
Exploring Methane Emission Drivers in Wetlands: The Cases of Massaciuccoli and Porta Lakes (Northern Tuscany, Italy)

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
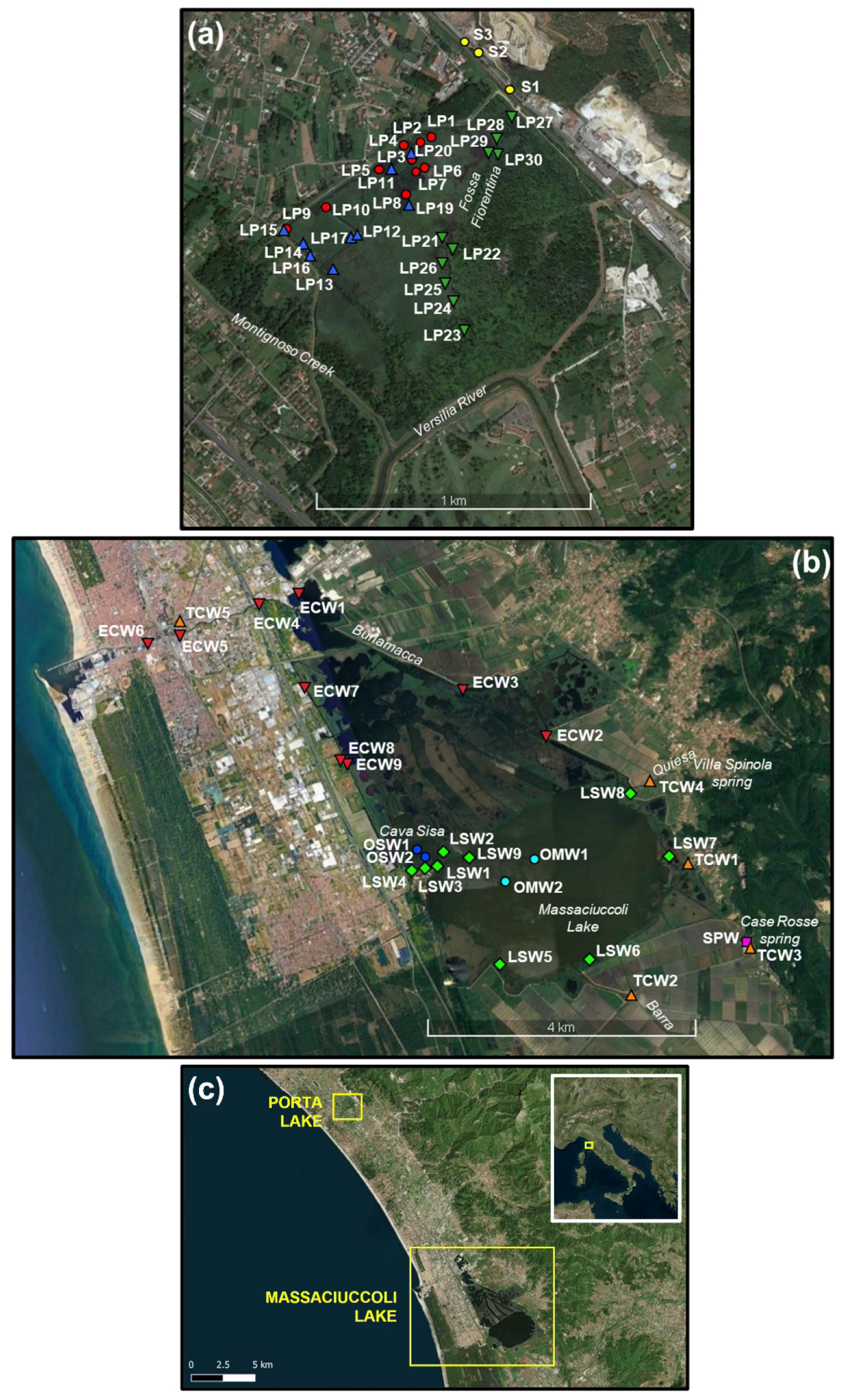
2.1. Study Areas
2.1.1. Porta Lake
2.1.2. Massaciuccoli Lake
2.2. Water and Dissolved Gas Sampling
2.3. Chemical and Isotopic Analysis of Water and Dissolved Gases
2.4. Diffusive Flux Calculation
3. Results
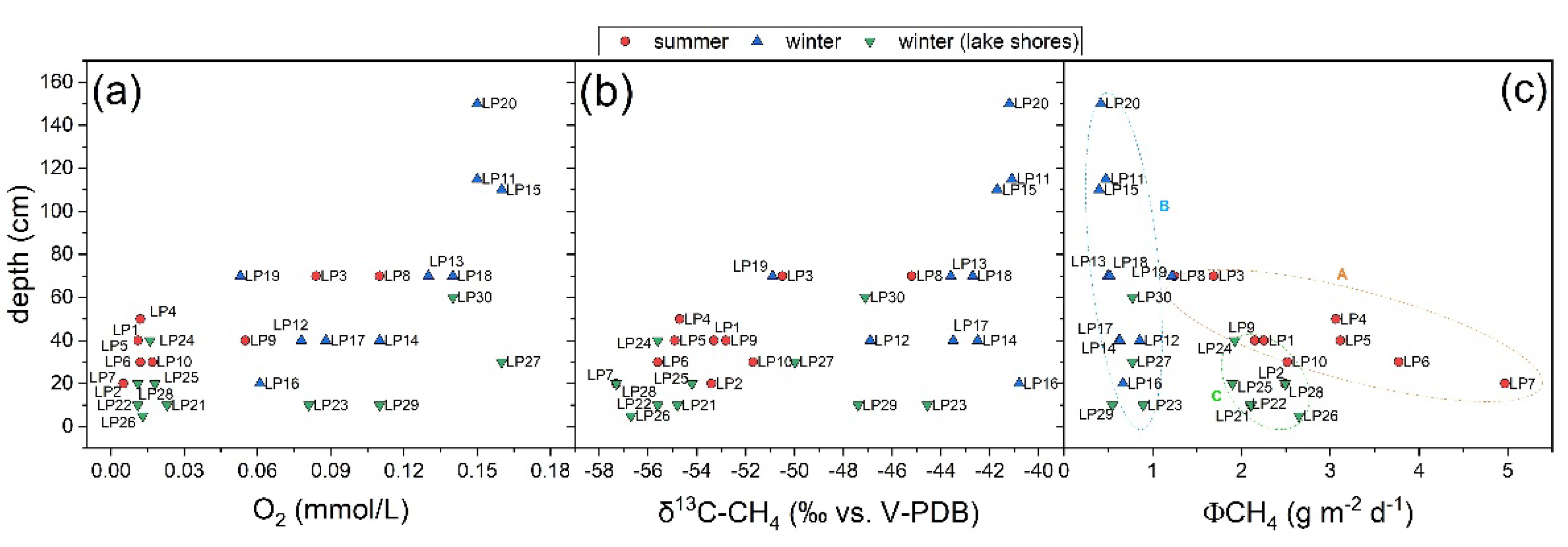
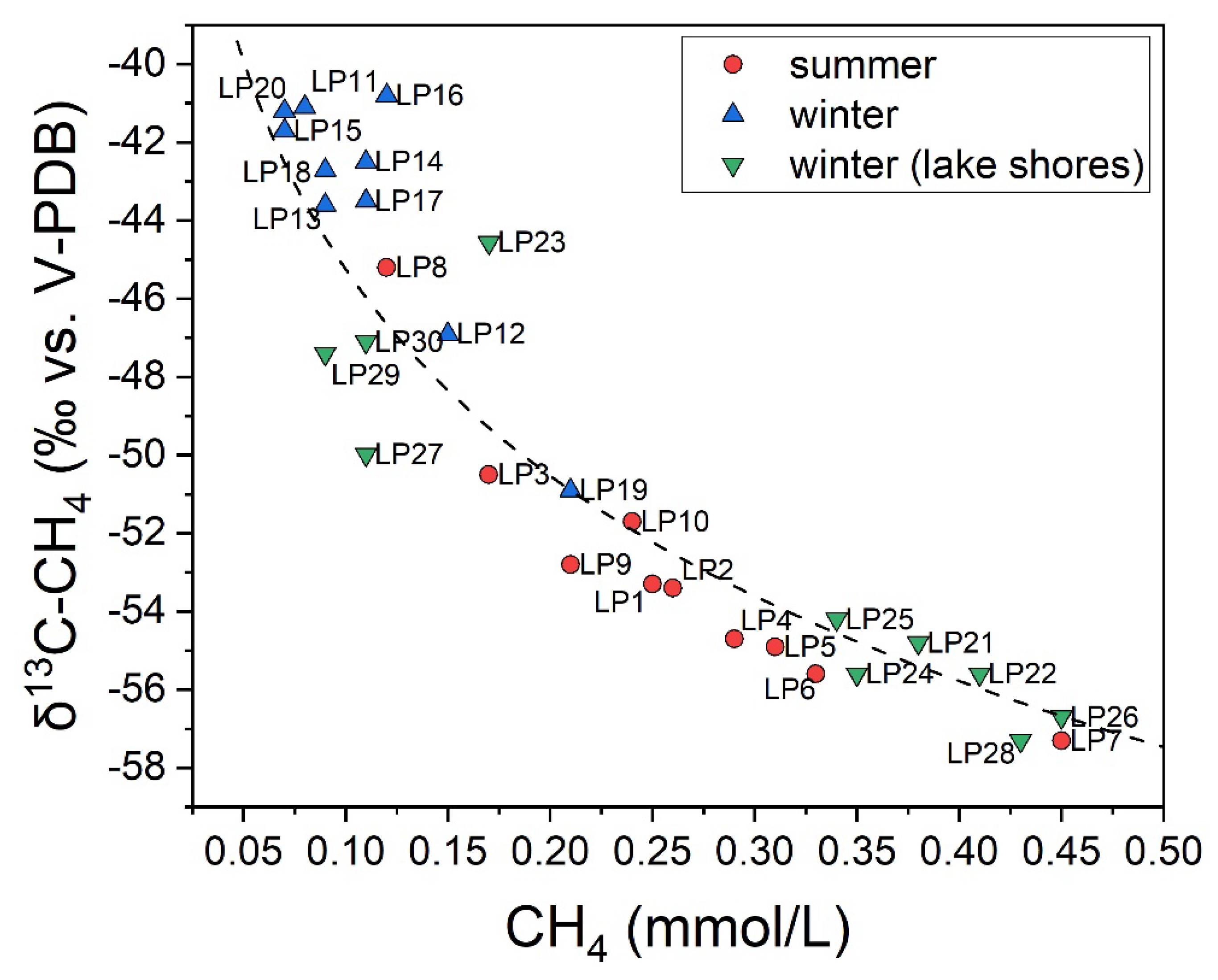
3.1. The Chemical Features of Porta Lake
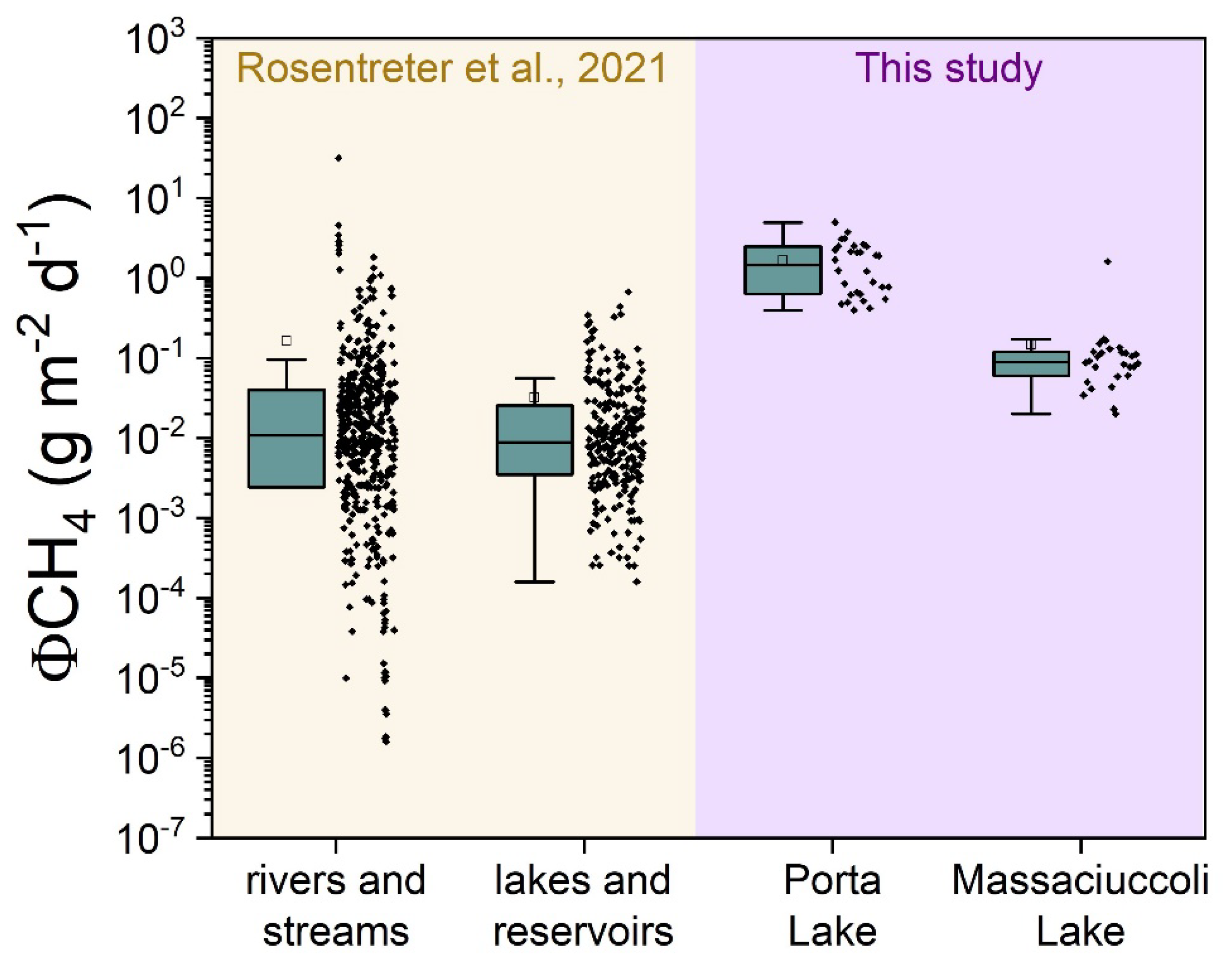
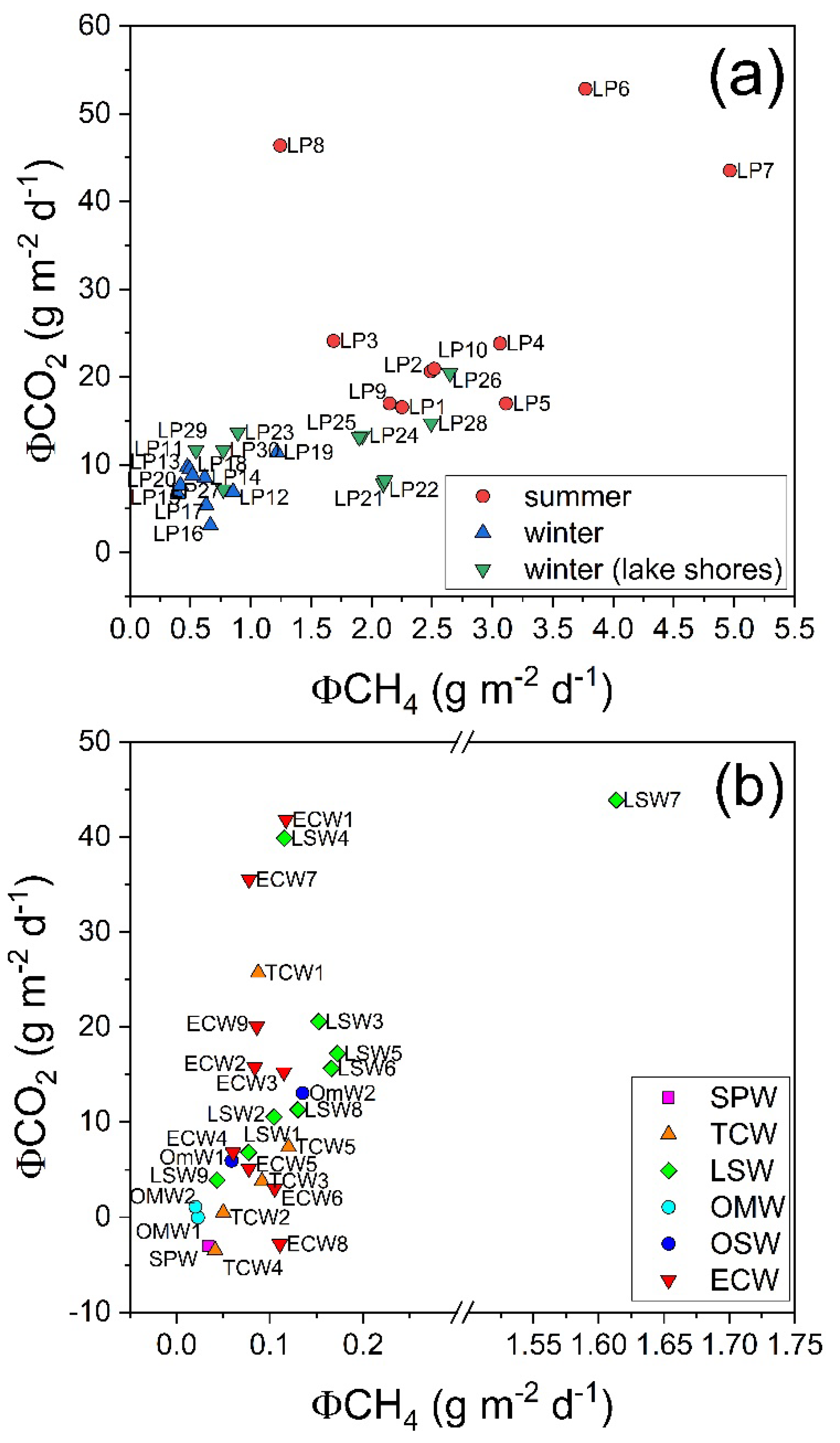
3.2. CH4 and CO2 Diffusive Fluxes at Porta Lake
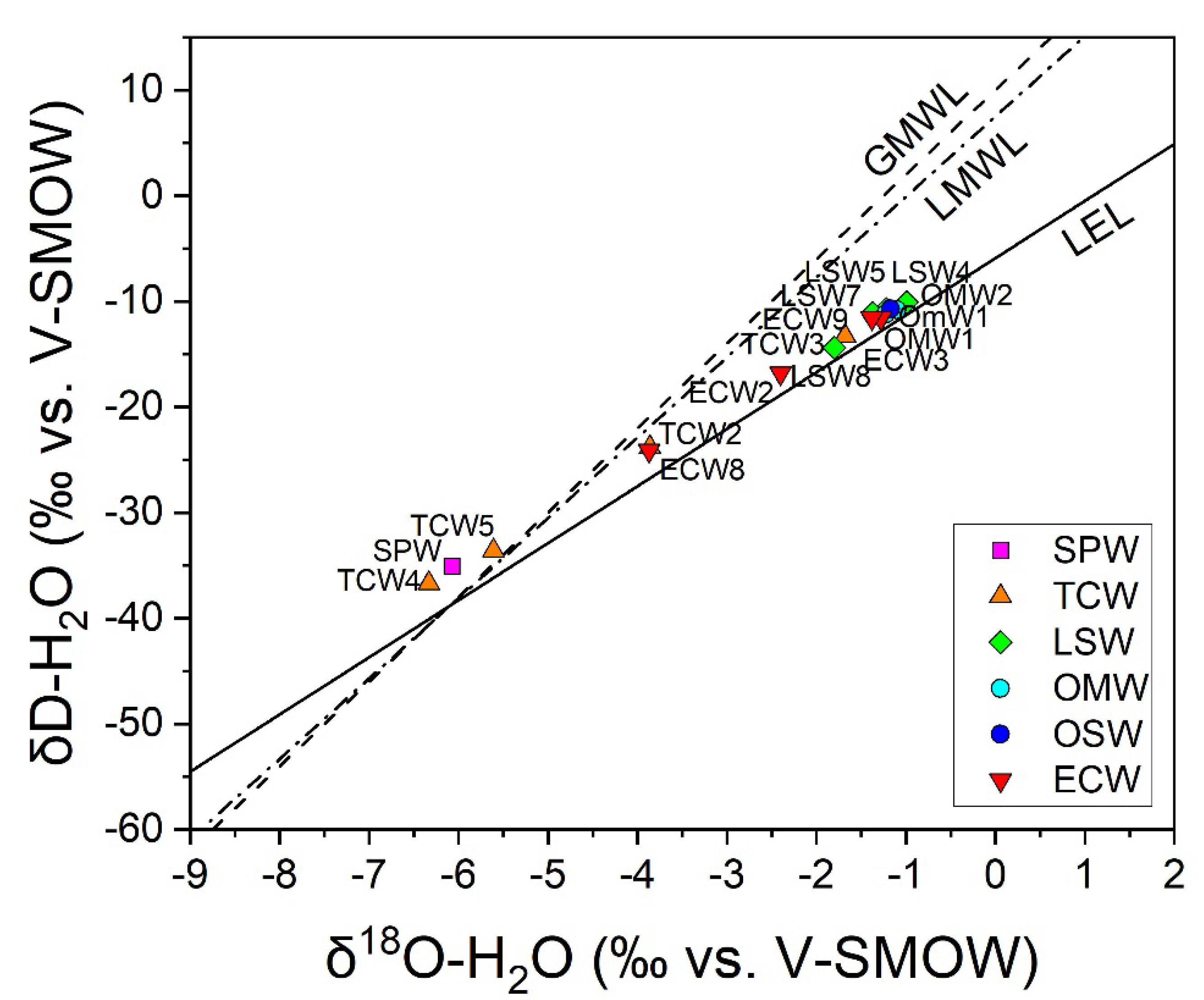
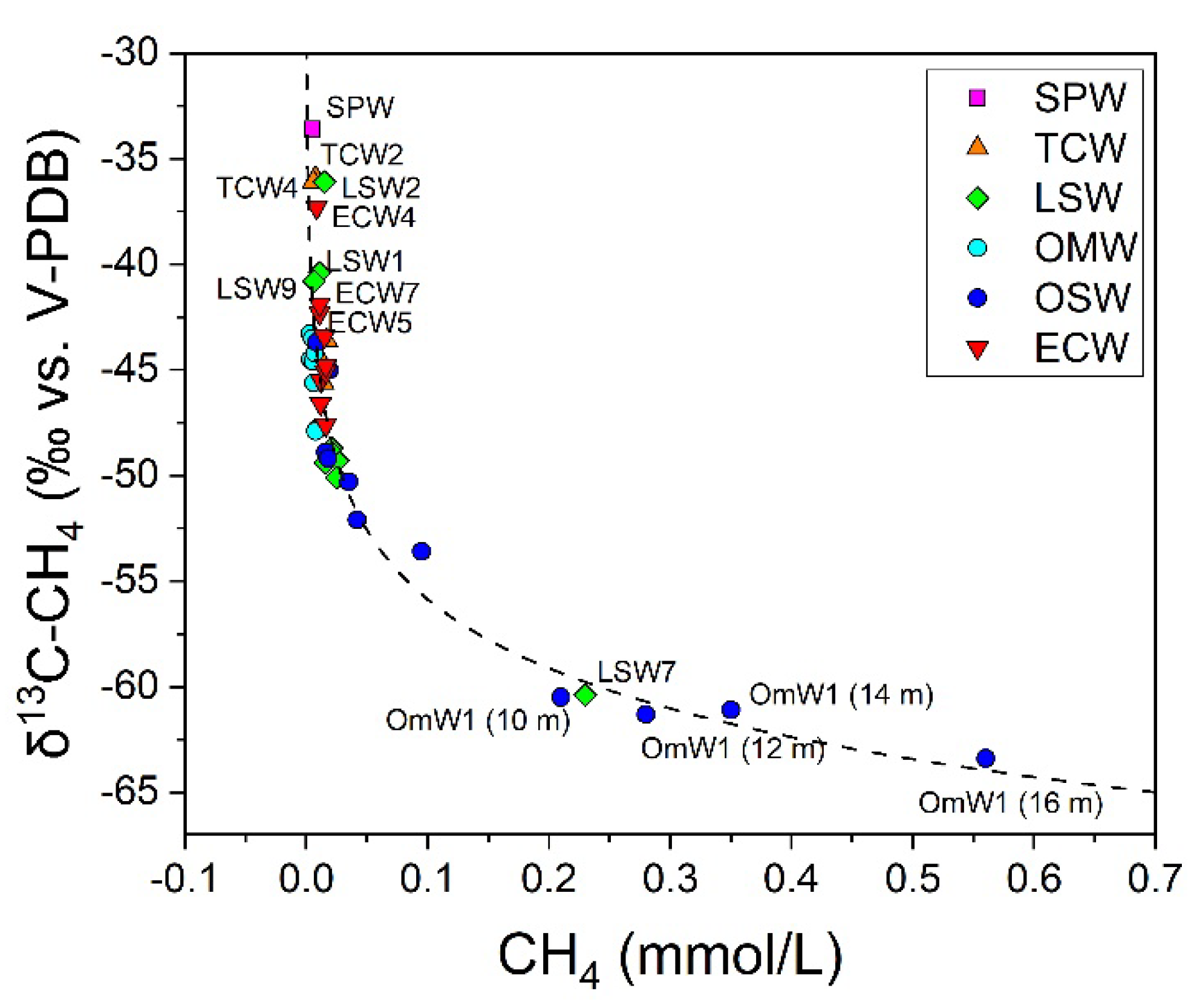
3.3. The Chemical Features of Massaciuccoli Lake System
3.4. CH4 and CO2 Diffusive Fluxes at Massaciuccoli Lake System
4. Discussion
4.1. CH4 Emission Drivers at Porta Lake
4.2. CH4 Emission Drivers at Massaciuccoli Lake System
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Downing, J.A.; Prairie, Y.T.; Cole, J.J.; Duarte, C.M.; Tranvik, L.J.; Striegl, R.G.; McDowell, W.H.; Kortelainen, P.; Caraco, N.F.; Melack, J.M.; et al. The gobal abundance and size distribution of lakes, ponds, and impoundments. Limnol. Oceanogr. 2006, 51, 2388–2397. [Google Scholar] [CrossRef] [Green Version]
