
Selenium-Containing Polysaccharides—Structural Diversity, Biosynthesis, Chemical Modifications and Biological Activity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Biologically Active Polysaccharides—Structure and Function
2.1. Bacterial Polysaccharides
2.2. Algal and Lichen Polysaccharides
2.3. Fungal Polysaccharides
2.4. Plant Polysaccharides
3. Organoselenium Compounds
3.1. Physiological Functions of Se
3.2. Selenium Species, Bioavailability, and Effects of Excess and Deficiency
3.3. Organoselenium Natural and Synthetic Compounds
3.4. Selenium in Therapy and Chemoprevention—Traditional and Next Generation of Se Supplements
4. Selenosugars: How Selenium May Be Bound to the Carbohydrate Structure
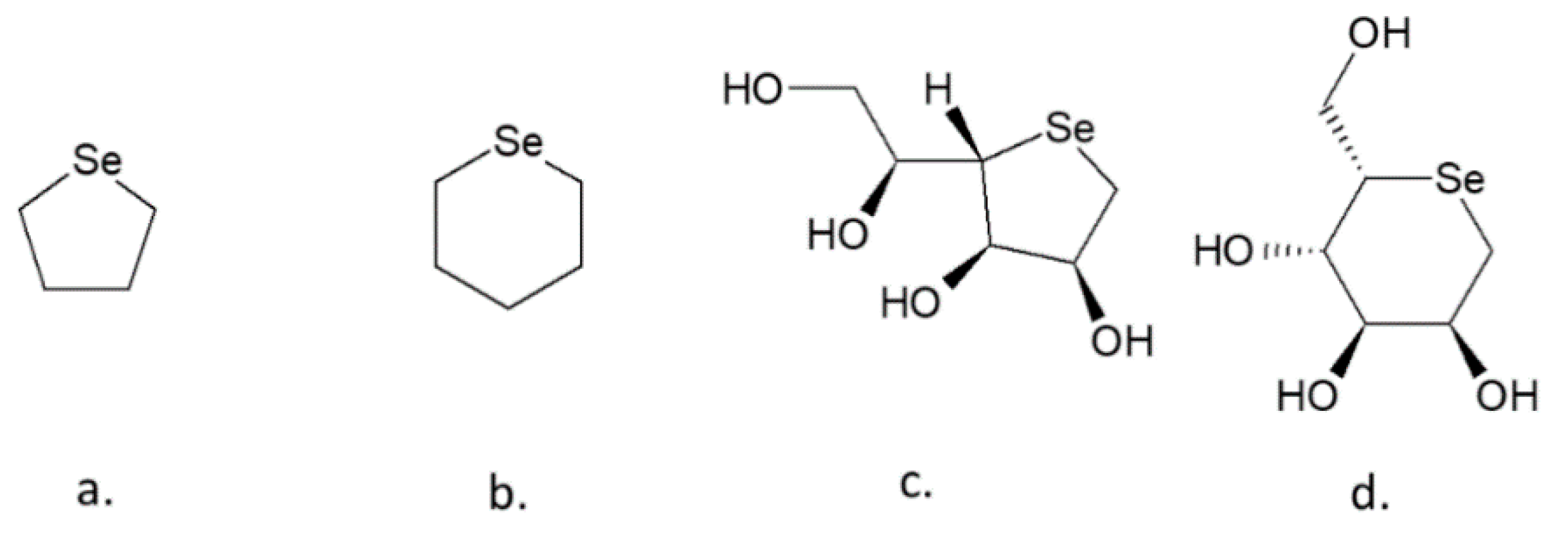
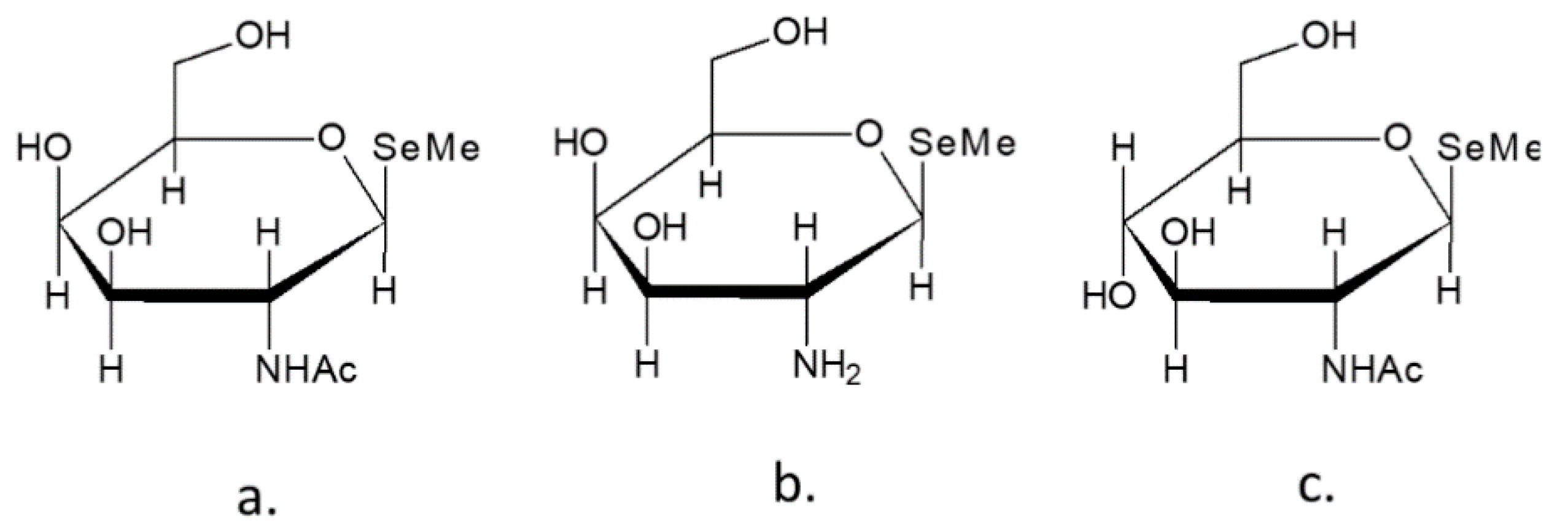
4.1. Selenopyranoses and Selenofuranoses
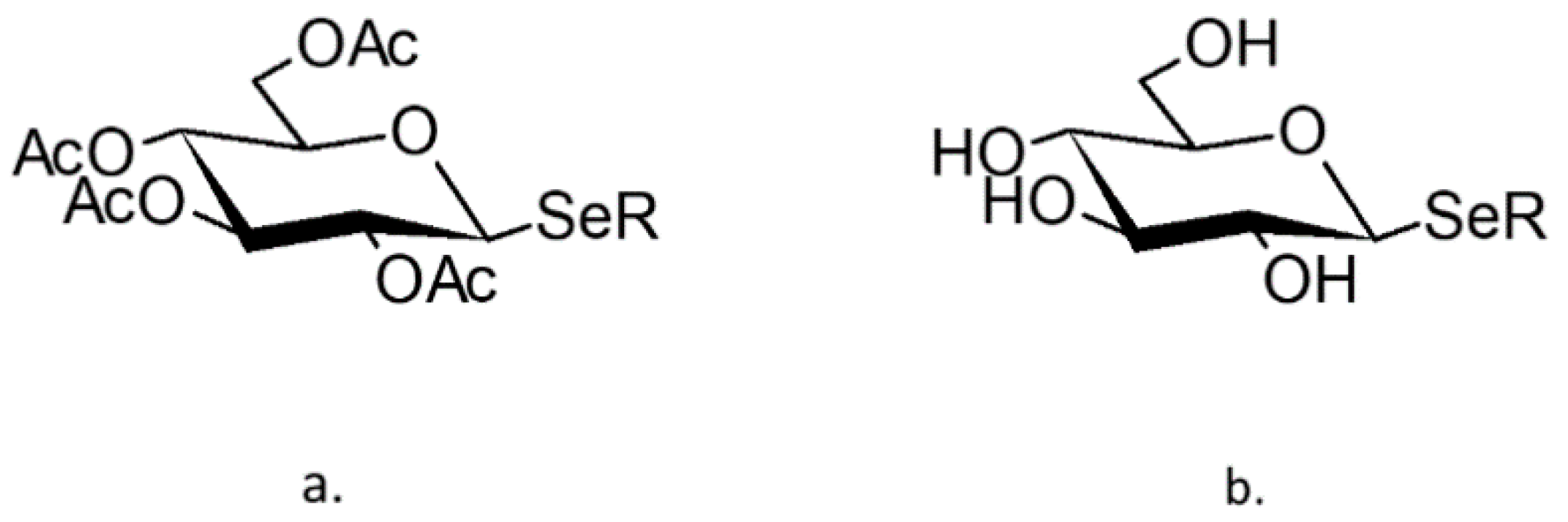
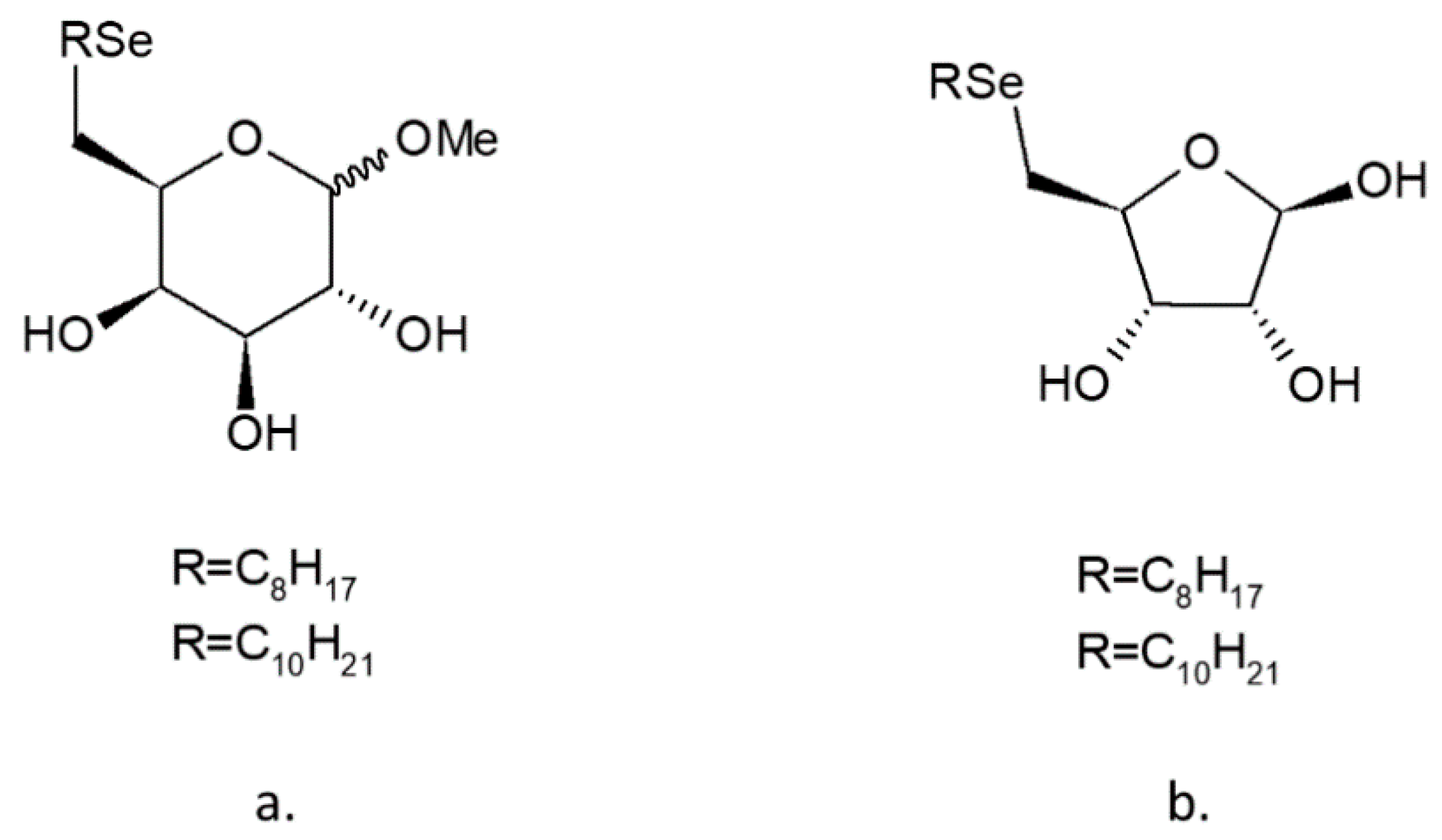
4.2. Se-Glycosides
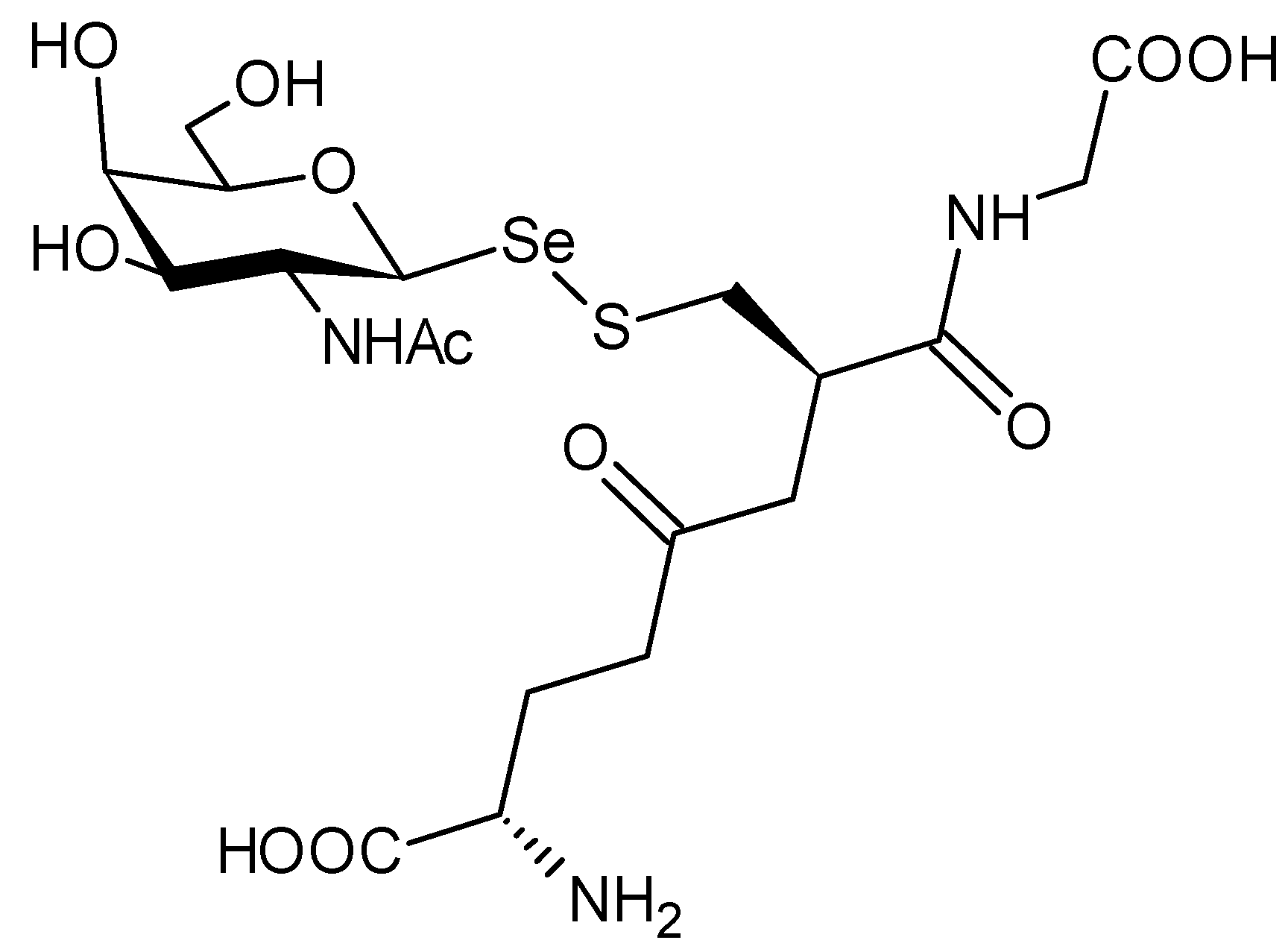
4.3. Se-Alkylselenides and Se-Arylselenides
4.4. Se-Esters
5. Selenium-Containing Polysaccharides
5.1. Divalent, Tetravalent, and Zerovalent Selenium in Polysaccharide Structure
5.2. Se-Glycosides and Selenopyranoses
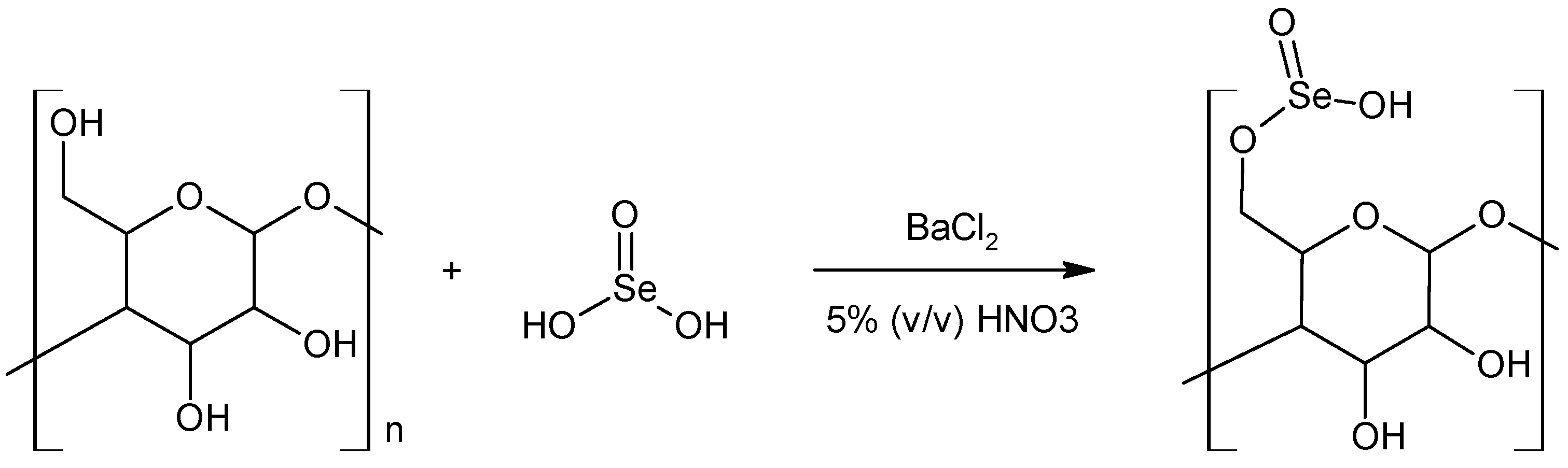
5.3. Se-Esters—Selenites and Diselenites of Polysaccharides
5.4. Polysaccharide Encapsulated Nanoselenium
6. Methods for Obtaining Selenium-Containing Polysaccharides
6.1. Extraction from Natural Products
6.2. Biotechnological Methods
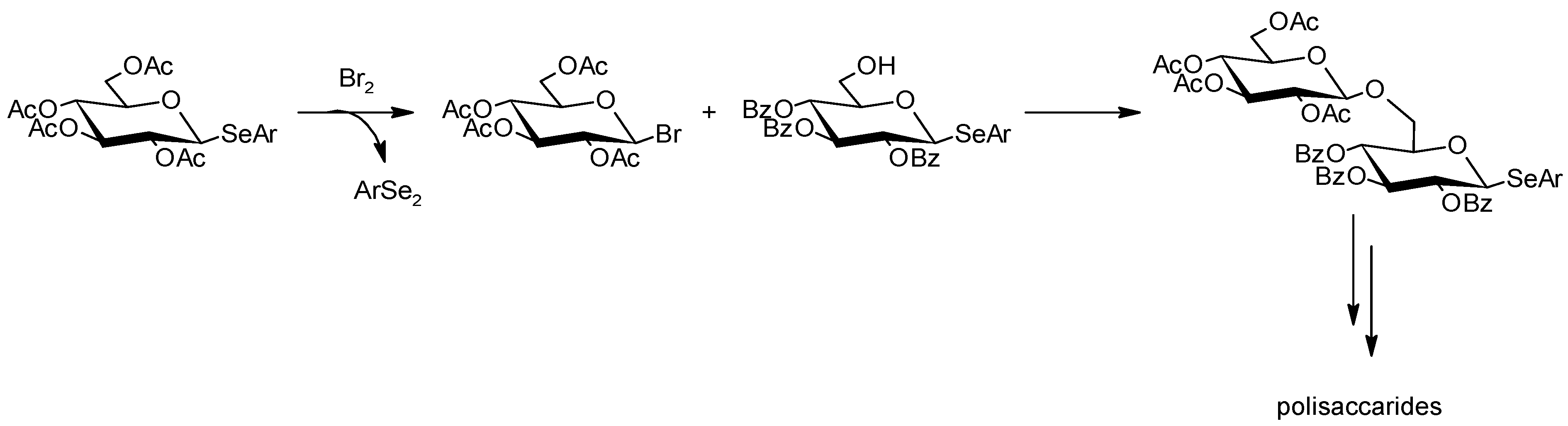
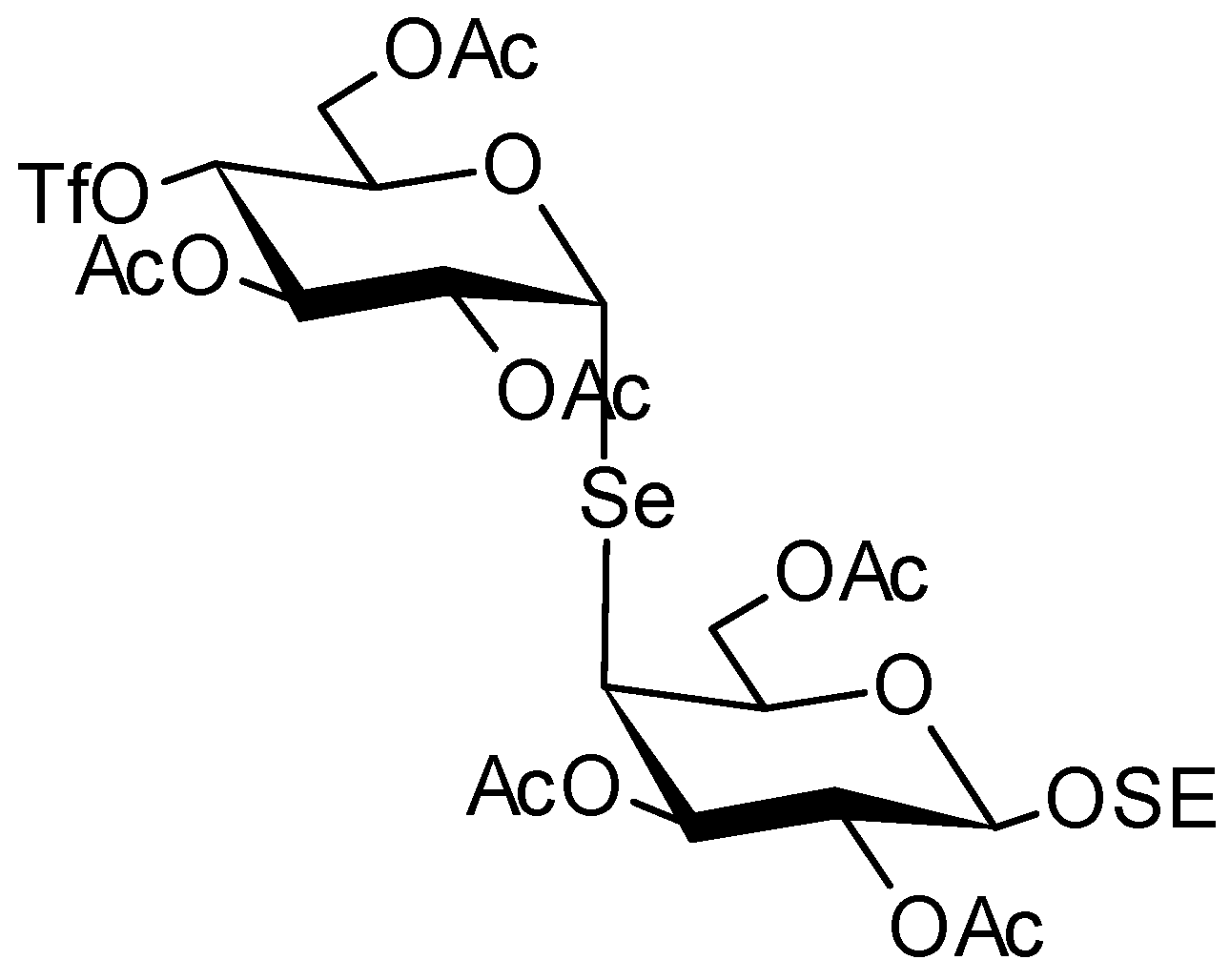
6.3. Chemical Modification of Natural Polysaccharides
6.4. Encapsulation of Nanoselenium Particles into Polysaccharides
7. Biological Activity of Se-Polysaccharides
7.1. Bacterial Se-Polysaccharides
7.2. Algal Se-Polysaccharides
7.3. Fungal Se-Polysaccharides
7.4. Plant Se-Polysaccharides
8. Se-Polysaccharides—Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cui, S.W. Food Carbohydrates: Chemistry, Physical Properties, and Applications; Taylor & Francis: Boca Raton, FL, USA, 2005; ISBN 978-0-8493-1574-9. [Google Scholar]
- Diener, M.; Adamcik, J.; Sánchez-Ferrer, A.; Jaedig, F.; Schefer, L.; Mezzenga, R. Primary, Secondary, Tertiary and Quaternary Structure Levels in Linear Polysaccharides: From Random Coil, to Single Helix to Supramolecular Assembly. Biomacromolecules 2019, 20, 1731–1739. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Shen, M.; Song, Q.; Xie, J. Biological Activities and Pharmaceutical Applications of Polysaccharide from Natural Resources: A Review. Carbohydr. Polym. 2018, 183, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Huang, G. Preparation and Immunological Activity of Polysaccharides and Their Derivatives. Int. J. Biol. Macromol. 2018, 112, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Naqash, F.; Masoodi, F.A.; Rather, S.A.; Wani, S.M.; Gani, A. Emerging Concepts in the Nutraceutical and Functional Properties of Pectin-A Review. Carbohydr. Polym. 2017, 168, 227–239. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Zhang, J.; Zhao, B.; Yao, J.; Wang, Y. Structure-Antioxidant Relationships of Sulfated Galactomannan from Guar Gum. Int. J. Biol. Macromol. 2010, 46, 59–66. [Google Scholar] [CrossRef]
- Cunha de Padua, M.M.; Suter Correia Cadena, S.M.; de Oliveira Petkowicz, C.L.; Martinez, G.R.; Rodrigues Noleto, G. Galactomannan from Schizolobium Amazonicum Seed and Its Sulfated Derivatives Impair Metabolism in HepG2 Cells. Int. J. Biol. Macromol. 2017, 101, 464–473. [Google Scholar] [CrossRef]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure-Function Relationships of Immunostimulatory Polysaccharides: A Review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Shen, M.; Hong, Y.; Ye, H.; Huang, L.; Xie, J. Chemical Modifications of Polysaccharides and Their Anti-Tumor Activities. Carbohydr. Polym. 2020, 229, 115436. [Google Scholar] [CrossRef]
- Li, J.; Shen, B.; Nie, S.; Duan, Z.; Chen, K. A Combination of Selenium and Polysaccharides: Promising Therapeutic Potential. Carbohydr. Polym. 2019, 206, 163–173. [Google Scholar] [CrossRef]
- Kaleta, B.; Górski, A.; Zagożdżon, R.; Cieślak, M.; Kaźmierczak-Barańska, J.; Nawrot, B.; Klimaszewska, M.; Malinowska, E.; Górska, S.; Turło, J. Selenium-Containing Polysaccharides from Lentinula Edodes—Biological Activity. Carbohydr. Polym. 2019, 223, 115078. [Google Scholar] [CrossRef]
