
Bioactive Properties of Breads Made with Sourdough of Hull-Less Barley or Conventional and Pigmented Wheat Flours

 , , , and
, , , and
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. The Flours and Ingredients
2.2. Sourdough Preparation
2.3. Bread-Making
2.4. Microbiological Analysis and Determination of pH and Total Titratable Acidity (TTA)
2.5. Extraction and Fractionation of Water-Soluble Extracts (WSEs)
2.6. Cell Cultures for Ex Vivo Assays
2.6.1. Cell Toxicity Assays
2.6.2. Recovery of Cell Viability
2.6.3. Intracellular Reactive Oxygen Species (ROS) Measurement
2.6.4. Immunoblot Analysis
2.7. Statistical Analysis
3. Results
3.1. Dough Analysis
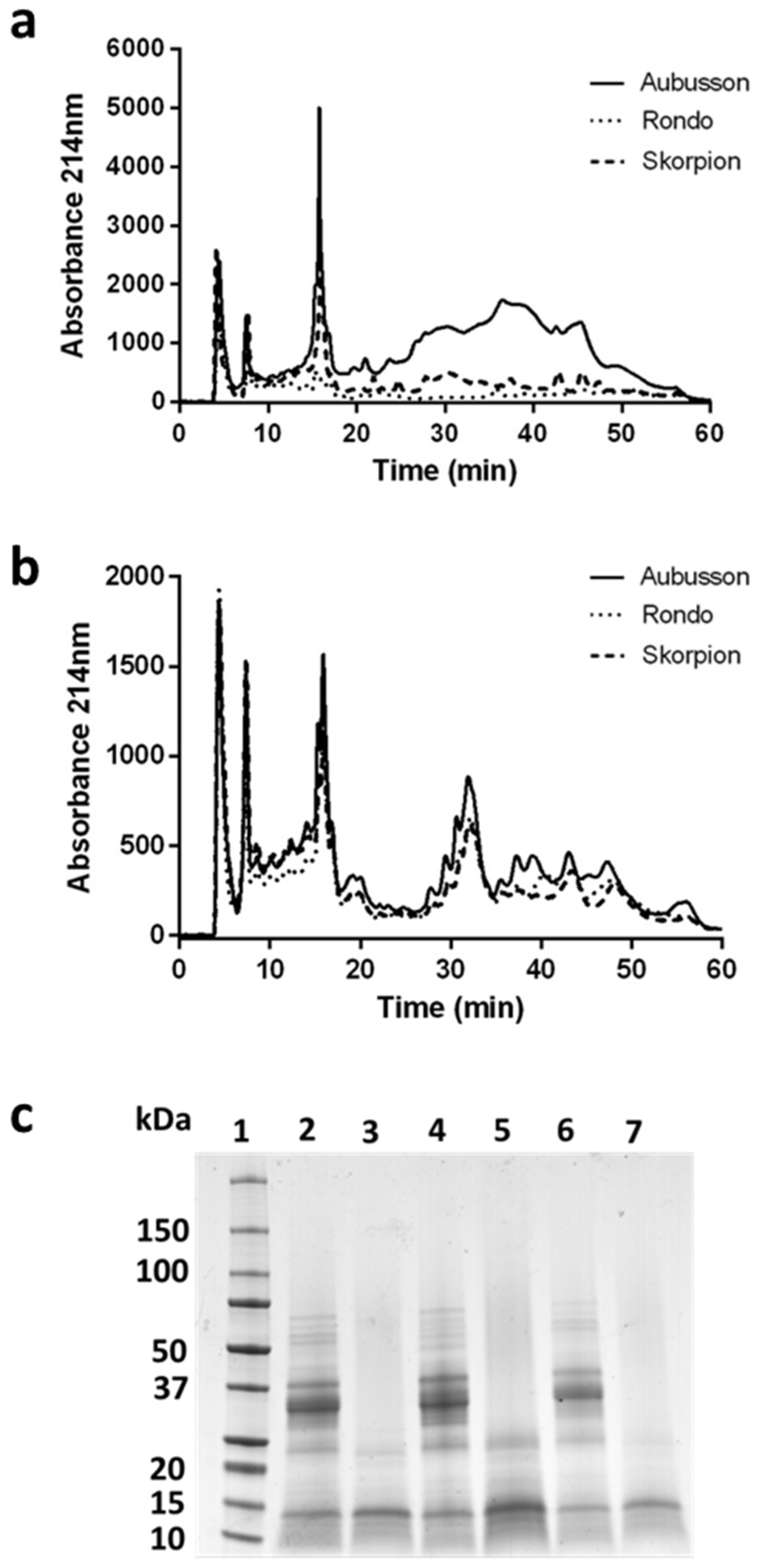
3.2. Low-Molecular-Weight Peptides Fractionation
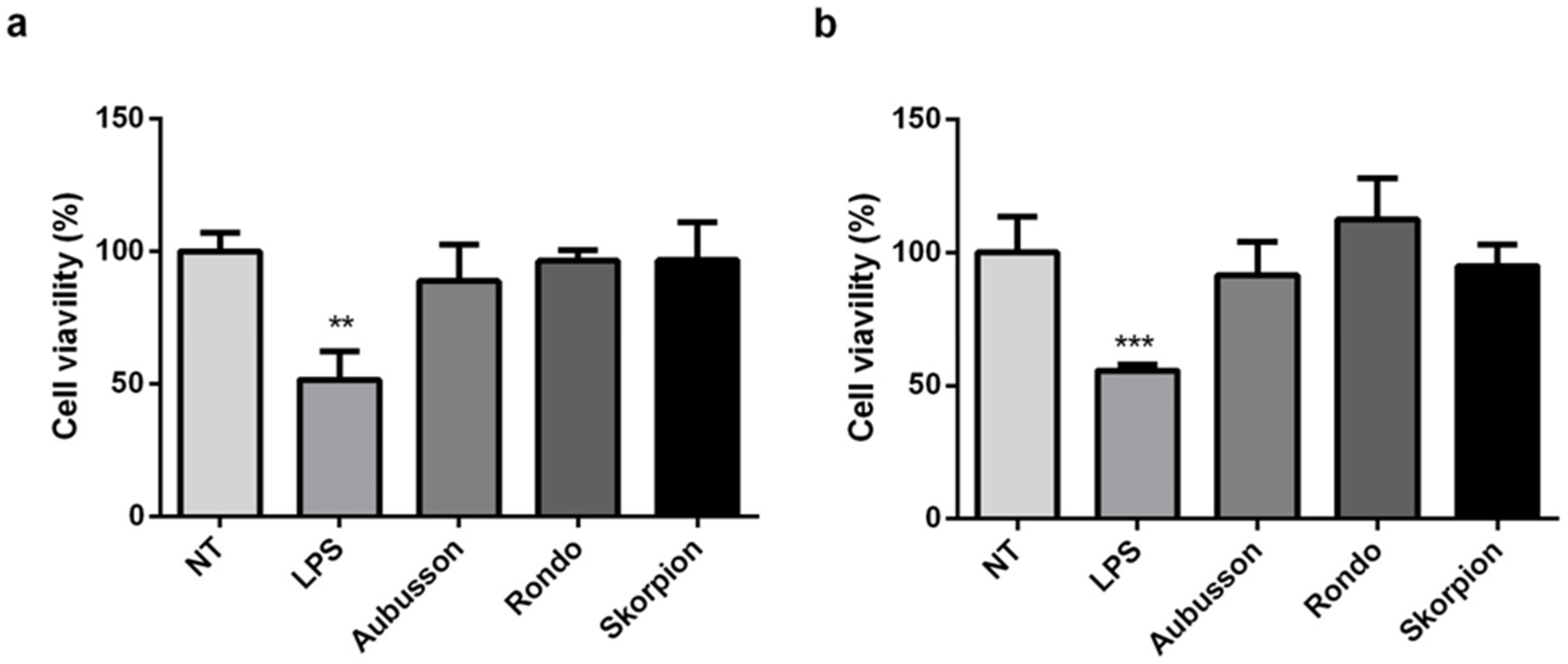
3.3. Cytotoxicity Assay
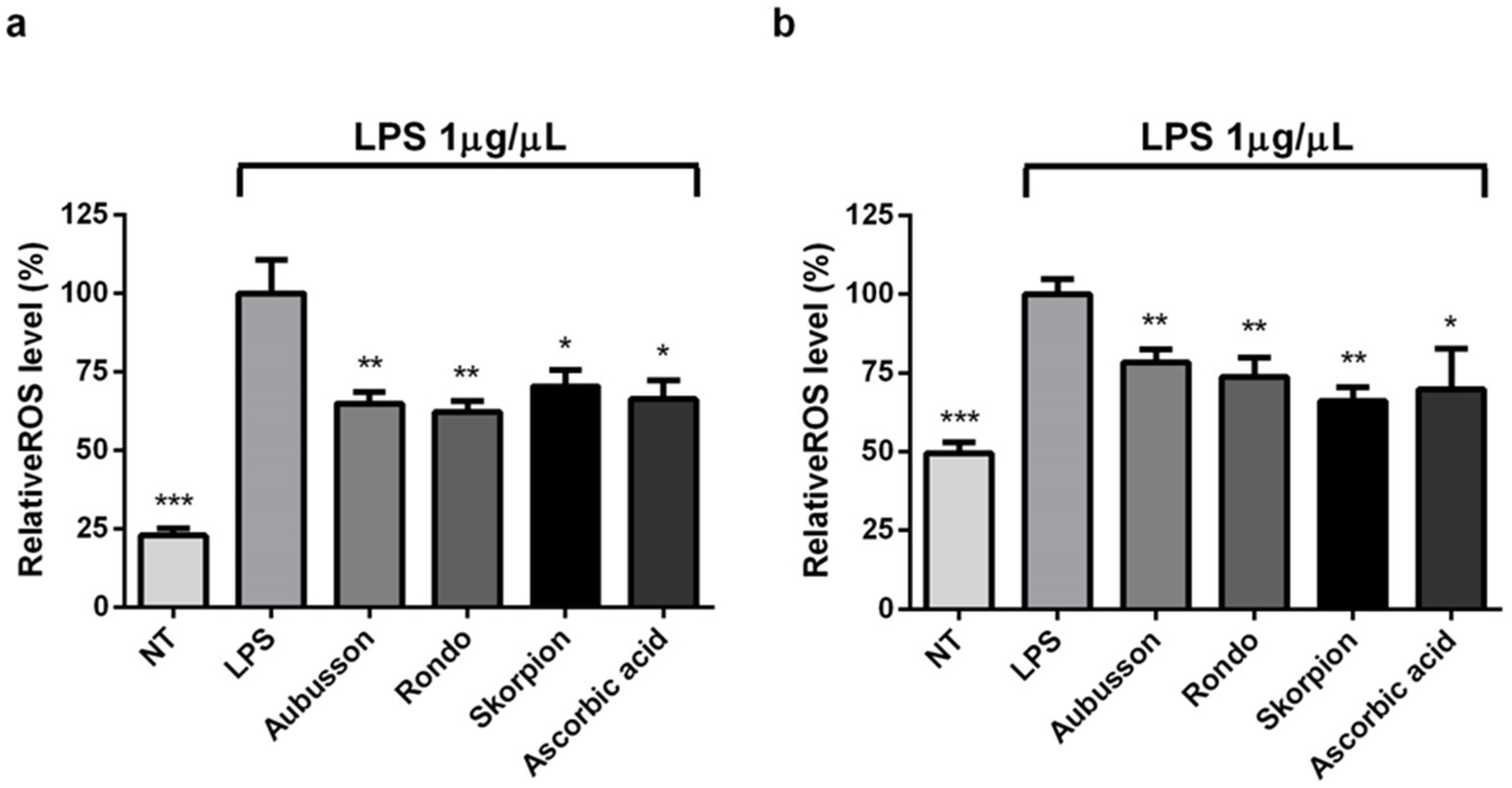
3.4. Antioxidant Activity of LMW Peptides
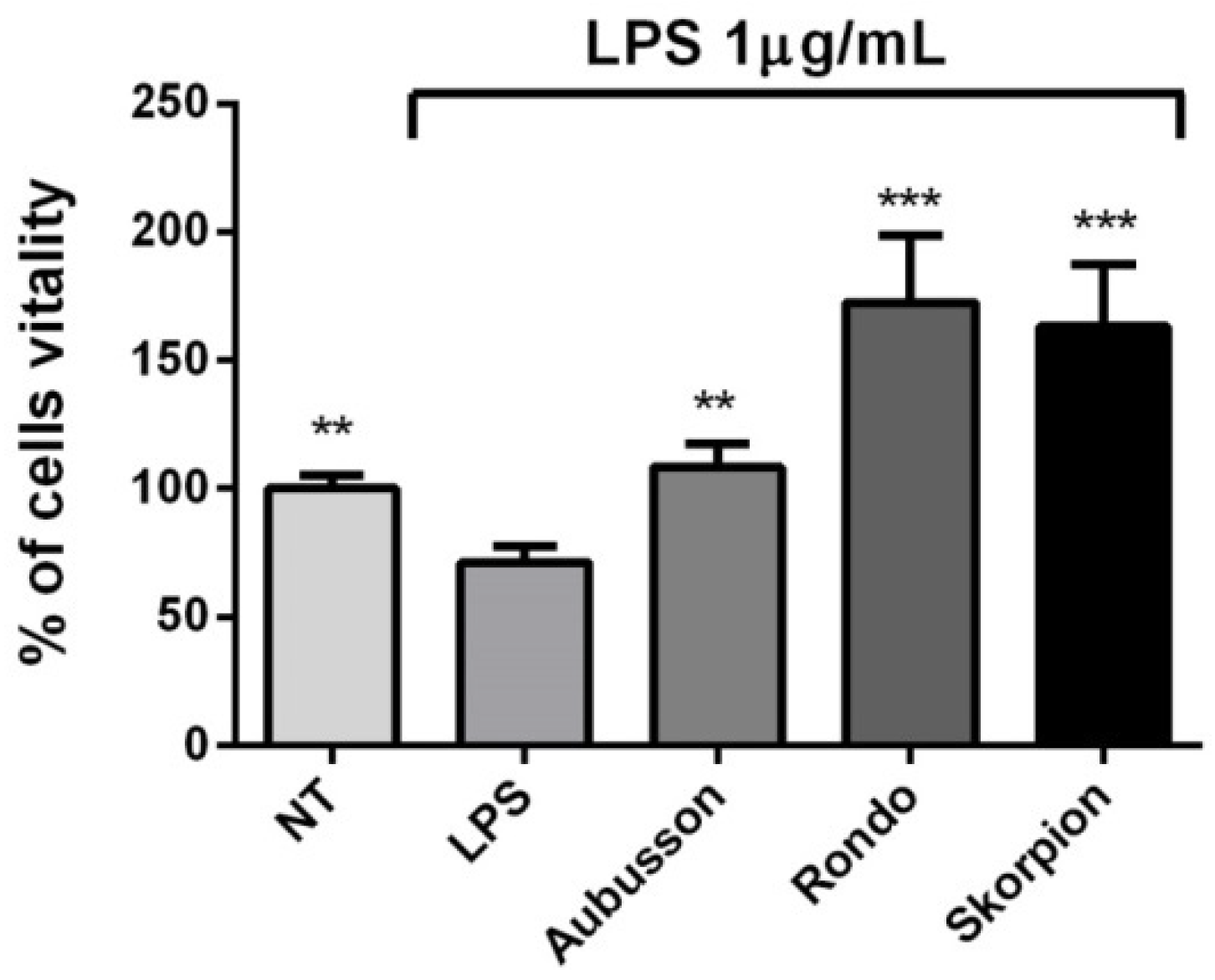
3.5. Recovery of Cell Vitality and Anti-Inflammatory Activity
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Calasso, M.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microbiol. 2019, 302, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Albiac, M.A.; Di Cagno, R.; Filannino, P.; Cantatore, V.; Gobbetti, M. How fructophilic lactic acid bacteria may reduce the FODMAPs content in wheat-derived baked goods: A proof of concept. Microb. Cell Fact. 2020, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Portincasa, P.; Montemurro, M.; Di Palo, D.M.; Lorusso, M.P.; De Angelis, M.; Bonfrate, L.; Genot, B.; Gobbetti, M. Sourdough fermented breads are more digestible than those started with baker’s yeast alone: An in vivo challenge dissecting distinct gastrointestinal responses. Nutrients 2019, 11, 2954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Schuppan, D.; Tovar, L.R.; Zevallos, V.; Loponen, J.; Gänzle, M. Sourdough fermentation degrades wheat alpha-amylase/trypsin inhibitor (ATI) and reduces pro-inflammatory activity. Foods 2020, 9, 943. [Google Scholar] [CrossRef]