- Ciężkowski, W.; Szporak-Wasilewska, S.; Kleniewska, M.; Jóźwiak, J.; Gnatowski, T.; Dąbrowski, P.; Góraj, M.; Szatyłowicz, J.; Ignar, S.; Chormański, J. Remotely sensed land surface temperature-based water stress index for wetland habitats. Remote Sens. 2020, 12, 631. [Google Scholar] [CrossRef] [Green Version]
- Koffi, E.N.; Bergamaschi, P.; Alkama, R.; Cescatti, A. An observation-constrained assessment of the climate sensitivity and future trajectories of wetland methane emissions. Sci. Adv. 2020, 6, eaay4444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salimi, S.; Almuktar, S.A.A.A.N.; Scholz, M. Impact of climate change on wetland ecosystems: A critical review of experimental wetlands. J. Environ. Manag. 2021, 286, 112160. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Carbon sequestration. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 815–830. [Google Scholar] [CrossRef]
- Nellemann, C.; Corcoran, E.; Duarte, C.M.; Valdrés, L.; De Young, C.; Fonseca, L.; Grimsditch, G.; Blue Carbon. A rapid response assessment.United Nations Environment Programme, GRID-Arendal. 2009. Available online: www.grida.no (accessed on 21 October 2021).
- Moomaw, W.R.; Chmura, G.L.; Davies, G.T.; Finlayson, C.M.; Middleton, B.A.; Natali, S.M.; Perry, J.E.; Roulet, N.; Sutton-Grier, A. Wetlands in a changing climate: Science, policy and management. Wetlands 2018, 38, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, J.J.; DelSontro, T.; Downing, J.A. Eutrophication will increase methane emissions from lakes and impoundments during the 21st century. Nat. Commun. 