- Lu, Z.; Jin, M.; Huang, M.; Wang, Y.; Wang, Y. Bioactivity of Selenium-Enriched Exopolysaccharides Produced by Enterobacter Cloacae Z0206 in Broilers. Carbohydr. Polym. 2013, 96, 131–136. [Google Scholar] [CrossRef]
- Malinowska, E.; Krzyczkowski, W.; Herold, F.; Lapienis, G.; Ślusarczyk, J.; Suchocki, P.; Kuraś, M.; Turlo, J. Biosynthesis of Selenium-Containing Polysaccharides with Antioxidant Activity in Liquid Culture of Hericium Erinaceum. Enzyme Microb. Technol. 2009, 44, 334–343. [Google Scholar] [CrossRef]
- Tapiero, H.; Townsend, D.M.; Tew, K.D. The Antioxidant Role of Selenium and Seleno-Compounds. Biomed. Pharmacother. Biomed. Pharmacother. 2003, 57, 134–144. [Google Scholar] [CrossRef]
- Roy, M.; Kiremidjian-Schumacher, L.; Wishe, H.I.; Cohen, M.W.; Stotzky, G. Supplementation with Selenium and Human Immune Cell Functions. I. Effect on Lymphocyte Proliferation and Interleukin 2 Receptor Expression. Biol. Trace Elem. Res. 1994, 41, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Dodig, S.; Cepelak, I. The Facts and Controversies about Selenium. Acta Pharm. 2004, 54, 261–276. [Google Scholar] [PubMed]
- Rayman, M.P. Selenium and Human Health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium Intake, Status, and Health: A Complex Relationship. Hormones 2020, 19, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Murdolo, G.; Bartolini, D.; Tortoioli, C.; Piroddi, M.; Torquato, P.; Galli, F. Selenium and Cancer Stem Cells. Adv. Cancer Res. 2017, 136, 235–257. [Google Scholar] [CrossRef]
- Fernandes, A.P.; Gandin, V. Selenium Compounds as Therapeutic Agents in Cancer. Biochim. Biophys. Acta 2015, 1850, 1642–1660. [Google Scholar] [CrossRef]
- Kim, C.; Kim, S.-B.; Park, M.-S. Synthesis of Novel 3-Allylseleno-6-Alkylthiopyridazines: Their Anticancer Activity against MCF-7 Cells. Arch. Pharm. Res. 2014, 37, 452–458. [Google Scholar] [CrossRef]
- Domínguez-Álvarez, E.; Gajdács, M.; Spengler, G.; Palop, J.A.; Marć, M.A.; Kieć-Kononowicz, K.; Amaral, L.; Molnár, J.; Jacob, C.; Handzlik, J.; et al. Identification of Selenocompounds with Promising Properties to Reverse Cancer Multidrug Resistance. Bioorg. Med. Chem. Lett. 2016, 26, 2821–2824. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.Y.; Zhu, Y.J.; Li, W.G.; Huang, Q.S.; Huang, C.Z.; Zhang, Q.N.; Hou, C. A Preliminary Report on the Intervention Trials of Primary Liver Cancer in High-Risk Populations with Nutritional Supplementation of Selenium in China. Biol. Trace Elem. Res. 1991, 29, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Hehr, T.; Hoffmann, W.; Bamberg, M. Role of sodium selenite as an adjuvant in radiotherapy of rectal carcinoma. Med. Klin. 1997, 92 (Suppl. S3), 48–49. [Google Scholar] [CrossRef] [PubMed]
- El-Bayoumy, K. The Protective Role of Selenium on Genetic Damage and on Cancer. Mutat. Res. 2001, 475, 123–139. [Google Scholar] [CrossRef]
- Hiffler, L.; Rakotoambinina, B. Selenium and RNA Virus Interactions: Potential Implications for SARS-CoV-2 Infection (COVID-19). Front. Nutr. 2020, 7, 164. [Google Scholar] [CrossRef]
- Chaturvedi, U.C.; Shrivastava, R.; Upreti, R.K. Viral Infections and Trace Elements: A Complex Interaction. Curr. Sci. 2004, 87, 1536–1554. [Google Scholar]
- Zong, A.; Cao, H.; Wang, F. Anticancer Polysaccharides from Natural Resources: A Review of Recent Research. Carbohydr. Polym. 2012, 90, 1395–1410. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Date, A.; Chawda, H.; Patel, K. Polysaccharides as Potential Anticancer Agents—A Review of Their Progress. Carbohydr. Polym. 2019, 210, 412–428. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, V.; Saravanan, S.; Al-Maleki, A.R.; Ramesh, M.; Vadivelu, J. A Review of Natural Polysaccharides for Drug Delivery Applications: Special Focus on Cellulose, Starch and Glycogen. Biomed. Pharmacother. 2018, 107, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Kulicke, W.-M.; Heinze, T. Improvements in Polysaccharides for Use as Blood Plasma Expanders. Macromol. Symp. 2005, 231, 47–59. [Google Scholar] [CrossRef]
- Sun, Y.; Jing, X.; Ma, X.; Feng, Y.; Hu, H. Versatile Types of Polysaccharide-Based Drug Delivery Systems: From Strategic Design to Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 9159. [Google Scholar] [CrossRef]
- Bohn, J.A.; BeMiller, J.N. (1→3)-β-d-Glucans as Biological Response Modifiers: A Review of Structure-Functional Activity Relationships. Carbohydr. Polym. 1995, 28, 3–14. [Google Scholar] [CrossRef]