- Galli, V.; Venturi, M.; Guerrini, S.; Blandino, M.; Luti, S.; Pazzagli, L.; Granchi, L. Antioxidant properties of sourdoughs made with whole grain flours of hull-less barley or conventional and pigmented wheat and by selected lactobacilli strains. Foods 2020, 9, 640. [Google Scholar] [CrossRef]
- De Vuyst, L.; Van Kerrebroeck, S.; Leroy, F. Microbial ecology and process technology of sourdough fermentation. Adv. Clin. Chem. 2017, 100, 49–160. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Loponen, J.; Gobbetti, M. Proteolysis in sourdough fermentations: Mechanisms and potential for improved bread quality. Trends Food Sci. Technol. 2008, 19, 513–521. [Google Scholar] [CrossRef]
- Galli, V.; Mazzoli, L.; Luti, S.; Venturi, M.; Guerrini, S.; Paoli, P.; Vincenzini, M.; Granchi, L.; Pazzagli, L. Effect of selected strains of lactobacilli on the antioxidant and anti-inflammatory properties of sourdough. Int. J. Food Microbiol. 2018, 286, 55–65. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Luti, S.; Mazzoli, L.; Ramazzotti, M.; Galli, V.; Venturi, M.; Marino, G.; Lehmann, M.; Guerrini, S.; Granchi, L.; Paoli, P.; et al. Antioxidant and anti-inflammatory properties of sourdoughs containing selected Lactobacilli strains are retained in breads. Food Chem. 2020, 322, 126710. [Google Scholar] [CrossRef]
- Hu, C.; Cai, Y.-Z.; Li, W.; Corke, H.; Kitts, D.D. Anthocyanin characterization and bioactivity assessment of a dark blue grained wheat (Triticum aestivum L. cv. Hedong Wumai) extract. Food Chem. 2007, 104, 955–961. [Google Scholar] [CrossRef]