2019, 10, 1375. [Google Scholar] [CrossRef] [Green Version]
- Limpert, K.E.; Carnell, P.E.; Trevathan-Tackett, S.M.; Macreadie, P.I. Reducing emissions from degraded floodplain wetlands. Front. Environ. Sci. 2020, 8, 8. [Google Scholar] [CrossRef]
- Myhre, G.; Shindell, D.; Bréon, F.M.; Collins, W.; Fuglestvedt, J.; Huang, J.; Koch, D.; Lamarque, J.-F.; Lee, D.; Mendoza, B.; et al. Anthropogenic and Natural Radiative Forcing. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Rigby, M.; Prinn, R.G.; Fraser, P.J.; Simmonds, P.G.; Langenfelds, R.L.; Huang, J.; Cunnold, D.M.; Steele, L.P.; Krummel, P.B.; Weiss, R.F.; et al. Renewed growth of atmospheric methane. Geophys. Res. Lett. 2008, 35, L22805. [Google Scholar] [CrossRef] [Green Version]
- Dlugokencky, E.J.; Bruhwiler, L.; White, J.W.C.; Emmons, L.K.; Novelli, P.C.; Montzka, S.A.; Masarie, K.A.; Lang, P.M.; Crotwell, A.M.; Miller, J.B.; et al. Observational constraints on recent increases in the atmospheric CH4 burden. Geophys. Res. Lett. 2009, 36, L18803. [Google Scholar] [CrossRef] [Green Version]
- Dlugokencky, E. Trends in Atmospheric Methane. In NOAA/GML; 2021. Available online: https://gml.noaa.gov/ccgg/trends_ch4/ (accessed on 21 October 2021).