- Okazaki, M.; Adachi, Y.; Ohno, N.; Yadomae, T. Structure-Activity Relationship of (1.RAR.3)-.BETA.-D-Glucans in the Induction of Cytokine Production from Macrophages, in Vitro. Biol. Pharm. Bull. 1995, 18, 1320–1327. [Google Scholar] [CrossRef] [Green Version]
- Surenjav, U.; Zhang, L.; Xu, X.; Zhang, X.; Zeng, F. Effects of Molecular Structure on Antitumor Activities of (1→3)-β-d-Glucans from Different Lentinus Edodes. Carbohydr. Polym. 2006, 63, 97–104. [Google Scholar] [CrossRef]
- Ohno, N.; Miura, N.N.; Chiba, N.; Adachi, Y.; Yadomae, T. Comparison of the Immunopharmacological Activities of Triple and Single-Helical Schizophyllan in Mice. Biol. Pharm. Bull. 1995, 18, 1242–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, S.; Wang, X.; Zhang, L.; Cheung, P.C.K. Advances in Lentinan: Isolation, Structure, Chain Conformation and Bioactivities. Food Hydrocoll. 2011, 25, 196–206. [Google Scholar] [CrossRef]
- Leung, M.Y.K.; Liu, C.; Koon, J.C.M.; Fung, K.P. Polysaccharide Biological Response Modifiers. Immunol. Lett. 2006, 105, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, E.; Klimaszewska, M.; Strączek, T.; Schneider, K.; Kapusta, C.; Podsadni, P.; Łapienis, G.; Dawidowski, M.; Kleps, J.; Górska, S.; et al. Selenized Polysaccharides—Biosynthesis and Structural Analysis. Carbohydr. Polym. 2018, 198, 407–417. [Google Scholar] [CrossRef]
- Lombard, Y.; Giaimis, J.; Makaya-Kumba, M.; Fonteneau, P.; Poindron, P. A New Method for Studying the Binding and Ingestion of Zymosan Particles by Macrophages. J. Immunol. Methods 1994, 174, 155–165. [Google Scholar] [CrossRef]
- Sakagami, H.; Aoki, T.; Simpson, A.; Tanuma, S. Induction of Immunopotentiation Activity by a Protein-Bound Polysaccharide, PSK (Review). Anticancer Res. 1991, 11, 993–999. [Google Scholar]
- Tsiapali, E.; Whaley, S.; Kalbfleisch, J.; Ensley, H.E.; Browder, I.W.; Williams, D.L. Glucans Exhibit Weak Antioxidant Activity, but Stimulate Macrophage Free Radical Activity. Free Radic. Biol. Med. 2001, 30, 393–402. [Google Scholar] [CrossRef]
- He, N.; Shi, X.; Zhao, Y.; Tian, L.; Wang, D.; Yang, X. Inhibitory Effects and Molecular Mechanisms of Selenium-Containing Tea Polysaccharides on Human Breast Cancer MCF-7 Cells. J. Agric. Food Chem. 2013, 61, 579–588. [Google Scholar] [CrossRef]
- Liu, Y.; Zeng, S.; Liu, Y.; Wu, W.; Shen, Y.; Zhang, L.; Li, C.; Chen, H.; Liu, A.; Shen, L.; et al. Synthesis and Antidiabetic Activity of Selenium Nanoparticles in the Presence of Polysaccharides from Catathelasma Ventricosum. Int. J. Biol. Macromol. 2018, 114, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, J.; Ding, D.; Zhang, L.; Muehlmann, L.A.; Deng, S.-E.; Wang, X.; Li, W.; Zhang, W. Synthesis and Antioxidant Properties of Lycium Barbarum Polysaccharides Capped Selenium Nanoparticles Using Tea Extract. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1463–1470. [Google Scholar] [CrossRef] [PubMed]
- Satitmanwiwat, S.; Ratanakhanokchai, K.; Laohakunjit, N.; Chao, L.K.; Chen, S.-T.; Pason, P.; Tachaapaikoon, C.; Kyu, K.L. Improved Purity and Immunostimulatory Activity of β-(1→3)(1→6)-Glucan from Pleurotus Sajor-Caju Using Cell Wall-Degrading Enzymes. J. Agric. Food Chem. 2012, 60, 5423–5430. [Google Scholar] [CrossRef] [PubMed]
- Ercole, C.; Cacchio, P.; Botta, A.L.; Centi, V.; Lepidi, A. Bacterially Induced Mineralization of Calcium Carbonate: The Role of Exopolysaccharides and Capsular Polysaccharides. Microsc. Microanal. Off. J. Microsc. Soc. Am. Microbeam Anal. Soc. Microsc. Soc. Can. 2007, 13, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yildiz, H.; Karatas, N. Microbial Exopolysaccharides: Resources and Bioactive Properties. Process Biochem. 2018, 72, 41–46. [Google Scholar] [CrossRef]
- Borgenström, M.; Wärri, A.; Hiilesvuo, K.; Käkönen, R.; Käkönen, S.; Nissinen, L.; Pihlavisto, M.; Marjamäki, A.; Vlodavsky, I.; Naggi, A.; et al. O-Sulfated Bacterial Polysaccharides with Low Anticoagulant Activity Inhibit Metastasis. Semin. Thromb. Hemost. 2007, 33, 547–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Luo, J.; Ye, H.; Zeng, X. Preparation, Antioxidant and Antitumor Activities in Vitro of Different Derivatives of Levan from Endophytic Bacterium Paenibacillus polymyxa EJS-3. Food Chem. Toxicol. 2012, 50, 767–772. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, Y.; Ren, S.; Han, Y.; Cheng, H. Studies on the Chemical Structure and Antitumor Activity of an Exopolysaccharide from Rhizobium Sp. N613. Carbohydr. Res. 2010, 345, 637–643. [Google Scholar] [CrossRef]
