela, M.; Freire-Garabal, M.; Calleja, J.M. Anti-inflammatory and immunomodulatory activities of polysaccharide from Chlorella stigmatophora and Phaeodactylum tricornutum. Phytother. Res. 2003, 17, 665–670. [Google Scholar] [CrossRef]
- Rechter, S.; König, T.; Auerochs, S.; Thulke, S.; Walter, H.; Dörnenburg, H.; Walter, C.; Marschall, M. Antiviral activity of Arthrospira-derived spirulan-like substances. Antivir. Res. 2006, 72, 197–206. [Google Scholar] [CrossRef]
- Usman, A.; Khalid, S.; Usman, A.; Hussain, Z.; Wang, Y. Algal polysaccharides, novel application, and outlook. In Algae Based Polymers, Blends, and Composites; Zia, K.M., Zuber, M., Ali, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 115–153. [Google Scholar]
- Vera, J.; Castro, J.; Figueroa, A.G.; Moenne, A. Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection against Pathogens in Plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef]
- Cezare-Gomes, E.A.; Mejia-Da-Silva, L.D.C.; Pérez-Mora, L.S.; Matsudo, M.C.; Ferreira-Camargo, L.S.; Singh, A.K.; De Carvalho, J.C.M. Potential of Microalgae Carotenoids for Industrial Application. Appl. Biochem. Biotechnol. 2019, 188, 602–634. [Google Scholar] [CrossRef] [PubMed]
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from Marine Organisms: Biological Functions and Industrial Applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as Sources of Carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Zhao, Z.; Wang, Y. The effect of ryegrass and fertilizer on the petroleum contaminated soil remediation. Fresenius Environ. Bull. 2016, 25, 2243–2250. [Google Scholar]
- Rajesh, K.; Rohit, M.V.; Venkata Mohan, S. Microalgae-based carotenoids production. In Algal Green Chemistry; Rastogi, R.P., Madamwar, D., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 139–147. [Google Scholar]
- Raposo, M.F.D.J.; De Morais, R.M.S.C.; De Morais, R.M.S.C. Carotenoids from Marine Microalgae: A Valuable Natural Source for the Prevention of Chronic Diseases. Mar. Drugs 2015, 13, 5128–5155. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, Y.; Mori, K.; Matsuo, Y.; Mukojima, N.; Watanabe, W.; Sobaru, N.; Tamiya, S.; Nakao, T.; Hayashi, K.; Watanuki, H.; et al. Breeding of a new potato variety ‘Nagasaki Kogane’ with high eating quality, high carotenoid content, and resistance to diseases and pests. Breed. Sci. 2017, 67, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Ördög, V.; Stirk, W.A.; Van Staden, J.; Novák, O.; Strnad, M. Endogenous cytokinins in three genera of microalgae from the Chlorophyta. J. Phycol. 2004, 40, 88–95. [Google Scholar] [CrossRef]
- Freile-Pelegrín, Y.; Robledo, D. Bioactive phenolic compounds from algae. In Bioactive Compounds from Marine Foods: Plant and Animal Sources; Hernández-Ledesma, B., Herrero, M., Eds.; Wiley: Chichester, UK, 2013; pp. 113–129. [Google Scholar]
- Harizani, M.; Ioannou, E.; Roussis, V. The Laurencia paradox: An endless source of chemodiversity. In Progress in the Chemistry of Organic Natural Products; Kinghorn, A.D., Falk, H., Gibbons, S., Kobayashi, J.I., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 91–252. [Google Scholar]
- Hellier, P.; Al-Haj, L.; Talibi, M.; Purton, S.; Ladommatos, N. Combustion and emissions characterization of terpenes with a view to their biological production in cyanobacteria. Fuel 2013, 111, 670–688. [Google Scholar] [CrossRef] [Green Version]
- Veillette, M.; Giroir-Fendler, A.; Faucheux, N.; Heitz, M. Esterification of free fatty acids with methanol to biodiesel using heterogeneous catalysts: From model acid oil to microalgae lipids. Chem. Eng. J. 2017, 308, 101–109. [Google Scholar] [CrossRef]
- Gong, M.; Bassi, A.S. Carotenoids from microalgae: A review of recent developments. Biotechnol. Adv. 2016, 34, 1396–1412. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Z.; Sun, J.; Xue, C.; Mao, X. Biotechnological production of zeaxanthin by microorganisms. Trends Food Sci. Technol. 2018, 71, 225–234. [Google Scholar] [CrossRef]
- Kozlova, T.A.; Hardy, B.P.; Krishna, P.; Levin, D.B. Effect of phytohormones on growth and accumulation of pigments and fatty acids in the microalgae Scenedesmus quadricauda. Algal Res. 2017, 27, 325–334. [Google Scholar] [CrossRef]