- Nisbet, E.G.; Dlugokencky, E.J.; Manning, M.R.; Lowry, D.; Fisher, R.E.; France, J.L.; Michel, S.E.; Miller, J.B.; White, J.W.C.; Vaughn, B.; et al. Rising atmospheric methane: 2007–2014 growth and isotopic shift. Glob. Biogeochem. Cycles 2016, 30, 1356–1370. [Google Scholar] [CrossRef] [Green Version]
- Nisbet, E.G.; Manning, M.R.; Dlugokencky, E.J.; Fisher, R.E.; Lowry, D.; Michel, S.E.; Lung Myhre, C.; Platt, S.M.; Allen, G.; Bousquet, P.; et al. Very strong atmospheric methane growth in the 4 years 2014–2014: Implications for the Paris Agreement. Glob. Biogeochem. Cycles 2019, 33, 318–342. [Google Scholar] [CrossRef]
- Lan, X.; Basu, S.; Schwietzke, S.; Bruhwiler, L.M.P.; Dlugokencky, E.J.; Michel, S.E.; Sherwood, O.A.; Tans, P.P.; Thoning, K.; Etiope, G.; et al. Improved constraints on global methane emissions and sinks using δ13C-CH4. Glob. Biogeochem. Cycles 2021, 35, e2021GB007000. [Google Scholar] [CrossRef]
- Rosentreter, J.A.; Borges, A.V.; Deemer, B.R.; Holgerson, M.A.; Liu, S.; Song, C.; Melack, J.; Raymond, P.A.; Duarte, C.M.; Allen, G.H.; et al. Half of global methane emissions come from highly variable aquatic ecosystem sources. Nat. Geosci. 2021, 14, 225–230. [Google Scholar] [CrossRef]
- Carmichael, M.J.; Bernhardt, E.S.; Bräuer, S.L.; Smith, W.K. The role of vegetation in methane flux to the atmosphere: Should vegetation be included as a distinct category in the global methane budget? Biogeochemistry 2014, 119, 1–24. [Google Scholar] [CrossRef]
- Waldo, N.B.; Hunt, B.K.; Fadely, E.C.; Moran, J.J.; Neumann, R.B. Plant root exudates increase methane emissions through direct and indirect pathways. Biogeochemistry 2019, 145, 213–234. [Google Scholar] [CrossRef]
- Tang, K.W.; McGinnis, D.F.; Ionescu, D.; Grossart, H.P. Methane production in oxic lake waters potentially increases aquatic methane flux to air. Environ. Sci. Technol. Lett. 2016, 3, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Donis, D.; Flury, S.; Stöckli, A.; Spangenberg, J.E.; Vachon, D.; McGinnis, D.F. Full-scale evaluation of methane production under oxic conditions in a mesotrophic lake. Nat. Commun. 2017, 8, 1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassi, F.; Cabassi, J.; Andrade, C.; Callieri, C.; Silva, C.; Viveiros, F.; Corno, G.; Vaselli, O.; Selmo, E.; Gallorini, A.; et al. Mechanisms regulating CO2 and CH4 dynamics in the Azorean volcanic lakes (São Miguel Island, Portugal). J. Limnol. 2018, 77, 483–504. [Google Scholar] [CrossRef] [Green Version]
- Günthel, M.; Donis, D.; Kirillin, G.; Ionescu, D.; Bizic, M.; McGinnis, D.F.; Grossart, H.P.; Tang, K.W. Contribution of oxic methane production to surface methane emission in lakes and its global importance. Nat. Commun. 2019, 10, 5497. [Google Scholar] [CrossRef] [Green Version]
- Fazi, S.; Amalfitano, S.; Venturi, S.; Pacini, N.; Vazquez, E.; Olaka, L.A.; Tassi, F.; Crognale, S.; Herzsprung, P.; Lechtenfeld, O.J.; et al. High concentrations of dissolved biogenic methane associated with cyanobacterial blooms in East African lake surface water. Commun. Biol. 2021, 4, 845. [Google Scholar] [CrossRef]
- Sanches, L.F.; Guenet, B.; Marinho, C.C.; Barros, N.; de Assis Esteves, F. Global regulation of methane emission from natural lakes. Sci. Rep. 2019, 9, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastviken, D.; Cole, J.; Pace, M.; Tranvik, L. Methane emissions from lakes: Dependence of, GB lake characteristics, two regional assessments, and a global estimate. Glob. Biogeochem. Cycles 2004, 18, GB4009. [Google Scholar] [CrossRef]
- Hofmann, H. Spatiotemporal distribution patterns of dissolved methane in lakes: How accurate are the current estimations of the diffusive flux path? Geophys. Res. Lett. 2013, 40, 2779–2784. [Google Scholar] [CrossRef]
- Attermeyer, K.; Flury, S.; Jayakumar, R.; Fiener, P.; Steger, K.; Arya, V.; Wilken, F.; van Geldern, R.; Premke, K. Invasive floating macrophytes reduce greenhouse gas emissions from a small tropical lake. Sci. Rep. 2016, 6, 20424. [Google Scholar] [CrossRef]
- dos Santos Fonseca, A.L.; Cardoso Marinho, C.; de Assis Esteves, F. Floating aquatic macrophytes decrease the methane concentration in the water column of a tropical coastal lagoon: Implications for methane oxidation and emission. Braz. Arch. Biol. Technol. 2017, 60, e17160381. [Google Scholar]
- Milberg, P.; Törnqvist, L.; Westerberg, L.M.; Bastviken, D. Temporal variations in methane emissions from emergent aquatic macrophytes in two boreonemoral lakes. AoB Plants 2017, 9, plx029. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, L.C.; Maher, D.T.; Johnston, S.G.; Kelaher, B.P.; Steven, A.; Tait, D.R. Wetland methane emissions dominated by plant-mediated fluxes: Contrasting emissions pathways and seasons within a shallow freshwater subtropical wetland. Limnol. Oceanogr. 2019, 64, 1895–1912. [Google Scholar] [CrossRef]
- Bansal, S.; Johnson, O.F.; Meier, J.; Zhu, X. Vegetation affects timing and location of wetland methane emissions. J. Geophys. Res. Biogeosci. 2020, 125, e2020JG005777. [Google Scholar] [CrossRef]
- van den Berg, M.; Ingwersen, J.; Lamers, M.; Streck, T. The role of Phragmites in the CH4 and CO2 fluxes in a minerotrophic peatland in southwest Germany. Biogeosciences 2016, 13, 6107–6119. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, M.; van den Elzen, E.; Ingwersen, J.; Kosten, S.; Lamers, L.P.M.; Streck, T. Contribution of plant-induced pressurized flow to CH4 emission from a Phragmites fen. Sci. Rep. 2020, 10, 12304. [Google Scholar] [CrossRef]
- Kim, J.; Chaudhary, D.R.; Lee, J.; Byun, C.; Byun, C.; Ding, W.; Kwon, B.O.; Khim, J.S.; Kang, H. Microbial mechanism for enhanced methane emission in deep soil layer of Phragmites-introduced tidal marsh. Environ. Int. 2020, 134, 105251. [Google Scholar] [CrossRef] [PubMed]
- Deemer, B.R.; Holgerson, M.A. Drivers of methane flux differ between lakes and reservoirs, complicating global upscaling efforts. J. Geophys. Res. Biogeosci. 2021, 126, e2019JG005600. [Google Scholar] [CrossRef]
- Vaselli, O.; Venturi, S.; Cabassi, J.; Lazzaroni, M.; Randazzo, A.; Tassi, F.; Capecchiacci, F.; Giannini, L.; Vietina, B. Monitoraggio di parametri fisico-chimici di acque e sedimenti e modello idrogeochimico del Lago di Porta (Massa-Carrara, Toscana); Technical Report of the Department of Earth Sciences, University of Florence, for the Municipality of Montignoso (Massa-Carrara, Tuscany); University of Florence: Florence, Italy, 2021. [Google Scholar]
- Ganesan, A.L.; Stell, A.C.; Gedney, N.; Comyn-Platt, E.; Hayman, G.; Rigby, M.; Poulter, B.; Hornibrook, E.R.C. Spatially resolved isotopic source signatures of wetland methane emissions. Geophys. Res. Lett. 2018, 45, 3737–3745. [Google Scholar] [CrossRef]
- Da Prato, S.; Baneschi, I. The coastal wetland systems of northern Tuscany: Massaciuccoli Lake and ex Porta Lake. State of knowledge and new opportunities for multidisciplinary approach. Ital. J. Groundw. 2020, 9, 51–54. [Google Scholar] [CrossRef]
- Costantini, E.A.C.; Barbetti, R.; Fantappiè, M.; L’Abate, G.; Lorenzetti, R.; Magini, S. Pedodiversity. In The Soils of Italy; Costantini, E.A.C., Dazzi, C., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 105–178. [Google Scholar]
- Turba, C.A.; Lunardini, F. Studio geologico, geomorfologico ed idrogeologico. A.I.A. Progetto di completamento della discarica per rifiuti speciali non pericolosi sita in Loc. Porta; Ambiente Apuane S.p.A.: Massa Carrara, Italy, 2011; p. 238. [Google Scholar]