- Harada, T.; Fujimori, K.; Hirose, S.; Masada, M. Growth and β-Glucan 10C3K Production by a Mutant of Alcaligenes faecalis Var. Myxogenes in Defined Medium. Agric. Biol. Chem. 1966, 30, 764–769. [Google Scholar] [CrossRef]
- McIntosh, M.; Stone, B.A.; Stanisich, V.A. Curdlan and Other Bacterial (1-->3)-Beta-D-Glucans. Appl. Microbiol. Biotechnol. 2005, 68, 163–173. [Google Scholar] [CrossRef]
- Zhan, X.-B.; Lin, C.-C.; Zhang, H.-T. Recent Advances in Curdlan Biosynthesis, Biotechnological Production, and Applications. Appl. Microbiol. Biotechnol. 2012, 93, 525–531. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An Immunomodulatory Molecule of Symbiotic Bacteria Directs Maturation of the Host Immune System. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Huang, G. Preparation and Drug Delivery of Dextran-Drug Complex. Drug Deliv. 2019, 26, 252–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Philippis, R. Exocellular Polysaccharides from Cyanobacteria and Their Possible Applications. FEMS Microbiol. Rev. 1998, 22, 151–175. [Google Scholar] [CrossRef]
- Wang, B.; Liu, Q.; Huang, Y.; Yuan, Y.; Ma, Q.; Du, M.; Cai, T.; Cai, Y. Extraction of Polysaccharide from Spirulina and Evaluation of Its Activities. Evid. Based Complement. Alternat. Med. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rajasekar, P.; Palanisamy, S.; Anjali, R.; Vinosha, M.; Elakkiya, M.; Marudhupandi, T.; Tabarsa, M.; You, S.; Prabhu, N.M. Isolation and Structural Characterization of Sulfated Polysaccharide from Spirulina Platensis and Its Bioactive Potential: In Vitro Antioxidant, Antibacterial Activity and Zebrafish Growth and Reproductive Performance. Int. J. Biol. Macromol. 2019, 141, 809–821. [Google Scholar] [CrossRef]
- Usov, A.I.; Zelinsky, N.D. Chemical structures of algal polysaccharides. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Cambridge, UK, 2013; pp. 23–86. ISBN 978-0-85709-512-1. [Google Scholar]
- Aquino, R.S.; Grativol, C.; Mourão, P.A.S. Rising from the Sea: Correlations between Sulfated Polysaccharides and Salinity in Plants. PLoS ONE 2011, 6, e18862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Weelden, G.; Bobiński, M.; Okła, K.; van Weelden, W.J.; Romano, A.; Pijnenborg, J.M.A. Fucoidan Structure and Activity in Relation to Anti-Cancer Mechanisms. Mar. Drugs 2019, 17, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitton, J.H. Therapies from Fucoidan; Multifunctional Marine Polymers. Mar. Drugs 2011, 9, 1731–1760. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.; de Morais, A.; de Morais, R. Marine Polysaccharides from Algae with Potential Biomedical Applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef] [PubMed]
- Yoo, Y.-C.; Kim, W.-J.; Kim, S.-Y.; Kim, S.-M.; Chung, M.-K.; Park, J.-W.; Suh, H.-H.; Lee, K.-B.; Park, Y.-I. Immunomodulating Activity of a Fucoidan Isolated from Korean Undaria Pinnatifida Sporophyll. ALGAE 2007, 22, 333–338. [Google Scholar] [CrossRef]
- Byon, Y.Y.; Kim, M.H.; Yoo, E.S.; Hwang, K.K.; Jee, Y.; Shin, T.; Joo, H.G. Radioprotective Effects of Fucoidan on Bone Marrow Cells: Improvement of the Cell Survival and Immunoreactivity. J. Vet. Sci. 2008, 9, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Sun, J.; Su, X.; Yu, Q.; Yu, Q.; Zhang, P. A Review about the Development of Fucoidan in Antitumor Activity: Progress and Challenges. Carbohydr. Polym. 2016, 154, 96–111. [Google Scholar] [CrossRef]
- Lim, S.J.; Wan Aida, W.M. Extraction of Sulfated Polysaccharides (Fucoidan) from Brown Seaweed. In Seaweed Polysaccharides; Elsevier: Amsterdam, The Netherlands, 2017; pp. 27–46. ISBN 978-0-12-809816-5. [Google Scholar]
- Zhang, J.; Zhang, Q.; Wang, J.; Shi, X.; Zhang, Z. Analysis of the Monosaccharide Composition of Fucoidan by Precolumn Derivation HPLC. Chin. J. Oceanol. Limnol. 2009, 27, 578–582. [Google Scholar] [CrossRef]
- Malyarenko, O.S.; Usoltseva, R.V.; Silchenko, A.S.; Ermakova, S.P. Aminated Laminaran from Brown Alga Saccharina Cichorioides: Synthesis, Structure, Anticancer, and Radiosensitizing Potential in Vitro. Carbohydr. Polym. 2020, 250, 117007. [Google Scholar] [CrossRef] [PubMed]