- Mousavi, P.; Montazeri-Najafabady, N.; Abolhasanzadeh, Z.; Mohagheghzadeh, A.; Hamidi, M.; Niazi, A.; Morowvat, M.H.; Ghasemi, Y. Investigating the effects of phytohormones on growth and beta-carotene production in a naturally isolates stain of Dunaliella salina. J. Appl. Pharm. Sci. 2016, 6, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Piotrowska-Niczyporuk, A.; Bajguz, A. The effect of natural and synthetic auxins on the growth, metabolite content and antioxidant response of green alga Chlorella vulgaris (Trebouxiophyceae). Plant Growth Regul. 2014, 73, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Salama, E.-S.; Kabra, A.N.; Ji, M.-K.; Kim, J.R.; Min, B.; Jeon, B.-H. Enhancement of microalgae growth and fatty acid content under the influence of phytohormones. Bioresour. Technol. 2014, 172, 97–103. [Google Scholar] [CrossRef]
- Udayan, A.; Arumugam, M. Selective enrichment of Eicosapentaenoic acid (20:5n-3) in N. oceanica CASA CC201 by natural auxin supplementation. Bioresour. Technol. 2017, 242, 329–333. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; De Klerk, G.-J. Plant Growth Regulators III: Gibberellins, Ethylene, Abscisic Acid, their Analogues and Inhibitors; Miscellaneous Compounds. In Plant Propagation by Tissue Culture; Springer Science and Business Media LLC: Berlin, Germany, 2007; Volume 1, pp. 227–281. [Google Scholar]
- Maršálek, B.; Zahradníčková, H.; Hronková, M. Extracellular Abscisic Acid Produced by Cyanobacteria under Salt Stress. J. Plant Physiol. 1992, 139, 506–508. [Google Scholar] [CrossRef]
- Yoshida, K.; Igarashi, E.; Wakatsuki, E.; Miyamoto, K.; Hirata, K. Mitigation of osmotic and salt stresses by abscisic acid through reduction of stress-derived oxidative damage in Chlamydomonas reinhardtii. Plant Sci. 2004, 167, 1335–1341. [Google Scholar] [CrossRef]
- Romanenko, E.; Kosakovskaya, I.; Romanenko, P. Phytohormones of microalgae: Biological role and involvement in the regulation of physiological processes. Pt II. Cytokinins and gibberellins. Algologia 2016, 26, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Piotrowska-Niczyporuk, A.; Czerpak, R. Cellular response of light/dark-grown green alga Chlorella vulgaris Beijerinck (Chlorophyceae) to exogenous adenine- and phenylurea-type cytokinins. Acta Physiol. Plant. 2009, 31, 573–585. [Google Scholar] [CrossRef]
- Renuka, N.; Guldhe, A.; Singh, P.; Ansari, F.A.; Rawat, I.; Bux, F. Evaluating the potential of cytokinins for biomass and lipid enhancement in microalga Acutodesmus obliquus under nitrogen stress. Energy Convers. Manag. 2017, 140, 14–23. [Google Scholar] [CrossRef]
- Souza, J.M.C.; Yokoya, N.S. Effects of cytokinins on physiological and biochemical responses of the agar-producing red alga Gracilaria caudata (Gracilariales, Rhodophyta). Environ. Boil. Fishes 2016, 28, 3491–3499. [Google Scholar] [CrossRef]
- Ju, C.; Van De Poel, B.; Cooper, E.D.; Thierer, J.H.; Gibbons, T.R.; Delwiche, C.F.; Chang, C. Conservation of ethylene as a plant hormone over 450 million years of evolution. Nat. Plants 2015, 1, 14004. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.J.; Gutierrez-Wing, M.T.; Rusch, K.A.; Benton, M.G. The Effects of Plant Growth Substances and Mixed Cultures on Growth and Metabolite Production of Green Algae Chlorella sp.: A Review. J. Plant Growth Regul. 2012, 32, 417–428. [Google Scholar] [CrossRef]
- Yordanova, Z.P.; Iakimova, E.T.; Cristescu, S.M.; Harren, F.J.M.; Kapchina-Toteva, V.M.; Witkowska, E.W.I. Involvement of ethylene and nitric oxide in cell death in mastoparan-treated unicellular alga Chlamydomonas reinhardtii. Cell Biol. Int. 2010, 34, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Du, K.; Tao, H.; Wen, X.; Geng, Y.; Li, Y. Enhanced growth and lipid production of Chlorella pyrenoidosa by plant growth regulator GA3. Fresenius Environ. Bull. 2015, 24, 3414–3419. [Google Scholar]
- Falkowska, M.; Pietryczuk, A.; Piotrowska, A.; Bajguz, A.; Grygoruk, A.; Czerpak, R. The effect of gibberellic acid (GA3) on growth, metal biosorption and metabolism of the green algae Chlorella vulgaris (Chlorophyceae) Beijerinck exposed to cad-mium and lead stress. Pol. J. Environ. Stud. 2011, 20, 53–59. [Google Scholar]
- González-Garcinuño, Á.; Sánchez-Álvarez, J.M.; Galán, M.A.; Del Valle, E.M.M. Understanding and optimizing the addition of phytohormones in the culture of microalgae for lipid production. Biotechnol. Prog. 2016, 32, 1203–1211. [Google Scholar] [CrossRef]
- Pan, X.; Chang, F.; Kang, L.; Liu, Y.; Li, G.; Li, D. Effects of gibberellin A3 on growth and microcystin production in Microcystis aeruginosa (cyanophyta). J. Plant Physiol. 2008, 165, 1691–1697. [Google Scholar] [CrossRef]
- Jha, M.N.; Prasad, A.N. Efficacy of New Inexpensive Cyanobacterial Biofertilizer Including its Shelf-life. World J. Microbiol. Biotechnol. 2006, 22, 73–79. [Google Scholar] [CrossRef]