- van der Putten, W.H. Assessing ecological change in European wetlands: How to know what parameters should be monitored to evaluate the die-back of common reed (Phragmites australis)? Stapfia 1994, 31, 61–68. [Google Scholar]
- van der Putten, W.H. Die-back of Phragmites australis in European wetlands: An overview of the European Research Programme on reed die-back and progression (1993–1994). Aquat. Biol. 1997, 59, 267–275. [Google Scholar] [CrossRef]
- Gigante, D.; Venanzoni, R.; Zuccarello, V. Reed die-back in southern Europe? A case study from Central Italy. C. R. Biol. 2011, 334, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Lastrucci, L.; Gigante, D.; Vaselli, O.; Nisi, B.; Viciani, D.; Reale, L.; Coppi, A.; Fazzi, V.; Bonari, G.; Angiolini, C. Sediment chemistry and flooding exposure: A fatal cocktail for Phragmites australis in the Mediterranean basin? Ann. Limnol. – Int. J. Limnol. 2016, 52, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Baldaccini, G.N. Zone umide: Dal degrado al recupero ecologico. Il caso del lago di Massaciuccoli (Toscana nord-occidentale). Biol. Ambient. 2018, 32, 85–98. [Google Scholar]
- Baneschi, I. Geochemical and Environmental Study of a Coastal Ecosystem: Massaciuccoli Lake (Northern Tuscany, Italy). Ph.D. Thesis, Department of Environmental Sciences, University Cà Foscari of Venice, Venice, Italy, 2007. [Google Scholar]
- Ciurli, A.; Zuccari, P.; Alpi, A. Growth and nutrient absorption of two submerged aquatic macrophytes in mesocosms, for reinsertion in a eutrophicated shallow lake. Wetl. Ecol. Manag. 2009, 17, 107–115. [Google Scholar] [CrossRef]
- Zuccarini, P.; Ciurli, A.; Alpi, A. Implications for shallow lake manipulation: Results of aquaria and enclosure experiments manipulating macrophytes, zooplankton and fish. Appl. Ecol. Environ. Res. 2011, 9, 123–140. [Google Scholar] [CrossRef]
- Viciani, D.; Dell’Olmo, L.; Vicenti, C.; Lastrucci, L. Natura 2000 protected habitats, Massaciuccoli Lake (northern Tuscany, Italy). J. Maps 2017, 13, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Caliro, S.; Panichi, C.; Stanzione, D. Variation in the total dissolved carbon isotope composition of thermal waters of the Island of Ischia (Italy) and its implications for volcanic surveillance. J. Volcanol. Geotherm. Res. 1999, 90, 219–240. [Google Scholar] [CrossRef]
- Tassi, F.; Vaselli, O.; Luchetti, G.; Montegrossi, G.; Minissale, A. Metodo per la determinazione dei gas disciolti in acque naturali; Int. Rep.; CNR-IGG: Florence, Italy, 2008; p. 11. [Google Scholar]
- Tassi, F.; Vaselli, O.; Tedesco, D.; Montegrossi, G.; Darrah, T.; Cuoco, E.; Mapendano, M.Y.; Poreda, R.; Delgado Huertas, A. Water and gas chemistry at Lake Kivu (DRC): Geochemical evidence of vertical and horizontal heterogeneities in a multi-basin structure. Geochem. Geophys. Geosyst. 2009, 10, Q02005. [Google Scholar] [CrossRef]
- Tassi, F.; Fazi, S.; Rossetti, S.; Pratesi, P.; Ceccotti, M.; Cabassi, J.; Capecchiacci, F.; Venturi, S.; Vaselli, O. The biogeochemical vertical structure renders a meromictic volcanic lake a trap for geogenic CO2 (Lake Averno, Italy). PLoS ONE 2018, 13, e0193914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salata, G.G.; Roelke, L.A.; Cifuentes, L.A. A rapid and precise method for measuring stable carbon isotope ratios of dissolved inorganic carbon. Mar. Chem. 2000, 69, 153–161. [Google Scholar] [CrossRef]
- Venturi, S.; Tassi, F.; Bicocchi, G.; Cabassi, J.; Capecchiacci, F.; Capasso, G.; Vaselli, O.; Ricci, A.; Grassa, F. Fractionation processes affecting the stable carbon isotope signature of thermal waters from hydrothermal/volcanic systems: The examples of Campi Flegrei and Volcano Island (southern Italy). J. Volcanol. Geotherm. Res. 2017, 345, 46–57. [Google Scholar] [CrossRef]
- Chiodini, G. Gases dissolved in groundwaters: Analytical methods and examples of applications in central Italy. In Proceedings of the Rome Seminar on Environmental Geochemistry, Castelnuovo Di Porto, Rome, Italy, 22–26 May 1996; pp. 135–148. [Google Scholar]
- Caliro, S.; Chiodini, G.; Avino, R.; Cardellini, C.; Frondini, F. Volcanic degassing at Somma-Vesuvio (Italy) inferred by chemical and isotopic signatures of groundwater. Appl. Geochem. 2005, 20, 1060–1076. [Google Scholar] [CrossRef]
- Frondini, F.; Cardellini, C.; Caliro, S.; Chiodini, G.; Morgantini, N. Regional groundwater flow and interactions with deep fluids in western Apennine: The case of Narni-Amelia chain (Central Italy). Geofluids 2012, 12, 182–196. [Google Scholar] [CrossRef]
- Zhang, J.; Quay, P.D.; Wilbur, D.O. Carbon isotope fractionation during gas-water exchange and dissolution of CO2. Geochim. Cosmochim. Acta 1995, 59, 107–114. [Google Scholar] [CrossRef]
- Liss, P.S.; Slater, P.G. Flux of Gases across the Air-Sea Interface. Nature 1974, 247, 181–184. [Google Scholar] [CrossRef]
- Wanninkhof, R. Relationship between wind speed and gas exchange over the ocean revisited. Limnol. Oceanogr. Methods 2014, 12, 351–362. [Google Scholar] [CrossRef]
- Crusius, J.; Wanninkhof, R. Gas transfer velocities measured at low wind speed over a lake. Limnol. Oceanogr. 2003, 48, 1010–1017. [Google Scholar] [CrossRef]
- Nightingale, P.D.; Malin, G.; Law, C.S.; Watson, A.J.; Liss, P.S.; Liddicoat, M.I.; Boutin, J.; Upstill-Goddard, R.C. In situ evaluation of air-sea gas exchange parametrizations using novel conservative and volatile tracers. Global Biogeochem. Cycles 2000, 14, 373–387. [Google Scholar] [CrossRef]
- Hoover, T.E.; Berkshire, D.C. Effects of hydration on carbon dioxide exchange across an air-water interface. J. Geophys. Res. 1969, 74, 456–464. [Google Scholar] [CrossRef]
- Wanninkhof, R.; Knox, M. Chemical enhancement of CO2 exchange in natural waters. Limnol. Oceanogr. 1996, 41, 689–697. [Google Scholar] [CrossRef]
- Zeebe, R.E. On the molecular diffusion coefficients of dissolved CO, HCO3-, and CO32- and their dependence on isotopic mass. Geochim. Cosmochim. Acta 2011, 75, 2483–2498. [Google Scholar] [CrossRef]
- Johnson, K.S. Carbon dioxide hydration and dehydration kinetics in seawater. Limnol. Oceanogr. 1982, 27, 849–855. [Google Scholar] [CrossRef]
- Clark, I. Groundwater Geochemistry and Isotopes; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Deppenmeier, U.; Müller, V.; Gottschalk, G. Pathways of energy conservation in methanogenic archaea. Arch. Microbiol. 1996, 165, 149–163. [Google Scholar] [CrossRef]
- Conrad, R. Importance of hydrogenotrophic, aceticlastic and methylotrophic methanogenesis for methane production in terrestrial, aquatic and other anoxic environments: A mini review. Pedosphere 2020, 30, 25–39. [Google Scholar] [CrossRef]
- Gruca-Rokosz, R.; Szal, D.; Bartoszek, L.; Pękala, A. Isotopic evidence for vertical diversification of methane production pathways in freshwater sediments of Nielisz reservoir (Poland). CATENA 2020, 195, 104803. [Google Scholar] [CrossRef]
- Praetzel, L.S.E.; Plenter, N.; Schilling, S.; Schmiedeskamp, M.; Broll, G.; Knorr, K.H. Organic matter and sediment properties determine in-lake variability of sediment CO2 and CH4 production and emissions of a small and shallow lake. Biogeosciences 2020, 17, 5057–5078. [Google Scholar] [CrossRef]