- Gamal-Eldeen, A.M.; Ahmed, E.F.; Abo-Zeid, M.A. In Vitro Cancer Chemopreventive Properties of Polysaccharide Extract from the Brown Alga, Sargassum Latifolium. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2009, 47, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Lins, K.O.A.L.; Bezerra, D.P.; Alves, A.P.N.N.; Alencar, N.M.N.; Lima, M.W.; Torres, V.M.; Farias, W.R.L.; Pessoa, C.; de Moraes, M.O.; Costa-Lotufo, L.V. Antitumor Properties of a Sulfated Polysaccharide from the Red Seaweed Champia Feldmannii (Diaz-Pifferer). J. Appl. Toxicol. JAT 2009, 29, 20–26. [Google Scholar] [CrossRef]
- Zhou, G. In Vivo Antitumor and Immunomodulation Activities of Different Molecular Weight Lambda-Carrageenans from Chondrus Ocellatus. Pharmacol. Res. 2004, 50, 47–53. [Google Scholar] [CrossRef]
- Zhou, G.; Sheng, W.; Yao, W.; Wang, C. Effect of Low Molecular Lambda-Carrageenan from Chondrus Ocellatus on Antitumor H-22 Activity of 5-Fu. Pharmacol. Res. 2006, 53, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Necas, J.; Bartosikova, L. Carrageenan: A Review. Veterinární Medicína 2013, 58, 187–205. [Google Scholar] [CrossRef] [Green Version]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H. Chemical Structures and Bioactivities of Sulfated Polysaccharides from Marine Algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [Green Version]
- Martínez Andrade, K.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsen, B.S.; Olafsdóttir, E.S.; Ingólfsdóttir, K. Chromatography and Electrophoresis in Separation and Characterization of Polysaccharides from Lichens. J. Chromatogr. A 2002, 967, 163–171. [Google Scholar] [CrossRef]
- Olafsdottir, E.S.; Ingólfsdottir, K. Polysaccharides from Lichens: Structural Characteristics and Biological Activity. Planta Med. 2001, 67, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Willför, S.; Xu, C. A Review of Bioactive Plant Polysaccharides: Biological Activities, Functionalization, and Biomedical Applications. Bioact. Carbohydr. Diet. Fibre 2015, 5, 31–61. [Google Scholar] [CrossRef]
- Chang, S.-T.; Wasser, S.P. The Role of Culinary-Medicinal Mushrooms on Human Welfare with a Pyramid Model for Human Health. Int. J. Med. Mushrooms 2012, 14, 95–134. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Nishitani, Y. Immunomodulating Compounds in Basidiomycetes. J. Clin. Biochem. Nutr. 2013, 52, 202–207. [Google Scholar] [CrossRef] [Green Version]
- Wasser, S.P. Current Findings, Future Trends, and Unsolved Problems in Studies of Medicinal Mushrooms. Appl. Microbiol. Biotechnol. 2011, 89, 1323–1332. [Google Scholar] [CrossRef]
- Giavasis, I. Production of microbial polysaccharides for use in food. In Microbial Production of Food Ingredients, Enzymes and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2013; pp. 413–468. ISBN 978-0-85709-343-1. [Google Scholar]
- Friedman, M. Mushroom Polysaccharides: Chemistry and Antiobesity, Antidiabetes, Anticancer, and Antibiotic Properties in Cells, Rodents, and Humans. Foods 2016, 5, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moradali, M.-F.; Mostafavi, H.; Ghods, S.; Hedjaroude, G.-A. Immunomodulating and Anticancer Agents in the Realm of Macromycetes Fungi (Macrofungi). Int. Immunopharmacol. 2007, 7, 701–724. [Google Scholar] [CrossRef]
- Reshetnikov, S.V.; Tan, K.-K. Higher Basidiomycota as a Source of Antitumor and Immunostimulating Polysaccharides (Review). Int. J. Med. Mushrooms 2001, 3, 34. [Google Scholar] [CrossRef]
- Boon, H.; Wong, J. Botanical Medicine and Cancer: A Review of the Safety and Efficacy. Expert Opin. Pharmacother. 2004, 5, 2485–2501. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R.; Smith, J.E.; Rowan, N.J. Medicinal Mushrooms and Cancer Therapy: Translating a Traditional Practice into Western Medicine. Perspect. Biol. Med. 2006, 49, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Yeung, K.; Gubili, J. Shiitake Mushroom (Lentinula edodes). J. Soc. Integr. Oncol. 2008, 6, 134–135. [Google Scholar]
- Zhang, Y.; Kong, H.; Fang, Y.; Nishinari, K.; Phillips, G.O. Schizophyllan: A Review on Its Structure, Properties, Bioactivities and Recent Developments. Bioact. Carbohydr. Diet. Fibre 2013, 1, 53–71. [Google Scholar] [CrossRef]
- Kumari, M.; Survase, S.A.; Singhal, R.S. Production of Schizophyllan Using Schizophyllum Commune NRCM. Bioresour. Technol. 2008, 99, 1036–1043. [Google Scholar] [CrossRef]