- Singh, S.; Datta, P. Outdoor evaluation of herbicide resistant strains of Anabaena variabilis as biofertilizer for rice plants. Plant Soil 2007, 296, 95–102. [Google Scholar] [CrossRef]
- Innok, S.; Chunleuchanon, S.; Boonkerd, N.; Teaumroong, N. Cyanobacterial akinete induction and its application as biofertilizer for rice cultivation. Environ. Boil. Fishes 2009, 21, 737–744. [Google Scholar] [CrossRef]
- Osman, M.E.H.; El-Sheekh, M.M.; El-Naggar, A.H.; Gheda, S.F. Effect of two species of cyanobacteria as biofertilizers on some metabolic activities, growth, and yield of pea plant. Biol. Fertil. Soils 2010, 46, 861–875. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Prasanna, R.; Kumar, A.; Pattnaik, S.; Chakravarty, K.; Shivay, Y.S.; Singh, R.; Saxena, A.K. Evaluating the influence of novel cyanobacterial biofilmed biofertilizers on soil fertility and plant nutrition in wheat. Eur. J. Soil Biol. 2013, 55, 107–116. [Google Scholar] [CrossRef]
- Prasanna, R.; Triveni, S.; Bidyarani, N.; Babu, S.; Yadav, K.; Adak, A.; Khetarpal, S.; Pal, M.; Shivay, Y.S.; Saxena, A.K. Evaluating the efficacy of cyanobacterial formulations and biofilmed inoculants for leguminous crops. Arch. Agron. Soil Sci. 2013, 60, 349–366. [Google Scholar] [CrossRef]
- Karthikeyan, N.; Prasanna, R.; Nain, L.; Kaushik, B.D. Evaluating the potential of plant growth promoting cyanobacteria as inoculants for wheat. Eur. J. Soil Biol. 2007, 43, 23–30. [Google Scholar] [CrossRef]
- Nisha, R.; Kaushik, A.; Kaushik, C. Effect of indigenous cyanobacterial application on structural stability and productivity of an organically poor semi-arid soil. Geoderma 2007, 138, 49–56. [Google Scholar] [CrossRef]
- Coppens, J.; Grunert, O.; Hende, S.V.D.; Vanhoutte, I.; Boon, N.; Haesaert, G.; De Gelder, L. The use of microalgae as a high-value organic slow-release fertilizer results in tomatoes with increased carotenoid and sugar levels. J. Appl. Phycol. 2016, 28, 2367–2377. [Google Scholar] [CrossRef]
- Manjunath, M.; Kanchan, A.; Ranjan, K.; Venkatachalam, S.; Prasanna, R.; Ramakrishnan, B.; Hossain, F.; Nain, L.; Shivay, Y.S.; Rai, A.B.; et al. Beneficial cyanobacteria and eubacteria synergistically enhance bioavailability of soil nutrients and yield of okra. Heliyon 2016, 2, e00066. [Google Scholar] [CrossRef] [Green Version]
- Renuka, N.; Prasanna, R.; Sood, A.; Ahluwalia, A.S.; Bansal, R.; Babu, S.; Singh, R.; Shivay, Y.S.; Nain, L. Exploring the efficacy of wastewater-grown microalgal biomass as a biofertilizer for wheat. Environ. Sci. Pollut. Res. 2016, 23, 6608–6620. [Google Scholar] [CrossRef]
- Renuka, N.; Prasanna, R.; Sood, A.; Bansal, R.; Bidyarani, N.; Singh, R.; Shivay, Y.S.; Nain, L.; Ahluwalia, A.S. Wastewater grown microalgal biomass as inoculants for improving micronutrient availability in wheat. Rhizosphere 2017, 3, 150–159. [Google Scholar] [CrossRef]
- Marks, E.A.; Montero, O.; Rad, J.C. The biostimulating effects of viable microalgal cells applied to a calcareous soil: Increases in bacterial biomass, phosphorus scavenging, and precipitation of carbonates. Sci. Total. Environ. 2019, 692, 784–790. [Google Scholar] [CrossRef]
- Hu, C.; Liu, Y.; Song, L.; Zhang, D. Effect of desert soil algae on the stabilization of fine sands. Environ. Boil. Fishes 2002, 14, 281–292. [Google Scholar] [CrossRef]
- Issa, O.M.; Défarge, C.; Le Bissonnais, Y.; Marin, B.; Duval, O.; Bruand, A.; D’Acqui, L.P.; Nordenberg, S.; Annerman, M. Effects of the inoculation of cyanobacteria on the microstructure and the structural stability of a tropical soil. Plant Soil 2007, 290, 209–219. [Google Scholar] [CrossRef] [Green Version]
- DeCesaro, A.; Rampel, A.; Machado, T.S.; Thomé, A.; Reddy, K.; Margarites, A.C.; Colla, L.M. Bioremediation of Soil Contaminated with Diesel and Biodiesel Fuel Using Biostimulation with Microalgae Biomass. J. Environ. Eng. 2017, 143, 04016091. [Google Scholar] [CrossRef]
- Renuka, N.; Guldhe, A.; Prasanna, R.; Singh, P.; Bux, F. Microalgae as multi-functional options in modern agriculture: Current trends, prospects and challenges. Biotechnol. Adv. 2018, 36, 1255–1273. [Google Scholar] [CrossRef]
- Sharma, R.; Khokhar, M.; Jat, R.; Khandelwal, S. Role of algae and cyanobacteria in sustainable agriculture system. Wud Pecker J. Agric. Res. 2012, 1, 381–388. [Google Scholar]
- Priya, M.; Gurung, N.; Mukherjee, K.; Bose, S. Microalgae in removal of heavy metal and organic pollutants from soil. In Microbial Biodegradation and Bioremediation; Das, S., Ed.; Elsevier: Oxford, UK, 2014; pp. 519–537. [Google Scholar]