- Whiticar, M.J. Carbon and hydrogen isotope systematics of bacterial formation and oxidation of methane. Chem. Geol. 1999, 161, 291–314. [Google Scholar] [CrossRef]
- Conrad, R. Quantification of methanogenic pathways using stable carbon isotopic signatures: A review and a proposal. Org. Geochem. 2005, 36, 739–752. [Google Scholar] [CrossRef]
- Sorrell, B.K.; Brix, H.; Schierup, H.H.; Lorenzen, B. Die-back of Phragmites australis: Influence on the distribution and rate of sediment methanogenesis. Biogeochemistry 1997, 36, 173–188. [Google Scholar] [CrossRef]
- Reitsema, R.E.; Meire, P.; Schoelynck, J. The future of freshwater macrophytes in a changing world: Dissolved organic carbon quantity and quality and its interactions with macrophytes. Front. Plant Sci. 2018, 9, 629. [Google Scholar] [CrossRef]
- Grasset, C.; Abril, G.; Mendonça, R.; Roland, F.; Sobek, S. The transformation of macrophyte-derived organic matter to methane relates to plant water and nutrient contents. Limnol. Oceanogr. 2019, 64, 1737–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thottathil, S.D.; Prairie, Y.T. Coupling of stable carbon isotopic signature of methane and ebullitive fluxes in northern temperate lakes. Sci. Total Environ. 2021, 777, 146117. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Lenstra, W.; Jong, D.; Meysman, F.J.R.; Sapart, C.J.; van der Veen, C.; Röckmann, T.; Gonzalez, S.; Slomp, C.P. Rapid sediment accumulation results in high methane effluxes from coastal sediments. PLoS ONE 2016, 11, e0161609. [Google Scholar] [CrossRef] [Green Version]
- Martins, P.D.; Hoyt, D.W.; Bansal, S.; Mills, C.T.; Tfaily, M.; Tangen, B.A.; Finocchiaro, R.G.; Johnston, M.D.; McAdams, B.C.; Solensky, M.J.; et al. Abundant carbon substrates drive extremely high sulfate reduction rates and methane fluxes in Prairie Pothole Wetlands. Glob. Chang. Biol. 2017, 23, 3107–3120. [Google Scholar] [CrossRef] [PubMed]
- Sela-Adler, M.; Ronen, Z.; Herut, B.; Antler, G.; Vigderovich, H.; Eckert, W.; Sivan, O. Co-existence of methanogenesis and sulfate reduction with common substrates in sulfate-rich estuarine sediments. Front. Microbiol. 2017, 8, 766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, A.; Lyautey, E.; Montuelle, B.; Casper, P. Effects of increasing temperatures on methane concentrations and methanogenesis during experimental incubation of sediments from oligotrophic and mesotrophic lakes. J. Geophys. Res. Biogeosci. 2016, 121, 1394–1406. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Xie, P.; Yang, H.; Wu, A.; Ni, L. The change of gaseous carbon fluxes following the switch of dominant producers from macrophytes to algae in a shallow subtropical lake of China. Atmos. Environ. 2006, 40, 8034–8043. [Google Scholar] [CrossRef]
- Zhang, M.; Xiao, Q.; Zhang, Z.; Gao, Y.; Zhao, J.; Pu, Y.; Wang, W.; Xiao, W.; Liu, S.; Lee, X. Methane flux dynamics in a submerged aquatic vegetation zone in a subtropical lake. Sci. Total Environ. 2019, 672, 400–409. [Google Scholar] [CrossRef]
- Emilson, E.J.S.; Carson, M.A.; Yakimovich, K.M.; Osterholz, H.; Dittmar, T.; Gunn, J.M.; Mykytczuk, N.C.S.; Basiliko, N.; Tanentzap, A.J. Climate-driven shifts in sediment chemistry enhance methane production in northern lakes. Nat. Commun. 2018, 9, 1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiellini, C.; Guglielminetti, L.; Sarrocco, S.; Ciurli, A. Isolation of four microalgal strains from the Lake Massaciuccoli: Screening of common pollutants tolerance pattern and perspectives for their use in biotechnological applications. Front. Plant Sci. 2020, 11, 607651. [Google Scholar] [CrossRef]
- Craig, H. Isotopic variations in meteoric waters. Science 1961, 133, 1702–1703. [Google Scholar] [CrossRef]
- Bastviken, D.; Nygren, J.; Schenk, J.; Parellada Massana, R.; Duc, N.T. Technical note: Facilitating the use of low-cost methane (CH4) sensors in flux chambers—Calibration, data processing, and an open-source make-it-yourself logger. Biogeosciences 2020, 17, 3659–3667. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | E | N | w.c.d. | T | pH | Eh | HCO3 | Cl | SO4 | Na | K | Ca | Mg | NH4 | NO3 | F | Br | TDS | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| summertime | LP1 | 593756 | 4872054 | 40 | 23.7 | 7.41 | −153 | 215 | 27 | 328 | 16 | 1.7 | 163 | 32 | 0.33 | 2.1 | 0.76 | 0.09 | 787 |
| LP2 | 593716 | 4872032 | 20 | 25.6 | 7.18 | −295 | 204 | 24 | 301 | 16 | 2.0 | 153 | 32 | 0.23 | 0.11 | 0.33 | 0.08 | 733 | |
| LP3 | 593687 | 4871974 | 70 | 26.7 | 7.4 | 89 | 201 | 26 | 328 | 16 | 1.7 | 161 | 32 | 0.08 | 0.61 | 0.44 | 0.09 | 768 | |
| LP4 | 593661 | 4872021 | 50 | 28.7 | 7.47 | −32 | 192 | 26 | 325 | 17 | 2.5 | 160 | 33 | 0.11 | 0.06 | 0.49 | 0.08 | 755 | |
| LP5 | 593573 | 4871929 | 40 | 27.1 | 7.4 | −66 | 205 | 26 | 323 | 17 | 2.2 | 162 | 32 | 0.16 | 0.40 | 0.33 | 0.09 | 768 | |
| LP6 | 593732 | 4871939 | 30 | 31.2 | 8.11 | −129 | 192 | 27 | 327 | 17 | 1.9 | 160 | 32 | 0.27 | 0.25 | 0.47 | 0.09 | 757 | |
| LP7 | 593704 | 4871926 | 20 | 30.1 | 7.54 | −192 | 205 | 26 | 320 | 17 | 1.4 | 161 | 32 | 0.38 | 0.04 | 0.32 | 0.08 | 763 | |
| LP8 | 593666 | 4871844 | 70 | 28.1 | 7.64 | 20 | 199 | 27 | 328 | 17 | 2.2 | 163 | 33 | 0.09 | 0.45 | 0.36 | 0.09 | 769 | |
| LP9 | 593226 | 4871708 | 40 | 27.7 | 7.3 | −51 | 201 | 27 | 307 | 17 | 1.6 | 160 | 32 | 0.20 | 0.14 | 0.61 | 0.09 | 746 | |
| LP10 | 593377 | 4871788 | 30 | 28.5 | 7.4 | −120 | 201 | 27 | 334 | 16 | 2.1 | 160 | 32 | 0.21 | 0.13 | 0.38 | 0.09 | 773 | |
| wintertime | LP11 | 593613 | 4871927 | 115 | 11.3 | 7.4 | 96 | 217 | 20 | 281 | 19 | 1.3 | 150 | 30 | 0.09 | 1.97 | 0.24 | 0.06 | 722 |
| LP12 | 593493 | 4871693 | 40 | 10.3 | 7.5 | 35 | 216 | 20 | 272 | 19 | 2.3 | 147 | 30 | 0.08 | 1.46 | 0.23 | 0.15 | 708 | |
| LP13 | 593402 | 4871555 | 70 | 9.4 | 7.4 | 87 | 204 | 18 | 273 | 18 | 2.1 | 130 | 27 | 0.08 | 0.37 | 0.18 | 0.09 | 672 | |
| LP14 | 593298 | 4871649 | 40 | 10.0 | 7.55 | 24 | 220 | 19 | 245 | 17 | 2.4 | 138 | 26 | 0.11 | 1.47 | 0.22 | 0.08 | 670 | |
| LP15 | 593219 | 4871708 | 110 | 10.2 | 7.48 | 95 | 221 | 22 | 285 | 20 | 3.6 | 151 | 30 | 0.11 | 1.86 | 0.36 | 0.09 | 735 | |
| LP16 | 593325 | 4871615 | 20 | 9.6 | 7.49 | 12 | 219 | 19 | 215 | 18 | 2.7 | 118 | 24 | 0.37 | 0.82 | 0.80 | 0.09 | 618 | |
| LP17 | 593473 | 4871679 | 40 | 10.5 | 7.45 | 19 | 220 | 19 | 258 | 19 | 1.2 | 137 | 30 | 0.28 | 1.42 | 0.24 | 0.06 | 686 | |
| LP18 | 593376 | 4871786 | 70 | 10.6 | 7.54 | 52 | 218 | 19 | 274 | 21 | 4.3 | 143 | 31 | 0.13 | 1.60 | 0.21 | 0.08 | 713 | |
| LP19 | 593680 | 4871797 | 70 | 10.8 | 7.53 | 4 | 215 | 21 | 261 | 20 | 2.6 | 136 | 30 | 0.53 | 1.93 | 0.20 | 0.08 | 688 | |
| LP20 | 593684 | 4871988 | 150 | 11.6 | 7.5 | 81 | 215 | 19 | 257 | 19 | 1.6 | 135 | 30 | 0.19 | 2.68 | 0.28 | 0.08 | 680 | |
| LP21 | 593805 | 4871686 | 10 | 9.5 | 7.2 | −77 | 331 | 46 | 129 | 25 | 33 | 105 | 33 | 0.51 | 1.02 | 0.13 | 0.17 | 703 | |
| LP22 | 593837 | 4871648 | 10 | 7.7 | 7.25 | −80 | 386 | 36 | 39 | 22 | 7.9 | 95 | 31 | 0.79 | 0.48 | 0.18 | 0.12 | 618 | |
| LP23 | 593885 | 4871350 | 10 | 8.2 | 7.52 | 12 | 398 | 70 | 30 | 44 | 7.1 | 95 | 34 | 0.27 | 0.04 | 0.08 | 0.22 | 678 | |
| LP24 | 593841 | 4871464 | 40 | 9.4 | 7.5 | −54 | 432 | 33 | 61 | 32 | 5.7 | 116 | 34 | 0.73 | 0.02 | 0.17 | 0.14 | 715 | |
| LP25 | 593814 | 4871521 | 20 | 9.8 | 7.6 | −31 | 322 | 26 | 145 | 25 | 4.4 | 122 | 31 | 0.54 | 0.02 | 0.19 | 0.12 | 676 | |