- Giavasis, I. Bioactive Fungal Polysaccharides as Potential Functional Ingredients in Food and Nutraceuticals. Curr. Opin. Biotechnol. 2014, 26, 162–173. [Google Scholar] [CrossRef]
- Bao, X.-F.; Wang, X.-S.; Dong, Q.; Fang, J.-N.; Li, X.-Y. Structural Features of Immunologically Active Polysaccharides from Ganoderma Lucidum. Phytochemistry 2002, 59, 175–181. [Google Scholar] [CrossRef]
- Eo, S.-K.; Kim, Y.-S.; Lee, C.-K.; Han, S.-S. Possible Mode of Antiviral Activity of Acidic Protein Bound Polysaccharide Isolated from Ganoderma lucidum on Herpes Simplex Viruses. J. Ethnopharmacol. 2000, 72, 475–481. [Google Scholar] [CrossRef]
- Wasser, S.P. Medicinal Mushrooms as a Source of Antitumor and Immunomodulating Polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [CrossRef] [PubMed]
- Bergendiova, K.; Tibenska, E.; Majtan, J. Pleuran (β-Glucan from Pleurotus Ostreatus) Supplementation, Cellular Immune Response and Respiratory Tract Infections in Athletes. Eur. J. Appl. Physiol. 2011, 111, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Míčková, K.; Synytsya, A.; Jablonský, I.; Spěváček, J.; Erban, V.; Kováříková, E.; Čopíková, J. Glucans from Fruit Bodies of Cultivated Mushrooms Pleurotus Ostreatus and Pleurotus Eryngii: Structure and Potential Prebiotic Activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar] [CrossRef]
- Boh, B.; Berovic, M. Grifola Frondosa (Dicks.: Fr.) S.F. Gray (Maitake Mushroom): Medicinal Properties, Active Compounds, and Biotechnological Cultivation. Int. J. Med. Mushrooms 2007, 9, 89–108. [Google Scholar] [CrossRef]
- Ye, L.; Li, J.; Zhang, J.; Pan, Y. NMR Characterization for Polysaccharide Moiety of a Glycopeptide. Fitoterapia 2010, 81, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Biliaderis, C.G.; Izydorczyk, M.S. (Eds.) Functional Food Carbohydrates; Functional foods and nutraceuticals series; CRC Press: Boca Raton, FL, USA, 2007; ISBN 978-0-8493-1822-1. [Google Scholar]
- Mizuno, T. Medicinal Properties and Clinical Effects of Culinary-Medicinal Mushroom Agaricus Blazei Murrill (Agaricomycetideae) (Review). Int. J. Med. Mushrooms 2002, 4, 14. [Google Scholar] [CrossRef]
- Hobbs, C. Medicinal Value of Turkey Tail Fungus Trametes Versicolor (L.:Fr.) Pilat (Aphyllophoromycetideae). A Literature Review. Int. J. Med. Mushrooms 2004, 6, 195–218. [Google Scholar] [CrossRef] [Green Version]
- Ooi, V.E.; Liu, F. Immunomodulation and Anti-Cancer Activity of Polysaccharide-Protein Complexes. Curr. Med. Chem. 2000, 7, 715–729. [Google Scholar] [CrossRef] [Green Version]
- De Baets, S.; Vandamme, E.J. Extracellular Tremella Polysaccharedes: Structure, properties and applications. Biotechnol. Lett. 2001, 23, 1361–1366. [Google Scholar] [CrossRef]
- Khondkar, P. Composition and Partial Structure Characterization of Tremella Polysaccharides. Mycobiology 2009, 37, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Lo, H.-C.; Tsai, F.-A.; Wasser, S.P.; Yang, J.-G.; Huang, B.-M. Effects of Ingested Fruiting Bodies, Submerged Culture Biomass, and Acidic Polysaccharide Glucuronoxylomannan of Tremella Mesenterica Retz.:Fr. on Glycemic Responses in Normal and Diabetic Rats. Life Sci. 2006, 78, 1957–1966. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, H. Production of Intracellular Selenium-Enriched Polysaccharides from Thin Stillage by Cordyceps Sinensis and Its Bioactivities. Food Nutr. Res. 2016, 60, 30153. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Peng, T. Traditional Chinese Herbal Medicine as a Source of Molecules with Antiviral Activity. Antiviral Res. 2013, 97, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Quinn, M.T. Botanical Polysaccharides: Macrophage Immunomodulation and Therapeutic Potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.-Q.; Zheng, Z.-J.; Peng, Y.; Li, W.-X.; Chen, X.-M.; Lu, J.-X. Opposite Effects on Tumor Growth Depending on Dose of Achyranthes Bidentata Polysaccharides in C57BL/6 Mice. Int. Immunopharmacol. 2007, 7, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wu, D.; Ning, X.; Yang, G.; Lin, Z.; Tian, M.; Zhou, Y. α-Amylase-Assisted Extraction of Polysaccharides from Panax Ginseng. Int. J. Biol. Macromol. 2015, 75, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cai, J.; Geng, J.; Li, Y.; Wang, Z.; Li, R. Purification, Characterization and Anticancer Activity of a Polysaccharide from Panax Ginseng. Int. J. Biol. Macromol. 2012, 51, 968–973. [Google Scholar] [CrossRef]