- Gong, L.; Cao, W.; Chi, H.; Wang, J.; Zhang, H.; Liu, J.; Sun, B. Whole cereal grains and potential health effects: Involvement of the gut microbiota. Food Res. Int. 2018, 103, 84–102. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Whole grains and phenolic acids: A review on bioactivity, functionality, health benefits and bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef] [Green Version]
- Siebenhandl, S.; Grausgruber, H.; Pellegrini, N.; Del Rio, D.; Fogliano, V.; Pernice, R.; Berghofer, E. Phytochemical profile of main antioxidants in different fractions of purple and blue wheat, and black barley. J. Agric. Food Chem. 2007, 55, 8541–8547. [Google Scholar] [CrossRef]
- Bartl, P.; Albreht, A.; Skrt, M.; Tremlová, B.; Ošťádalová, M.; Šmejkal, K.; Vovk, I.; Ulrih, N.P. Anthocyanins in purple and blue wheat grains and in resulting bread: Quantity, composition, and thermal stability. Int. J. Food Sci. Nutr. 2015, 66, 514–519. [Google Scholar] [CrossRef]
- Pontonio, E.; Dingeo, C.; Di Cagno, R.; Blandino, M.; Gobbetti, M.; Rizzello, C.G. Brans from hull-less barley, emmer and pigmented wheat varieties: From by-products to bread nutritional improvers using selected lactic acid bacteria and xylanase. Int. J. Food Microbiol. 2020, 313, 108384. [Google Scholar] [CrossRef]
- Palla, M.; Agnolucci, M.; Calzone, A.; Giovannetti, M.; Di Cagno, R.; Gobbetti, M.; Rizzello, C.G.; Pontonio, E. Exploitation of autochthonous Tuscan sourdough yeasts as potential starters. Int. J. Food Microbiol. 2019, 302, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Milanović, V.; Osimani, A.; Garofalo, C.; Belleggia, L.; Maoloni, A.; Cardinali, F.; Mozzon, M.; Foligni, R.; Aquilanti, L.; Clementi, F. Selection of cereal-sourced lactic acid bacteria as candidate starters for the baking industry. PLoS ONE 2020, 15, e0236190. [Google Scholar] [CrossRef] [PubMed]