- Chatterjee, A.; Singh, S.; Agrawal, C.; Yadav, S.; Rai, R.; Rai, L. Role of Algae as a Biofertilizer. In Algal Green Chemistry; Elsevier BV: Amsterdam, The Netherlands, 2017; pp. 189–200. [Google Scholar]
- Kumar, K.S.; Dahms, H.-U.; Won, E.-J.; Lee, J.-S.; Shin, K.-H. Microalgae—A promising tool for heavy metal remediation. Ecotoxicology and Environmental Safety 2015, 113, 329–352. [Google Scholar] [CrossRef]
- Chatzissavvidis, C.; Therios, I. Role of algae in agriculture. In Seaweeds: Agricultural Uses, Biological and Antioxidant Agents; Pomin, V.H., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2014; pp. 1–37. [Google Scholar]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bézier, A.; Lambert, B.; Joubert, J.-M.; Pugin, A. Laminarin Elicits Defense Responses in Grapevine and Induces Protection Against Botrytis cinerea and Plasmopara viticola. Mol. Plant Microbe Interact. 2003, 16, 1118–1128. [Google Scholar] [CrossRef] [Green Version]
- Arunkumar, K.; Selvapalam, N.; Rengasamy, R. The antibacterial compound sulphoglycerolipid 1-0 palmitoyl-3-0(6′-sulpho-α-quinovopyranosyl)-glycerol from Sargassum wightii Greville (Phaeophyceae). Bot. Mar. 2005, 48, 441–445. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-D. Screening of Cyanobacteria (Blue-Green algae) from Rice Paddy Soil for Antifungal Activity against Plant Pathogenic Fungi. Mycobiology 2006, 34, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.; Kim, Y.H.; Kim, S.; Kim, H. Observations on the suppression of root-knot nematode (Meloidogyne arenaria) on tomato by incorporation of cyanobacterial powder (Oscillatoria chlorina) into potting field soil. Bioresour. Technol. 2007, 98, 69–73. [Google Scholar] [CrossRef]
- Jaulneau, V.; Lafitte, C.; Corio-Costet, M.-F.; Stadnik, M.J.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.-T.; Dumas, B. An Ulva armoricana extract protects plants against three powdery mildew pathogens. Eur. J. Plant Pathol. 2011, 131, 393–401. [Google Scholar] [CrossRef]
- Chaudhary, V.; Prasanna, R.; Nain, L.; Dubey, S.C.; Gupta, V.; Singh, R.; Jaggi, S.; Bhatnagar, A.K. Bioefficacy of novel cyanobacteria-amended formulations in suppressing damping off disease in tomato seedlings. World J. Microbiol. Biotechnol. 2012, 28, 3301–3310. [Google Scholar] [CrossRef]
- Baloch, G.N.; Tariq, S.; Ehteshamul-Haque, S.; Athar, M.; Sultana, V.; Ara, J. Management of root diseases of eggplant and watermelon with the application of asafoetida and seaweeds. J. Appl. Bot. Food Qual. 2013, 86, 138–142. [Google Scholar] [CrossRef]
- Kumar, M.; Prasanna, R.; Bidyarani, N.; Babu, S.; Mishra, B.K.; Kumar, A.; Adak, A.; Jauhari, S.; Yadav, K.; Singh, R.; et al. Evaluating the plant growth promoting ability of thermotolerant bacteria and cyanobacteria and their interactions with seed spice crops. Sci. Hortic. 2013, 164, 94–101. [Google Scholar] [CrossRef]
- Babu, S.; Prasanna, R.; Bidyarani, N.; Singh, R. Analysing the colonisation of inoculated cyanobacteria in wheat plants using biochemical and molecular tools. Environ. Boil. Fishes 2014, 27, 327–338. [Google Scholar] [CrossRef]
- Priya, H.; Prasanna, R.; Ramakrishnan, B.; Bidyarani, N.; Babu, S.; Thapa, S.; Renuka, N. Influence of cyanobacterial inoculation on the culturable microbiome and growth of rice. Microbiol. Res. 2015, 171, 78–89. [Google Scholar] [CrossRef]
- Esserti, S.; Smaili, A.; Rifai, L.A.; Koussa, T.; Makroum, K.; Belfaiza, M.; Kabil, E.M.; Faize, L.; Burgos, L.; Alburquerque, N.; et al. Protective effect of three brown seaweed extracts against fungal and bacterial diseases of tomato. Environ. Boil. Fishes 2016, 29, 1081–1093. [Google Scholar] [CrossRef]
- Rodriguez, A.A.; Stella, A.M.; Storni, M.M.; Zulpa, G.; Zaccaro, M.C. Effects of cyanobacterial extracellular products and gibberellic acid on salinity tolerance in Oryza sativa L. Saline Syst. 2006, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- Sarhan, T.Z.; Ismael, S.F. Effect of Low Temperature and Seaweed Extracts on Flowering and Yield of Two Cucumber Cultivars (Cucumis sativus L.). Int. J. Agric. Food Res. 2014, 3, 41–54. [Google Scholar] [CrossRef]
- Stamatiadis, S.; Evangelou, L.; Yvin, J.-C.; Tsadilas, C.; Mina, J.M.G.; Cruz, F. Responses of winter wheat to Ascophyllum nodosum (L.) Le Jol. extract application under the effect of N fertilization and water supply. Environ. Boil. Fishes 2014, 27, 589–600. [Google Scholar] [CrossRef]