- Cheng, H.; Li, S.; Fan, Y.; Gao, X.; Hao, M.; Wang, J.; Zhang, X.; Tai, G.; Zhou, Y. Comparative Studies of the Antiproliferative Effects of Ginseng Polysaccharides on HT-29 Human Colon Cancer Cells. Med. Oncol. 2011, 28, 175–181. [Google Scholar] [CrossRef]
- Tomoda, M.; Shimada, K.; Konno, C.; Sugiyama, K.; Hikino, H. Partial Structure of Panaxan A, a Hypoglycaemic Glycan of Panax ginseng Roots. Planta Med. 1984, 50, 436–438. [Google Scholar] [CrossRef]
- Sun, L.; Peng, X.; Sun, P.; Shi, J.; Yuan, X.; Zhu, J.; Tai, G.; Zhou, Y. Structural Characterization and Immunostimulatory Activity of a Novel Linear α-(1 → 6)-D-Glucan Isolated from Panax Ginseng C. A. Meyer. Glycoconj. J. 2012, 29, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, C.; Gafner, S. Quantitative Analysis of the Polysaccharide and Glycoprotein Fractions in Echinacea purpurea. and Echinacea angustifolia. by HPLC-ELSD for Quality Control of Raw Material. Pharm. Biol. 2007, 45, 98–105. [Google Scholar] [CrossRef]
- Jain, V.K. Chapter 1. An Overview of Organoselenium Chemistry: From Fundamentals to Synthesis. In Organoselenium Compounds in Biology and Medicine; Jain, V.K., Priyadarsini, K.I., Eds.; Royal Society of Chemistry: Cambridge, UK, 2017; pp. 1–33. ISBN 978-1-78801-029-0. [Google Scholar]
- Soriano-Garcia, M. Organoselenium Compounds as Potential Therapeutic and Chemopreventive Agents: A Review. Curr. Med. Chem. 2004, 11, 1657–1669. [Google Scholar] [CrossRef]
- Takahashi, K.; Suzuki, N.; Ogra, Y. Bioavailability Comparison of Nine Bioselenocompounds In Vitro and In Vivo. Int. J. Mol. Sci. 2017, 18, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrauzer, G.N.; Surai, P.F. Selenium in Human and Animal Nutrition: Resolved and Unresolved Issues. A Partly Historical Treatise in Commemoration of the Fiftieth Anniversary of the Discovery of the Biological Essentiality of Selenium, Dedicated to the Memory of Klaus Schwarz (1914–1978) on the Occasion of the Thirtieth Anniversary of His Death. Crit. Rev. Biotechnol. 2009, 29, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. The Importance of Selenium to Human Health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [Green Version]
- O’Dell, B.L. (Ed.) Handbook of Nutritionally Essential Mineral Elements; Clinical Nutrition in Health and Disease; Dekker: New York, NY, USA, 1997; ISBN 978-0-8247-9312-8. [Google Scholar]
- Roussel, A.-M.; Anderson, R.A.; Favier, A. International Symposium on Trace Elements in Man and Animals. In Trace Elements in Man and Animals 10; Kluwer Academic Publishers: New York, NY, USA, 2006; ISBN 978-0-306-47466-8. [Google Scholar]
- Schomburg, L.; Köhrle, J. On the Importance of Selenium and Iodine Metabolism for Thyroid Hormone Biosynthesis and Human Health. Mol. Nutr. Food Res. 2008, 52, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G.P. Selenium-Dependent Antioxidant Enzymes: Actions and Properties of Selenoproteins. Antioxidants 2018, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Speckmann, B.; Grune, T. Epigenetic Effects of Selenium and Their Implications for Health. Epigenetics 2015, 10, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Broome, C.S.; McArdle, F.; Kyle, J.A.M.; Andrews, F.; Lowe, N.M.; Hart, C.A.; Arthur, J.R.; Jackson, M.J. An Increase in Selenium Intake Improves Immune Function and Poliovirus Handling in Adults with Marginal Selenium Status. Am. J. Clin. Nutr. 2004, 80, 154–162. [Google Scholar] [CrossRef]
- Kiremidjian-Schumacher, L.; Roy, M.; Wishe, H.I.; Cohen, M.W.; Stotzky, G. Supplementation with Selenium and Human Immune Cell Functions. II. Effect on Cytotoxic Lymphocytes and Natural Killer Cells. Biol. Trace Elem. Res. 1994, 41, 115–127. [Google Scholar] [CrossRef]
- Hoffmann, P.R. Mechanisms by Which Selenium Influences Immune Responses. Arch. Immunol. Ther. Exp. 2007, 55, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The Role of Selenium in Inflammation and Immunity: From Molecular Mechanisms to Therapeutic Opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [Green Version]
- Vinceti, M.; Filippini, T.; Del Giovane, C.; Dennert, G.; Zwahlen, M.; Brinkman, M.; Zeegers, M.P.; Horneber, M.; D’Amico, R.; Crespi, C.M. Selenium for Preventing Cancer. Cochrane Database Syst. Rev. 2018, 1, CD005195. [Google Scholar] [CrossRef] [PubMed]