- Galli, V.; Venturi, M.; Pini, N.; Guerrini, S.; Granchi, L.; Vincenzini, M. Liquid and firm sourdough fermentation: Microbial robustness and interactions during consecutive backsloppings. LWT 2019, 105, 9–15. [Google Scholar] [CrossRef]
- Tolosa, L.; Donato, M.T.; Gómez-Lechón, M.J. General cytotoxicity assessment by means of the MTT assay. Methods Mol. Biol. 2015, 1250, 333–348. [Google Scholar]
- Feoktistova, M.; Geserick, P.; Leverkus, M. Crystal violet assay for determining viability of cultured cells. Cold Spring Harb. Protoc. 2016, 2016, 087379. [Google Scholar] [CrossRef] [PubMed]
- Ni, D.; Xu, P.; Gallagher, S. Immunoblotting and immunodetection. Curr. Protoc. Cell Biol. 2017, 74, 6.2.1–6.2.37. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Ahsan, A.U.; Wani, A.; Khajuria, V.; Nazir, L.A.; Sharma, S.; Bhagat, A.; Sharma, P.R.; Bhardwaj, S.; Peerzada, K.J.; et al. The amino analogue of β-boswellic acid efficiently attenuates the release of pro-inflammatory mediators than its parent compound through the suppression of NF-κB/IκBα signalling axis. Cytokine 2018, 107, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Milanovic, M.; Kracht, M.; Schmitz, M.L. The cytokine-induced conformational switch of nuclear factor κB p65 is mediated by p65 phosphorylation. Biochem. J. 2014, 457, 401–413. [Google Scholar] [CrossRef]
- Xue, Q.; Yan, Y.; Zhang, R.; Xiong, H. Regulation of iNOS on immune cells and its role in diseases. Int. J. Mol. Sci. 2018, 19, 3805. [Google Scholar] [CrossRef] [Green Version]
- Ramyaa, P.; Krishnaswamy, R.; Padma, V.V. Quercetin modulates OTA-induced oxidative stress and redox signalling in HepG2 cells—up regulation of Nrf2 expression and down regulation of NF-κB and COX-2. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 681–692. [Google Scholar] [CrossRef]