- Mansori, M.; Chernane, H.; Latique, S.; Benaliat, A.; Hsissou, D.; El Kaoua, M. Seaweed extract effect on water deficit and antioxidative mechanisms in bean plants (Phaseolus vulgaris L.). Environ. Boil. Fishes 2014, 27, 1689–1698. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Srivastava, A.K.; Saber, H.; Alwaleed, E.A.; Tran, L.-S.P. Sargassum muticum and Jania rubens regulate amino acid metabolism to improve growth and alleviate salinity in chickpea. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Haggag, W.; Hoballah, M.; Ali, R. Applications of nano biotechnological microalgae product for improve wheat productiv-ity in semai aird areas. Int. J. Agric. Technol. 2018, 14, 675–692. [Google Scholar]
- Hamed, S.M.; El-Rhman, A.A.A.; Abdel-Raouf, N.; Ibraheem, I.B. Role of marine macroalgae in plant protection & improvement for sustainable agriculture technology. Beni Suef Univ. J. Basic Appl. Sci. 2018, 7, 104–110. [Google Scholar] [CrossRef]
- Mercier, L.; Lafitte, C.; Borderies, G.; Briand, X.; Esquerré-Tugayé, M.T.; Fournier, J. The algal polysaccharide carrageenans can act as an elicitor of plant defence. New Phytol. 2001, 149, 43–51. [Google Scholar] [CrossRef]
- I Pardee, K.; Ellis, P.; Bouthillier, M.; Towers, G.H.; French, C.J. Plant virus inhibitors from marine algae. Can. J. Bot. 2004, 82, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Benkendorff, K.; Davis, A.R.; Rogers, C.N.; Bremner, J.B. Free fatty acids and sterols in the benthic spawn of aquatic molluscs, and their associated antimicrobial properties. J. Exp. Mar. Biol. Ecol. 2005, 316, 29–44. [Google Scholar] [CrossRef]
- Gerasimenko, N.I.; Martyyas, E.A.; Logvinov, S.V.; Busarova, N.G. Biological activity of lipids and photosynthetic pigments of Sargassum pallidum C. Agardh. Appl. Biochem. Microbiol. 2013, 50, 73–81. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Kaya, C.; Higgs, D.; Sakar, E. Response of two leafy vegetables grown at high salinity to supplementary potassium and phosphorus during different growth stages. J. Plant Nutr. 2002, 25, 2663–2676. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Saadatnia, H.; Riahi, H. Cyanobacteria from paddy fields in Iran as a biofertilizer in rice plants. Plant Soil Environ. 2009, 55, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Safinaz, A.; Ragaa, A. Effect of some red marine algae as biofertilizers on growth of maize (Zea mayz L.) plants. Int. Food Res. J. 2013, 20, 1629–1632. [Google Scholar]
- El Arroussi, H.; Elbaouchi, A.; Benhima, R.; Bendaou, N.; Smouni, A.; Wahby, I. Halophilic microalgae Dunaliella salina extracts improve seed germination and seedling growth of Triticum aestivum L. under salt stress. Acta Hortic. 2016, 1148, 13–26. [Google Scholar] [CrossRef]
- Omar, H.H.; Abdullatif, B.M.; Al-Kazan, M.M.; El-Gendy, A.M. Various Applications of Seaweed Improves Growth and Biochemical Constituents ofZea MaysL. andHelianthus AnnuusL. J. Plant Nutr. 2014, 38, 28–40. [Google Scholar] [CrossRef]
- Ramya, S.S.; Vijayanand, N.; Rathinavel, S. Foliar application of liquid biofertilizer of brown alga Stoechospermum marginatum on growth, biochemical and yield of Solanum melongena. Int. J. Recycl. Org. Waste Agric. 2015, 4, 167–173. [Google Scholar] [CrossRef] [Green Version]
- El-din, S.M.M.; Hassan, S.M. The promotive effect of different concentrations of marine algae on spinach plants (Spinacia oleracea L.). Egypt. J. Hortic. 2016, 43, 109–122. [Google Scholar] [CrossRef]
- Mógor, Á.F.; Ördög, V.; Lima, G.P.P.; Molnár, Z.; Mógor, G. Biostimulant properties of cyanobacterial hydrolysate related to polyamines. Environ. Boil. Fishes 2018, 30, 453–460. [Google Scholar] [CrossRef]
- Dineshkumar, R.; Subramanian, J.; Gopalsamy, J.; Jayasingam, P.; Arumugam, A.; Kannadasan, S.; Sampathkumar, P. The Impact of Using Microalgae as Biofertilizer in Maize (Zea mays L.). Waste Biomass Valorization 2017, 10, 1101–1110. [Google Scholar] [CrossRef]


{kind=link}
| Metabolites | Examples | Microalgal/Cyanobacterial Sources | Biological Activity | Role in Agriculture | Refs. |
|---|---|---|---|---|---|
| Phenolic compounds | Polyphenols; phenolic acids; flavonoids; phenylpropanoids | Botryococcus braunii; Chaetoceros calcitrans; Chlorella vulgaris; Isochrysis galbana; Isochrysis sp.; Neochloris oleoabundans; Odontella sinensis; Phaeodactylum tricornutum; Saccharina japonica; Skeletonema costatum; Tetraselmis suecica | Antibacterial; antioxidant; antifungal | Crops’ protection against pathogens or other biotic and abiotic stress conditions | [2,7,16,17,18,19,20,21] |