| LP26 | 593800 | 4871601 | 5 | 11.2 | 7.6 | −98 | 272 | 23 | 188 | 20 | 3.8 | 128 | 30 | 0.61 | 3.26 | 0.26 | 0.11 | 669 | |
| LP27 | 594047 | 4872130 | 30 | 16.1 | 7.17 | 85 | 212 | 24 | 363 | 18 | 2.7 | 177 | 34 | 0.05 | 7.68 | 0.43 | 0.09 | 839 | |
| LP28 | 593996 | 4872049 | 20 | 10.8 | 7.71 | −140 | 262 | 25 | 217 | 22 | 4.9 | 124 | 29 | 0.43 | 0.70 | 0.45 | 0.04 | 686 | |
| LP29 | 593972 | 4872003 | 10 | 12.0 | 7.78 | 27 | 221 | 19 | 323 | 21 | 3.0 | 157 | 34 | 0.04 | 5.08 | 0.21 | 0.08 | 784 | |
| LP30 | 593997 | 4871997 | 60 | 16.1 | 7.69 | 53 | 217 | 26 | 337 | 24 | 3.0 | 169 | 33 | 0.04 | 7.20 | 0.32 | 0.05 | 816 | |
| spring waters | S1 | 594043 | 4872228 | 17.0 | 7.8 | 155 | 212 | 32 | 286 | 23 | 2.3 | 156 | 29 | 0.02 | 7.2 | 0.45 | 0.09 | 748 | |
| S2 | 593923 | 4872362 | 17.5 | 7.92 | 141 | 195 | 20 | 375 | 12 | 2.2 | 171 | 33 | 0.02 | 7.0 | 0.39 | 0.05 | 814 | ||
| S3 | 593876 | 4872404 | 18.0 | 7.76 | 160 | 195 | 19 | 356 | 12 | 2.5 | 171 | 33 | 0.02 | 6.7 | 0.42 | 0.04 | 795 |
| ID | CO2 | N2 | Ar | CH4 | O2 | δ13C-CO2 | δ13C-CH4 | ΦCH4 | ΦCO2 | ΦCO2eq |
|---|---|---|---|---|---|---|---|---|---|---|
| LP1 | 0.09 | 0.53 | 0.013 | 0.25 | 0.011 | −21.5 | −53.3 | 2.25 | 16.6 | 79.6 |
| LP2 | 0.14 | 0.51 | 0.013 | 0.26 | 0.005 | −20.9 | −53.4 | 2.49 | 20.6 | 90.4 |
| LP3 | 0.11 | 0.52 | 0.013 | 0.17 | 0.084 | −18.7 | −50.5 | 1.69 | 24.1 | 71.3 |
| LP4 | 0.09 | 0.53 | 0.014 | 0.29 | 0.012 | −20.8 | −54.7 | 3.06 | 23.8 | 110 |
| LP5 | 0.08 | 0.49 | 0.012 | 0.31 | 0.011 | −21.3 | −54.9 | 3.11 | 17.0 | 104 |
| LP6 | 0.07 | 0.51 | 0.013 | 0.33 | 0.012 | −21.8 | −55.6 | 3.77 | 52.8 | 158 |
| LP7 | 0.13 | 0.49 | 0.013 | 0.45 | 0.005 | −22.1 | −57.3 | 4.97 | 43.5 | 183 |
| LP8 | 0.13 | 0.52 | 0.013 | 0.12 | 0.11 | −17.6 | −45.2 | 1.24 | 46.4 | 81.2 |
| LP9 | 0.09 | 0.55 | 0.014 | 0.21 | 0.055 | −21.3 | −52.8 | 2.15 | 17.0 | 77.2 |
| LP10 | 0.09 | 0.52 | 0.013 | 0.24 | 0.017 | −21.4 | −51.7 | 2.52 | 20.9 | 91.5 |
| LP11 | 0.11 | 0.68 | 0.017 | 0.08 | 0.15 | −18.4 | −41.1 | 0.472 | 9.73 | 23.0 |
| LP12 | 0.08 | 0.71 | 0.018 | 0.15 | 0.078 | −18.5 | −46.9 | 0.853 | 6.94 | 30.8 |
| LP13 | 0.12 | 0.69 | 0.018 | 0.09 | 0.13 | −18.7 | −43.6 | 0.495 | 9.50 | 23.3 |
| LP14 | 0.09 | 0.67 | 0.017 | 0.11 | 0.11 | −18.7 | −42.5 | 0.619 | 8.57 | 25.9 |
| LP15 | 0.08 | 0.69 | 0.017 | 0.07 | 0.16 | −18.2 | −41.7 | 0.397 | 6.68 | 17.8 |
| LP16 | 0.05 | 0.72 | 0.018 | 0.12 | 0.061 | −18.6 | −40.8 | 0.665 | 3.13 | 21.7 |
| LP17 | 0.07 | 0.69 | 0.017 | 0.11 | 0.088 | −18.4 | −43.5 | 0.630 | 5.44 | 23.1 |
| LP18 | 0.09 | 0.68 | 0.017 | 0.09 | 0.14 | −18.4 | −42.7 | 0.518 | 8.82 | 23.3 |
| LP19 | 0.11 | 0.67 | 0.017 | 0.21 | 0.053 | −19.7 | −50.9 | 1.22 | 11.4 | 45.5 |
| LP20 | 0.08 | 0.69 | 0.018 | 0.07 | 0.15 | −19.8 | −41.2 | 0.418 | 7.69 | 19.4 |
| LP21 | 0.13 | 0.65 | 0.016 | 0.38 | 0.023 | −23.3 | −54.8 | 2.10 | 7.83 | 66.5 |
| LP22 | 0.14 | 0.66 | 0.016 | 0.41 | 0.011 | −24.2 | −55.6 | 2.11 | 8.20 | 67.3 |
| LP23 | 0.15 | 0.65 | 0.017 | 0.17 | 0.081 | −26.7 | −44.6 | 0.892 | 13.7 | 38.7 |
| LP24 | 0.14 | 0.65 | 0.016 | 0.35 | 0.016 | −23.9 | −55.6 | 1.92 | 13.3 | 67.2 |
| LP25 | 0.12 | 0.66 | 0.017 | 0.34 | 0.018 | −21.9 | −54.2 | 1.90 | 13.1 | 66.3 |
| LP26 | 0.16 | 0.66 | 0.017 | 0.45 | 0.013 | −21.1 | −56.7 | 2.65 | 20.4 | 94.6 |
| LP27 | 0.09 | 0.65 | 0.016 | 0.11 | 0.16 | −18.0 | −50.0 | 0.772 | 7.18 | 28.8 |
| LP28 | 0.11 | 0.66 | 0.016 | 0.43 | 0.011 | −21.9 | −57.3 | 2.49 | 14.7 | 84.5 |
| LP29 | 0.08 | 0.65 | 0.016 | 0.09 | 0.11 | −18.3 | −47.4 | 0.545 | 11.6 | 26.9 |
| LP30 | 0.07 | 0.65 | 0.016 | 0.11 | 0.14 | −18.9 | −47.1 | 0.772 | 11.7 | 33.3 |
| ID | E | N | w.c.d. | T | pH | Eh | HCO3 | Cl | SO4 | Na | K | Ca | Mg | NH4 | NO3 | F | Br | TDS | δ18O−H2O | δD−H2O | δ13C−TDIC | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| spring water | SPW | 609910 | 4853354 | 0 | 15.4 | 7.77 | 645 | 263 | 40 | 13 | 24 | 1.7 | 85 | 6.2 | 0.04 | 6.1 | 0.26 | 0.11 | 439 | −6.07 | −35.1 | −14.75 |
| tributary channels | TCW1 | 609028 | 4854555 | 2 | 17.21 | 8.41 | 117 | 206 | 525 | 395 | 361 | 15 | 141 | 61 | 0.13 | 0.39 | 0.52 | 1.7 | 1707 | −13.6 | ||
| TCW2 | 608185 | 4852544 | 1 | 14.3 | 7.85 | 223 | 451 | 726 | 450 | 396 | 8.5 | 222 | 91 | 0.81 | 11 | 0.45 | 3.3 | 2360 | −3.86 | −23.8 | −13.03 | |
| TCW3 | 609931 | 4853285 | 1 | 14.1 | 7.93 | 221 | 222 | 811 | 222 | 423 | 20 | 110 | 68 | 0.16 | 0.2 | 0.39 | 3 | 1880 | −1.68 | −13.3 | −12.9 | |
| TCW4 | 608460 | 4855717 | 0.5 | 16.5 | 8.11 | 209 | 166 | 20 | 281 | 10 | 0 | 141 | 11 | 0.05 | 6.3 | 0.39 | 0.06 | 636 | −6.33 | −36.7 | −13.79 | |
| TCW5 | 601245 | 4858083 | 1.5 | 16.3 | 8.16 | 201 | 274 | 246 | 178 | 141 | 11 | 115 | 26 | 1.72 | 31 | 0.5 | 0.73 | 1025 | −5.61 | −33.6 | −13.75 | |
| lake shores | LSW1 | 605253 | 4854472 | 3 | 16.1 | 8.1 | 209 | 212 | 981 | 449 | 622 | 26 | 132 | 99 | 0.32 | 0.21 | 0.33 | 2.5 | 2524 | −11.7 | ||
| LSW2 | 605329 | 4854662 | 3 | 15.9 | 8.18 | 179 | 185 | 998 | 433 | 611 | 26 | 128 | 97 | 0.25 | 0.26 | 0.61 | 2.6 | 2482 | −12.6 | |||
| LSW3 | 605053 | 4854417 | 3 | 17.1 | 8.25 | 127 | 209 | 1040 | 431 | 627 | 26 | 140 | 100 | 0.2 | 0.21 | 0.42 | 2.4 | 2576 | −12.9 | |||
| LSW4 | 604864 | 4854403 | 1.8 | 17 | 8.45 | 135 | 198 | 984 | 452 | 614 | 25 | 131 | 98 | 0.23 | 0.23 | 0.45 | 2.4 | 2505 | −0.99 | −10.1 | −13.5 | |
| LSW5 | 606212 | 4852985 | 0.8 | 14.5 | 7.86 | 164 | 224 | 993 | 366 | 610 | 31 | 138 | 95 | 0.24 | 1.3 | 0.61 | 2.9 | 2462 | −1.22 | −10.7 | −12.4 | |
| LSW6 | 607547 | 4853088 | 0.8 | 14.5 | 7.87 | 223 | 240 | 989 | 399 | 591 | 27 | 136 | 94 | 0.23 | 0.13 | 0.37 | 4.1 | 2481 | −12.84 | |||
| LSW7 | 608772 | 4854627 | 1 | 16.1 | 7.98 | −179 | 231 | 1078 | 297 | 618 | 32 | 138 | 98 | 0.3 | 0 | 0.45 | 4.3 | 2497 | −1.37 | −11.1 | −13.06 | |
| LSW8 | 608132 | 4855571 | 1 | 15.4 | 7.82 | 184 | 281 | 1125 | 267 | 593 | 27 | 152 | 96 | 0.48 | 0.24 | 0.71 | 3.9 | 2546 | −1.8 | −14.4 | −13.82 | |
| LSW9 | 605717 | 4854577 | 2.5 | 14.8 | 8.22 | 195 | 209 | 1039 | 246 | 612 | 26 | 130 | 96 | 0.14 | 0.15 | 0.42 | 3.8 | 2363 | −14.23 | |||
| emissary channels | ECW1 | 603052 | 4858611 | 1 | 17.4 | 8.36 | 133 | 211 | 1730 | 495 | 1020 | 42 | 172 | 147 | 0.13 | 0.28 | 0.44 | 3.6 | 3822 | −14.1 | ||
| ECW2 | 606877 | 4856416 | 0.7 | 16 | 7.89 | 210 | 219 | 1166 | 302 | 611 | 28 | 135 | 92 | 0.11 | 0.38 | 0.66 | 4 | 2558 | −2.4 | −16.8 | −13.13 | |
| ECW3 | 605588 | 4857131 | 0.7 | 16.8 | 8.12 | 211 | 219 | 1515 | 304 | 763 | 47 | 150 | 118 | 0.18 | 0.36 | 0.54 | 5 | 3122 | −1.28 | −11.6 | −12.55 | |
| ECW4 | 602446 | 4858415 | 1 | 16.5 | 8.29 | 202 | 219 | 1672 | 306 | 829 | 34 | 151 | 120 | 0.11 | 0.22 | 0.56 | 6.4 | 3338 | −12.7 | |||
| ECW5 | 601273 | 4857944 | 1 | 16.2 | 8.37 | 208 | 227 | 1555 | 350 | 810 | 29 | 152 | 122 | 0.14 | 0.39 | 0.41 | 6.5 | 3252 | −13.18 | |||
| ECW6 | 600820 | 4857784 | 1 | 16.1 | 8.2 | 196 | 231 | 1490 | 297 | 753 | 32 | 146 | 113 | 0.13 | 0.75 | 0.69 | 5.2 | 3069 | −12.25 | |||
| ECW7 | 603180 | 4857119 | 0.7 | 16.3 | 8.38 | 193 | 241 | 1368 | 273 | 735 | 33 | 139 | 111 | 0.18 | 0.01 | 0.63 | 5.2 | 2906 | −12.17 | |||
| ECW8 | 603748 | 4856001 | 0.7 | 15.7 | 8.15 | 201 | 323 | 727 | 121 | 389 | 22 | 116 | 64 | 0.38 | 17 | 0.51 | 2.6 | 1782 | −3.87 | −24.1 | −14.51 | |
| ECW9 | 603842 | 4855965 | 0.7 | 16.7 | 8.1 | 219 | 244 | 1274 | 263 | 664 | 29 | 138 | 103 | 0.15 | 0.42 | 0.46 | 5.5 | 2722 | −1.38 | −11.6 | −12.98 | |