- Pan, F.; Wang, L.; Cai, Z.; Wang, Y.; Wang, Y.; Guo, J.; Xu, X.; Zhang, X.; Guoyue, J. Soybean peptide QRPR activates Autophagy and attenuates the inflammatory response in the RAW264.7 cell model. Protein Pept. Lett. 2019, 26, 301–312. [Google Scholar] [CrossRef]
- More, G.K.; Makola, R.T. In-vitro analysis of free radical scavenging activities and suppression of LPS-induced ROS production in macrophage cells by Solanum sisymbriifolium extracts. Sci. Rep. 2020, 10, 6493. [Google Scholar] [CrossRef]
- Śliwka, L.; Wiktorska, K.; Suchocki, P.; Milczarek, M.; Mielczarek, S.; Lubelska, K.; Cierpiał, T.; Łyżwa, P.; Kiełbasiński, P.; Jaromin, A.; et al. The comparison of MTT and CVS assays for the assessment of anticancer agent interactions. PLoS ONE 2016, 11, e0155772. [Google Scholar] [CrossRef] [Green Version]
- Arango-Varela, S.S.; Luzardo-Ocampo, I.; Maldonado-Celis, M.E.; Campos-Vega, R. Andean berry (Vaccinium meridionale Swartz) juice in combination with Aspirin modulated anti-inflammatory markers on LPS-stimulated RAW 264.7 macrophages. Food Res. Int. 2020, 137, 109541. [Google Scholar] [CrossRef]
- Singh, B.P.; Aluko, R.E.; Hati, S.; Solanki, D. Bioactive peptides in the management of lifestyle-related diseases: Current trends and future perspectives. Crit. Rev. Food Sci. Nutr. 2021, 1–14. [Google Scholar] [CrossRef]
- Ustunol, Z. Applied Food Protein Chemistry; John Wiley & Sons, Ltd.: Chichester, UK, 2014; ISBN 9781118860588. [Google Scholar]
- Esfandi, R.; Walters, M.E.; Tsopmo, A. Antioxidant properties and potential mechanisms of hydrolyzed proteins and peptides from cereals. Heliyon 2019, 5, e01538. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S. Oxidative stress, inflammation, and disease. In Oxidative Stress and Biomaterials; Elsevier: Amsterdam, The Netherlands, 2016; pp. 35–58. [Google Scholar]