| Terpenoids | Hemiterpenes; monoterpenes; sesquiterpenes; diterpenes; triterpenes; polyterpenes | Chondrococcus hornemanni; Hypnea pannosa; Oscillatoria perornata; Planktothricoids raciborskii; Plocamium cornutum; Plocamium leptophyllum; Portieria hornemann; Pseudanabaena articulate; Pseudanabaena sp.; Sphaerococcus coronopifolius; Synechocystis sp.; Thermosynechococcus elongatus | Antibacterial; anticarcinogenic; antioxidant | Crops’ protection against bacteria, insects and other organisms Stimulation of preliminary growth and development of plants Attraction of pollinators | [2,7,22,23,24,25,26,27,28] |
| Free fatty acids | Saturated and unsaturated fatty acids | Anabaena; Chlorella; Dunaliella; Nannochloropsis; Porphyridium; Scenedesmus; Spirulina | Antibiotic; anticarcinogenic; antifungal; antioxidant; antiviral | Crops’ protection against pathogens or other biotic and abiotic stress conditions | [2,7,29,30,31,32,33] |
| Polysaccharides | Extracellular polysaccharides; structural polysaccharides; energy-storage polysaccharides | Aphanothece; Arthrospira; Chlamydomonas; Chlorella; Cylindrotheca; Dunaliella; Navicula; Nostoc; Phaeodactylum; Porphyridium; Rhodella; Scytonema | Antibacterial; anticancer; anticoagulant; anti-inflammatory; antioxidant | Improvement of soil quality Plant growth stimulation Crops’ protection against biotic and abiotic stress conditions | [2,7,34,35,36,37,38,39,40,41,42,43,44,45,46] |
| Carotenoids | Alpha-carotene; beta-carotene; lutein; lycopene; astaxanthin; zeaxanthin | Chlorella protothecoides; Chlorella pyrenoidosa; Chlorella zofingiensis; Dunaliella salina; Haematococcus pluvialis; Muriellopsis sp.; Phaeodactylum tricornutum; Spirulina sp. | Anticancer; anti-inflammatory; antioxidant | Soil bioremediation and fertilisation Crops’ protection against bacteria, insects and other biotic and abiotic stress conditions Crops’ fortification | [7,47,48,49,50,51,52,53] |
| Phytohormones | Auxins; abscisic acid; cytokinins; ethylene; gibberellins | Arthrospira; Chlamydomonas; Chlorella; Phormidium; Protococcus; Scenedesmus | Chemical messengers | Plant growth stimulation Regulation of cellular activities in crops Crops’ response to stress conditions | [2,4,7,10,11,54] |
| Mode of Action | Algae/Algal Extracts | Target Crop/Soil | Observed Improvements | Ref. |
|---|---|---|---|---|
| Nitrogen fixation | Cyanobacterial inoculum composed by Aulosira fertilissima, Anabaena sphaerica, Nostoc hatei, Cylindrospermum majus and Westiellopsis prolifica | Rice | Increase in nitrogen availability in the soil Increase in grain and straw yields | [80] |
| Wild type and herbicide-resistant strains of Anabaena variabilis | Rice | Increase in grain, straw and seeds yields Increase in plant height and leaf length | [81] | |
| Nostoc sp. vegetative cells | Rice | Increase in grain yields comparable to those obtained with a chemical fertiliser | [82] | |
| Nostoc entophytum and Oscillatoria angustissima | Pea | Increase in nitrogen fixation Increase in growth parameters, germination percentage and photosynthetic pigments Increase in nutritional value of pea seeds | [83] | |
| Anabaena torulosa biofilm | Wheat | Increase in nitrogen availability in the soil | [84] | |
| Cyanobacterial-bacterial biofilms including the species: Calothrix sp., Anabaena laxa, Anabaena torulosa, Anabaena doliolum, Nostoc carneum, Nostoc piscinale, Trichoderma viride, Pseudomonas fluorescens and Azotobacter chroococcum | Soybean and mungbean | Increase in nitrogen availability in the soil Increase in plant fresh weight | [85] | |
| Nutrients’ availability in soils | Calothrix ghosei, Hapalosiphon intricatus and Nostoc sp. | Wheat | Increase in organic carbon content in the soil Increase in grain yield | [86] |
| Cyanobacterial consortia including the species: Anabaena doliolum, Cylindrospermum sphaerica and Nostoc calcicola | Wheat and millet | Increase in nitrogen and phosphorus availability in the soil Decrease in soil density Improvement of water retention capacity Increase in grain yields Improvement of nutritional properties (increase in protein content of grain and leaves) | [87] | |
| Microalgal-bacterial flocs and Nannochloropsis oculata | Tomato | Increase in ammonium, phosphorus and potassium availability in the soil Improvement of fruit quality (increase in sugar and carotenoids contents) | [88] | |
| Consortia and biofilms including the species: Azotobacter sp., Anabaena sp., Providencia sp. and Calothrix sp. | Okra | Increase in zinc and iron availability in the soil Beneficial changes in the microbiome Increase in root yield and weight | [89] | |