| main basin (Massaciuccoli lake) | OMW1 (0 m) | 606696 | 4854546 | 3 | 14.1 | 7.77 | 214 | 220 | 996 | 320 | 590 | 28 | 129 | 90 | 0.22 | 0.02 | 0.39 | 4.1 | 2378 | −1.22 | −11.2 | −13.78 |
| OMW1 (1 m) | 606696 | 4854546 | 13.9 | 7.69 | 214 | 212 | 1245 | 331 | 634 | 23 | 143 | 99 | 0.23 | 0.04 | 0.32 | 4.2 | 2692 | −16.79 | ||||
| OMW1 (2 m) | 606696 | 4854546 | 13.5 | 7.57 | 236 | 208 | 1196 | 255 | 639 | 22 | 138 | 101 | 0.23 | 0.01 | 0.39 | 5.1 | 2565 | −16.68 | ||||
| OMW1 (3 m) | 606696 | 4854546 | 13.5 | 7.39 | 230 | 213 | 1312 | 307 | 639 | 27 | 140 | 100 | 0.25 | 0.13 | 0.44 | 5.4 | 2744 | −16.71 | ||||
| OMW2 (0 m) | 606275 | 4854208 | 2.5 | 13.7 | 8.34 | 193 | 214 | 1261 | 296 | 629 | 28 | 138 | 99 | 0.2 | 0.01 | 0.48 | 4.8 | 2670 | −1.1 | −10.8 | −13.91 | |
| OMW2 (1 m) | 606275 | 4854208 | 13.7 | 8.26 | 183 | 213 | 1225 | 285 | 632 | 25 | 143 | 99 | 0.2 | 0.04 | 0.56 | 4.3 | 2627 | −13.34 | ||||
| OMW2 (2 m) | 606275 | 4854208 | 13.9 | 7.94 | 180 | 214 | 1213 | 289 | 638 | 24 | 139 | 100 | 0.23 | 0.12 | 0.51 | 4.1 | 2622 | −13.51 | ||||
| OMW2 (2.5 m) | 606275 | 4854208 | 13.9 | 7.88 | 190 | 212 | 1209 | 284 | 626 | 23 | 138 | 98 | 0.13 | 0.22 | 0.47 | 3.9 | 2595 | −13.55 | ||||
| Cava Sisa | OSW1 (0 m) | 604945 | 4854675 | 16 | 15.1 | 8.2 | 188 | 213 | 1042 | 313 | 616 | 27 | 128 | 96 | 0.15 | 0.24 | 0.45 | 4.4 | 2440 | −1.17 | −10.7 | −13.49 |
| OSW1 (1 m) | 604945 | 4854675 | 14.8 | 8.14 | 164 | 212 | 1206 | 275 | 608 | 19 | 129 | 95 | 0.11 | 0.07 | 0.39 | 4.3 | 2549 | −13.25 | ||||
| OSW1 (2 m) | 604945 | 4854675 | 14.8 | 8.08 | 149 | 212 | 1171 | 267 | 611 | 24 | 130 | 95 | 0.16 | 0.2 | 0.33 | 4.2 | 2515 | −13.86 | ||||
| OSW1 (4 m) | 604945 | 4854675 | 14.7 | 8.12 | 138 | 211 | 1176 | 268 | 614 | 25 | 130 | 96 | 0.11 | 0.2 | 0.39 | 3.8 | 2525 | −14.87 | ||||
| OSW1 (6 m) | 604945 | 4854675 | 14.7 | 8.14 | 137 | 217 | 1247 | 318 | 624 | 30 | 133 | 96 | 0.11 | 0.35 | 0.45 | 4.1 | 2670 | −14.59 | ||||
| OSW1 (8 m) | 604945 | 4854675 | 14.7 | 8.14 | 146 | 207 | 1169 | 266 | 622 | 21 | 131 | 97 | 0.14 | 0.15 | 0.46 | 3.8 | 2518 | −14.30 | ||||
| OSW1 (10 m) | 604945 | 4854675 | 14.5 | 8.16 | −11 | 212 | 1252 | 288 | 601 | 24 | 128 | 95 | 0.14 | 0.2 | 0.49 | 3.9 | 2605 | −16.33 | ||||
| OSW1 (12 m) | 604945 | 4854675 | 10.9 | 7.47 | −202 | 272 | 1160 | 268 | 621 | 91 | 138 | 94 | 1.5 | 0.1 | 0.54 | 3.6 | 2650 | −16.68 | ||||
| OSW1 (14 m) | 604945 | 4854675 | 10 | 7.35 | −212 | 297 | 1170 | 267 | 577 | 29 | 143 | 92 | 2.03 | 0.08 | 0.57 | 3.7 | 2581 | −16.70 | ||||
| OSW1 (16 m) | 604945 | 4854675 | 9.9 | 7.17 | −215 | 314 | 1101 | 268 | 574 | 22 | 138 | 92 | 2.7 | 0.36 | 0.66 | 3.4 | 2516 | −17.01 | ||||
| OSW2 | 605069 | 4854569 | 15 | 16.5 | 8.2 | 163 | 209 | 1050 | 444 | 640 | 29 | 135 | 100 | 0.18 | 0.18 | 0.42 | 2.6 | 2610 | −13.1 |
| ID | CO2 | N2 | Ar | CH4 | O2 | δ13C-CO2 | δ13C-CH4 | ΦCH4 | ΦCO2 | ΦCO2eq |
|---|---|---|---|---|---|---|---|---|---|---|
| SPW | 0.01 | 0.59 | 0.014 | 0.005 | 0.25 | −13.0 | −33.6 | 0.034 | −3.03 | −2.07 |
| TCW1 | 0.06 | 0.62 | 0.015 | 0.012 | 0.16 | −15.6 | −44.5 | 0.087 | 25.7 | 28.2 |
| TCW2 | 0.02 | 0.55 | 0.013 | 0.0076 | 0.19 | −17.1 | −35.9 | 0.050 | 0.500 | 1.90 |
| TCW3 | 0.03 | 0.54 | 0.013 | 0.014 | 0.18 | −18.5 | −45.6 | 0.092 | 3.78 | 6.35 |
| TCW4 | 0.01 | 0.65 | 0.016 | 0.0058 | 0.31 | −13.5 | −36.1 | 0.041 | −3.49 | −2.33 |
| TCW5 | 0.04 | 0.65 | 0.016 | 0.017 | 0.18 | −15.4 | −43.6 | 0.120 | 7.39 | 10.7 |
| LSW1 | 0.04 | 0.58 | 0.014 | 0.011 | 0.19 | −13.4 | −40.4 | 0.077 | 6.82 | 8.98 |
| LSW2 | 0.04 | 0.55 | 0.014 | 0.015 | 0.16 | −15.9 | −36.1 | 0.104 | 10.5 | 13.5 |
| LSW3 | 0.06 | 0.63 | 0.015 | 0.021 | 0.15 | −9.2 | −48.7 | 0.152 | 20.6 | 24.8 |
| LSW4 | 0.09 | 0.54 | 0.014 | 0.016 | 0.12 | −22.0 | −49.4 | 0.116 | 39.9 | 43.1 |
| LSW5 | 0.09 | 0.63 | 0.016 | 0.026 | 0.15 | −16.0 | −49.3 | 0.172 | 17.2 | 22.1 |
| LSW6 | 0.08 | 0.57 | 0.015 | 0.025 | 0.17 | −17.5 | −50.1 | 0.166 | 15.7 | 20.3 |
| LSW7 | 0.15 | 0.63 | 0.016 | 0.23 | 0.056 | −18.9 | −60.4 | 1.61 | 43.9 | 89.0 |
| LSW8 | 0.06 | 0.66 | 0.016 | 0.019 | 0.12 | −15.4 | −48.8 | 0.130 | 11.3 | 15.0 |
| LSW9 | 0.05 | 0.63 | 0.016 | 0.0065 | 0.15 | −15.3 | −40.8 | 0.044 | 3.89 | 5.11 |
| ECW1 | 0.10 | 0.56 | 0.013 | 0.016 | 0.13 | −18.8 | −47.6 | 0.117 | 41.8 | 45.1 |
| ECW2 | 0.07 | 0.64 | 0.015 | 0.012 | 0.15 | −17.4 | −45.5 | 0.084 | 15.8 | 18.1 |
| ECW3 | 0.06 | 0.61 | 0.015 | 0.016 | 0.17 | −16.5 | −45.1 | 0.115 | 15.3 | 18.5 |
| ECW4 | 0.03 | 0.56 | 0.014 | 0.0085 | 0.16 | −13.4 | −37.3 | 0.060 | 6.79 | 8.48 |
| ECW5 | 0.03 | 0.57 | 0.015 | 0.011 | 0.19 | −14.8 | −42.3 | 0.077 | 5.09 | 7.26 |
| ECW6 | 0.03 | 0.63 | 0.015 | 0.015 | 0.15 | −16.6 | −43.4 | 0.105 | 3.01 | 5.96 |
| ECW7 | 0.09 | 0.64 | 0.016 | 0.011 | 0.15 | −14.7 | −41.9 | 0.078 | 35.5 | 37.7 |
| ECW8 | 0.01 | 0.61 | 0.015 | 0.016 | 0.18 | −13.2 | −44.8 | 0.111 | −2.84 | 0.261 |
| ECW9 | 0.07 | 0.61 | 0.016 | 0.012 | 0.19 | −17.0 | −46.6 | 0.086 | 20.0 | 22.4 |
| OMW1 (0 m) | 0.02 | 0.55 | 0.014 | 0.0035 | 0.19 | −13.4 | −44.5 | 0.023 | −0.03 | 0.613 |
| OMW1 (1 m) | 0.04 | 0.58 | 0.014 | 0.0056 | 0.18 | −12.7 | −44.6 | |||
| OMW1 (2 m) | 0.05 | 0.64 | 0.016 | 0.0071 | 0.15 | −13.0 | −44.2 | |||
| OMW1 (3 m) | 0.08 | 0.63 | 0.016 | 0.0088 | 0.13 | −13.6 | −47.8 | |||
| OMW2 (0 m) | 0.02 | 0.57 | 0.014 | 0.0031 | 0.18 | −12.7 | −43.3 | 0.020 | 1.09 | 1.65 |
| OMW2 (1 m) | 0.04 | 0.59 | 0.015 | 0.0049 | 0.18 | −13.7 | −43.5 | |||
| OMW2 (2 m) | 0.06 | 0.54 | 0.014 | 0.0063 | 0.15 | −13.9 | −45.6 | |||
| OMW2 (2.5 m) | 0.10 | 0.56 | 0.014 | 0.0078 | 0.14 | −14.2 | −47.9 | |||
| OSW1 (0 m) | 0.07 | 0.62 | 0.016 | 0.0087 | 0.15 | −14.6 | −43.7 | 0.059 | 5.95 | 7.60 |
| OSW1 (1 m) | 0.09 | 0.59 | 0.014 | 0.016 | 0.13 | −15.1 | −48.9 | |||
| OSW1 (2 m) | 0.12 | 0.56 | 0.014 | 0.018 | 0.095 | −15.2 | −49.2 | |||
| OSW1 (4 m) | 0.13 | 0.55 | 0.013 | 0.035 | 0.071 | −15.1 | −50.3 | |||
| OSW1 (6 m) | 0.15 | 0.58 | 0.014 | 0.042 | 0.056 | −15.9 | −52.1 | |||
| OSW1 (8 m) | 0.16 | 0.62 | 0.015 | 0.095 | 0.011 | −16.6 | −53.6 | |||
| OSW1 (10 m) | 0.16 | 0.59 | 0.014 | 0.21 | 0 | −18.2 | −60.5 | |||
| OSW1 (12 m) | 0.18 | 0.63 | 0.015 | 0.28 | 0 | −19.1 | −61.3 | |||
| OSW1 (14 m) | 0.21 | 0.61 | 0.015 | 0.35 | 0 | −19.5 | −61.1 | |||
| OSW1 (16 m) | 0.23 | 0.58 | 0.014 | 0.56 | 0 | −19.6 | −63.4 | |||
| OSW2 | 0.05 | 0.61 | 0.015 | 0.019 | 0.17 | −15.1 | −45.0 | 0.135 | 13.1 | 16.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venturi, S.; Tassi, F.; Cabassi, J.; Randazzo, A.; Lazzaroni, M.; Capecchiacci, F.; Vietina, B.; Vaselli, O. Exploring Methane Emission Drivers in Wetlands: The Cases of Massaciuccoli and Porta Lakes (Northern Tuscany, Italy). Appl. Sci. 2021, 11, 12156. https://0-doi-org.brum.beds.ac.uk/10.3390/app112412156
Venturi S, Tassi F, Cabassi J, Randazzo A, Lazzaroni M, Capecchiacci F, Vietina B, Vaselli O. Exploring Methane Emission Drivers in Wetlands: The Cases of Massaciuccoli and Porta Lakes (Northern Tuscany, Italy). Applied Sciences. 2021; 11(24):12156. https://0-doi-org.brum.beds.ac.uk/10.3390/app112412156
Chicago/Turabian StyleVenturi, Stefania, Franco Tassi, Jacopo Cabassi, Antonio Randazzo, Marta Lazzaroni, Francesco Capecchiacci, Barbara Vietina, and Orlando Vaselli. 2021. "Exploring Methane Emission Drivers in Wetlands: The Cases of Massaciuccoli and Porta Lakes (Northern Tuscany, Italy)" Applied Sciences 11, no. 24: 12156. https://0-doi-org.brum.beds.ac.uk/10.3390/app112412156