- Calcabrini, C.; Mancini, U.; De Bellis, R.; Frati, A.; Mastrogiacomo, A.R.; Annibalini, G.; Sestili, P.; Cucchiarini, L.; Stocchi, V.; Potenza, L. Protective role of Italian Juglans regia L. nut ethanolic extract in human keratinocytes under oxidative and inflammatory stress. Curr. Pharm. Biotechnol. 2018, 18, 925–934. [Google Scholar] [CrossRef]
- Reyes-Díaz, A.; Del-Toro-Sánchez, C.L.; Rodríguez-Figueroa, J.C.; Valdéz-Hurtado, S.; Wong-Corral, F.J.; Borboa-Flores, J.; González-Osuna, M.F.; Perez-Perez, L.M.; González-Vega, R.I. Legume proteins as a promising source of anti-inflammatory peptides. Curr. Protein Pept. Sci. 2019, 20, 1204–1217. [Google Scholar] [CrossRef]
- Koistinen, V.M.; Mattila, O.; Katina, K.; Poutanen, K.; Aura, A.-M.; Hanhineva, K. Metabolic profiling of sourdough fermented wheat and rye bread. Sci. Rep. 2018, 8, 5684. [Google Scholar] [CrossRef]
- Păcularu-Burada, B.; Georgescu, L.A.; Vasile, M.A.; Rocha, J.M.; Bahrim, G.-E. Selection of wild lactic acid bacteria strains as promoters of postbiotics in gluten-free sourdoughs. Microorganisms 2020, 8, 643. [Google Scholar] [CrossRef]
- Petrova, P.; Petrov, K. Lactic acid fermentation of cereals and Pseudocereals: Ancient nutritional biotechnologies with modern applications. Nutrients 2020, 12, 1118. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bread | |||
|---|---|---|---|
| Ingredients | Aubusson | Skorpion | Rondo |
| Sourdough (g) | 200 | 200 | 200 |
| Wheat flour Type 00 (g) | 985 | 985 | 1010 |
| Fermented bran (g) | 40 | 40 | 40 |
| Water (mL) | 815 | 815 | 790 |
| Dough yield | 180 | 180 | 180 |
| Dough | Final pH | ∆pH | Final TTA | ΔV/V0 × 100 | LAB (CFU/g) | Yeasts (CFU/g) |