| Microalgal-cyanobacterial unicellular and filamentous consortia including species of: Chlorella, Scenedesmus, Chlorococcum, Chroococcus, Phormidium, Anabaena, Fischerella and Spirogyra | Wheat | Increase in nitrogen, phosphorus and potassium availability in the soil Increase in organic carbon content in the soil Improvement of product quality (increase in nitrogen, phosphorus and potassium contents in roots, shoots and grains) | [90] | |
| Microalgal-cyanobacterial unicellular and filamentous consortia including species of: Chlorella, Scenedesmus, Chlorococcum, Chroococcus, Phormidium, Anabaena, Fischerella and Spirogyra | Wheat | Increase in zinc, iron, copper and manganese availability in the soil Increase in organic carbon content in the soil Increase in grain yield Improvement of plant nutritional value (increase in grain micronutrient contents) | [91] | |
| Chlorella sorokiniana | Soil from a vineyard | Increase in nitrogen availability in the soil | [92] | |
| Soil physical and chemical amendments | Microcoleus vaginatus, Phormidium tenue, Scytonema javanicum, Nostoc sp. and Desmococcus olivaceus | Unconsolidated sand | Increase in crust cohesion Increase in the resistance to wind erosion | [93] |
| Nostoc strains | Poorly aggregated tropical soils | Increase in aggregates’ stability | [94] | |
| Phycocyanin extract from Spirulina platensis and inactive biomass of Spirulina platensis | Soil contaminated with diesel and biodiesel | Reduction in diesel and biodiesel concentration using S. platensis and phycocyanin, respectively | [95] |
| Mode of Action | Algae/Algal Extracts | Target Crop | Observed Improvements | Ref. |
|---|---|---|---|---|
| Protection against biotic factors | Laminaria digitata | Grapevine | Fungicidal activity against Botrytis cinerea and Plasmopara viticola | [102] |
| Methanolic extract from Sargassum wightii | Rice | Antibacterial activity against Xanthomonas oryzae | [103] | |
| Oscillatoria, Anabaena, Nostoc, Nodularia and Calothrix species | Pepper | Fungicidal activity against Alternaria alternate and Botrytis cinerea | [104] | |
| Oscillatoria chlorina | Tomato | Fungicidal activity against Meloidogyne arenaria Improved plant growth | [105] | |
| Extracts from Ulva armoricana | Bean, grapevine and cucumber | Fungicidal activity against Erysiphe polygoni, Erysiphe necator and Sphareotheca fuliginea Reduced foliar disease | [106] | |
| Anabaena variabilis and Anabaena oscillarioides | Tomato | Reduction in fungal disease severity Improved plant growth (increased height and fresh weight of plants) | [107] | |
| Spatoglossum variabile, Stokeyia indica and Melanothamnus afaqhusainii | Watermelon and eggplant | Fungicidal activity against the root rotting fungi, Fusarium solani, Fusarium oxysporum and Macrophomina phaseolina Antinematodal activity against Meloidogyne spp. Improved plant growth (increased fresh weight of shoots, increased length of vines in watermelon and increased shoot length in eggplant) | [108] | |
| Anabaena laxa and Calothrix elenkinii | Coriander, cumin and fennel | Increase in the activity of β-1,3 endoglucanase in shoots and roots Increase in fungicidal activity Increase in germination rates Increase in shoot and root length Increase in plant dry weight | [109] | |
| Anabaena torulosa, Anabaena laxa, Anabaena azollae, Anabaena oscillarioides and Calothrix sp. | Wheat | Increase in nitrogen fixation Increase in the activity of hydrolytic and defence enzymes (endoglucanase, peroxidase, polyphenol oxidase and phenylalanine ammonia lyase) Increase in fresh and dry weight of plant | [110] | |
| Calothrix elenkinii | Rice | Increase in the activity of hydrolytic and defence enzymes (peroxidase, polyphenol oxidase and phenylalanine ammonia lyase) Increase in nitrogenase activity and indole acetic acid production Increase in plant growth | [111] | |
| Aqueous extract from Cystoseira myriophylloides and Fucus spiralis | Tomato | Antibacterial activity against Agrobacterium tumefaciens | [112] | |
| Polysaccharides’ extract from Chlorella vulgaris, Chlorella sorokiniana and Chlamydomonas reinhardtii | Tomato | Increase in the activity of hydrolytic and defence enzymes (β-1,3 endoglucanase, peroxidase, ascorbate peroxidase and phenylalanine ammonia lyase) Improvement of plant nutritional value (increase in polyunsaturated fatty acids contents) | [40] | |
| Protection against abiotic factors | Scytonema hofmanni | Rice | Alleviation of salt stress comparable to the one obtained with gibberellic acid | [113] |
| Seaweed extracts | Cucumber | Low temperature stress was not observed in treated plants Improvement of plant productivity (increase in the number of fruit per plant, fruit weight and total yield) | [114] | |
| Extracts from Ascophyllum nodosum | Wheat | Alleviation of drought stress Increase in growth, yield and nutrients uptake | [115] | |
| Extracts from Ulva rigida and Fucus spiralis | Beans | Alleviation of drought stress Increase in the activity of antioxidant enzymes and in the polyphenol content (mediators of plants’ protection mechanisms) Improvement of plant growth | [116] | |
| Sargassum muticum and Jania rubens | Chickpea | Alleviation of salt stress Increase in the activity of enzymes (superoxide dismutase and peroxidase) and amino acids (serine, threonine, proline and aspartic acid) associated with plants’ stress responses Improvement of plant growth | [117] | |
| Polysaccharides extracted from Dunaliella salina | Tomato | Alleviation of salt stress Activation of metabolic pathways involved in plants’ tolerance to stress Mitigation of the decrease in length and dry weight of shoots and roots | [34] | |
| Oscillatoria agardhii | Wheat | Alleviation of drought stress Increase in the activity of hydrolytic and defence enzymes (catalase, superoxide dismutase and peroxidase) Increase in grain yield Improvement of plant quality parameters | [118] |
| Algae/Algal Extracts | Target Crop | Observed Improvements | Ref. |
|---|---|---|---|
| Anabaena strains (Anabaena spiroides, Anabaena osillarioides, Anabaena torulosa and Anabaena variabilis) | Rice | Increase in germination rate Increase in plant height and shoots length Increase in fresh and dry weights of leaf, stem and root Improvement of soil properties (increase in soil moisture and porosity) | [127] |
| Laurencia obtusa, Corallina elongata and Jania rubens | Maize | Increase in plant length and fresh and dry weight Increase in the number of leaves Improvement of plant nutritional value (increase in potassium, phosphorus and nitrogen contents) | [128] |
| Exopolysaccharides extracts from Dunaliella salina | Wheat | Increase in germination rate and seedling growth Increase in root and coleoptiles height Increase in tolerance to salt stress | [129] |
| Aqueous extracts from Gracilaria corticata and Enteromorpha flexuosa | Maize and sunflower | Increase in shoot and root length and dry weight Improvement of plant nutritional value (increase in photosynthetic pigments, carbohydrate, proteins and nutrients contents) | [130] |
| Liquid extracts from Stoechospermum marginatum | Brinjal | Increase in shoot and root length Increase in fresh and dry weight of leaves Increase in leaf area Improvement of leaves nutritional value (increase in moisture, photosynthetic pigments, protein, amino acids, reducing sugars and ascorbic acid contents) | [131] |
| Total polysaccharides extract from Spirulina platensis | Tomato and pepper | Increase in plants’ size Increase in roots’ weight Increase in size and number of nodes | [39] |
| Ulva lactuca and Jania rubens | Spinach | Increase in plant yield and height Improvement of plant nutritional value (increase in chlorophyll and nitrogen contents) | [132] |
| Arthrospira platensis | Lettuce | Increase in seedling growth Increase in spermine content in leaves | [133] |
| Chlorella vulgaris and Spirulina platensis | Maize | Increase in germination rate and plant yield Increase in fresh and dry weights of shoot, root and whole plant Increase in shoot length and in the number of leaves | [134] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonçalves, A.L. The Use of Microalgae and Cyanobacteria in the Improvement of Agricultural Practices: A Review on Their Biofertilising, Biostimulating and Biopesticide Roles. Appl. Sci. 2021, 11, 871. https://0-doi-org.brum.beds.ac.uk/10.3390/app11020871
Gonçalves AL. The Use of Microalgae and Cyanobacteria in the Improvement of Agricultural Practices: A Review on Their Biofertilising, Biostimulating and Biopesticide Roles. Applied Sciences. 2021; 11(2):871. https://0-doi-org.brum.beds.ac.uk/10.3390/app11020871
Chicago/Turabian StyleGonçalves, Ana L. 2021. "The Use of Microalgae and Cyanobacteria in the Improvement of Agricultural Practices: A Review on Their Biofertilising, Biostimulating and Biopesticide Roles" Applied Sciences 11, no. 2: 871. https://0-doi-org.brum.beds.ac.uk/10.3390/app11020871