|---|---|---|---|---|---|---|
| Aubusson | 4.37 ± 0.20 | 0.64 ± 0.11 | 5.10 ± 0.6 | 67 ± 5 b | (8.75 ± 0.83) × 108 | (5.83 ± 0.73) × 106 |
| Skorpion | 4.39 ± 0.25 | 0.70 ± 0.11 | 5.00 ± 0.1 | 94 ± 8 c | (8.15 ± 0.92) × 108 | (7.55 ± 0.82) × 106 |
| Rondo | 4.42 ± 0.16 | 0.67 ± 0.04 | 4.35 ± 0.5 | 11 ± 0 a | (8.65 ± 1.34) × 108 | (6.45 ± 0.64) × 106 |
| Flour | Bacteria and Yeasts | mg/mL | |
|---|---|---|---|
| Doughs | Aubusson | L. plantarum Fi13 L. brevis LD66 S. cerevisiae D20Y | 4.44 |
| Skorpion | L. plantarum Fi58 L. sanfranciscensis Fi33 S. cerevisiae L6Y | 5.17 | |
| Rondo | L. plantarum Fi13 L. sanfranciscensis Fi33 S. cerevisiae L6Y | 3.76 | |
| Breads | Aubusson | L. plantarum Fi13 L. brevis LD66 S. cerevisiae D20Y | 7.01 |
| Skorpion | L. plantarum Fi58 L. sanfranciscensis Fi33 S. cerevisiae L6Y | 5.39 | |
| Rondo | L. plantarum Fi13 L. sanfranciscensis Fi33 S. cerevisiae L6Y | 5.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luti, S.; Galli, V.; Venturi, M.; Granchi, L.; Paoli, P.; Pazzagli, L. Bioactive Properties of Breads Made with Sourdough of Hull-Less Barley or Conventional and Pigmented Wheat Flours. Appl. Sci. 2021, 11, 3291. https://0-doi-org.brum.beds.ac.uk/10.3390/app11073291
Luti S, Galli V, Venturi M, Granchi L, Paoli P, Pazzagli L. Bioactive Properties of Breads Made with Sourdough of Hull-Less Barley or Conventional and Pigmented Wheat Flours. Applied Sciences. 2021; 11(7):3291. https://0-doi-org.brum.beds.ac.uk/10.3390/app11073291
Chicago/Turabian StyleLuti, Simone, Viola Galli, Manuel Venturi, Lisa Granchi, Paolo Paoli, and Luigia Pazzagli. 2021. "Bioactive Properties of Breads Made with Sourdough of Hull-Less Barley or Conventional and Pigmented Wheat Flours" Applied Sciences 11, no. 7: 3291. https://0-doi-org.brum.beds.ac.uk/10.3390/app11073291