- Evans, S.O.; Khairuddin, P.F.; Jameson, M.B. Optimising Selenium for Modulation of Cancer Treatments. Anticancer Res. 2017, 37, 6497–6509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, M.A. Selenium and Host Defence towards Viruses. Proc. Nutr. Soc. 1999, 58, 707–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillin, O.M.; Vindry, C.; Ohlmann, T.; Chavatte, L. Selenium, Selenoproteins and Viral Infection. Nutrients 2019, 11, 2101. [Google Scholar] [CrossRef] [Green Version]
- Steinbrenner, H.; Al-Quraishy, S.; Dkhil, M.A.; Wunderlich, F.; Sies, H. Dietary Selenium in Adjuvant Therapy of Viral and Bacterial Infections. Adv. Nutr. 2015, 6, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harthill, M. Review: Micronutrient Selenium Deficiency Influences Evolution of Some Viral Infectious Diseases. Biol. Trace Elem. Res. 2011, 143, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Louria, D.B. Undernutrition Can Affect the Invading Microorganism. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2007, 45, 470–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parr, R.M. Trace Elements in Human Nutrition and Health; Weltgesundheitsorganisation, FAO, Internationale Atomenergie-Organisation, Eds.; FAO: Geneva, Switzerland, 1996; ISBN 978-92-4-156173-0. [Google Scholar]
- Micronutrients for Older Adults. 2014. Available online: https://Lpi.Oregonstate.Edu/Mic/Life-Stages/Older-Adults (accessed on 1 January 2021).
- Goldhaber, S.B. Trace Element Risk Assessment: Essentiality vs. Toxicity. Regul. Toxicol. Pharmacol. RTP 2003, 38, 232–242. [Google Scholar] [CrossRef]
- Pedrero, Z.; Madrid, Y. Novel Approaches for Selenium Speciation in Foodstuffs and Biological Specimens: A Review. Anal. Chim. Acta 2009, 634, 135–152. [Google Scholar] [CrossRef]
- Institute of Medicine (U.S.) (Ed.) Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids: A Report of the Panel on Dietary Antioxidants and Related Compounds, Subcommittees on Upper Reference Levels of Nutrients and of Interpretation and Use of Dietary Reference Intakes, and the Standing Committee on the Scientific Evaluation of Dietary Reference Intakes, Food and Nutrition Board, Institute of Medicine; National Academy Press: Washington, DC, USA, 2000; ISBN 978-0-309-06949-6. [Google Scholar]
- Amoako, P.O.; Uden, P.C.; Tyson, J.F. Speciation of Selenium Dietary Supplements; Formation of S-(Methylseleno)Cysteine and Other Selenium Compounds. Anal. Chim. Acta 2009, 652, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Meltzer, H.M.; Norheim, G.; Bibow, K.; Myhre, K.; Holm, H. The Form of Selenium Determines the Response to Supplementation in a Selenium Replete Population. Eur. J. Clin. Nutr. 1990, 44, 435–446. [Google Scholar] [PubMed]
- Thiry, C.; Ruttens, A.; Temmerman, L.; Schneider, Y.-J.; Pussemier, L. Current knowledge in species-related bioavailability of selenium in food. Food Chem. 2012, 130, 767–784. [Google Scholar] [CrossRef]
- SES-Clinical: Selenium, Serum. Available online: mayocliniclabs.com (accessed on 1 January 2021).
- Rayman, M.P.; Stranges, S. Epidemiology of Selenium and Type 2 Diabetes: Can We Make Sense of It? Free Radic. Biol. Med. 2013, 65, 1557–1564. [Google Scholar] [CrossRef]
- Bleys, J.; Navas-Acien, A.; Stranges, S.; Menke, A.; Miller, E.R.; Guallar, E. Serum Selenium and Serum Lipids in US Adults. Am. J. Clin. Nutr. 2008, 88, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Duffield-Lillico, A.J.; Reid, M.E.; Turnbull, B.W.; Combs, G.F.; Slate, E.H.; Fischbach, L.A.; Marshall, J.R.; Clark, L.C. Baseline Characteristics and the Effect of Selenium Supplementation on Cancer Incidence in a Randomized Clinical Trial: A Summary Report of the Nutritional Prevention of Cancer Trial. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2002, 11, 630–639. [Google Scholar]
- Johnson, C.C.; Fordyce, F.M.; Rayman, M.P. Symposium on “Geographical and Geological Influences on Nutrition”: Factors Controlling the Distribution of Selenium in the Environment and Their Impact on Health and Nutrition. Proc. Nutr. Soc. 2010, 69, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.M.; Mo, X.Y.; Zou, X.Z.; Song, R.X.; Sun, W.Y.; Lu, W.; Chen, Q.; Yu, Y.X.; Zang, W.J. Association Study between Polymorphisms in Selenoprotein Genes and Susceptibility to Kashin-Beck Disease. Osteoarthritis Cartilage 2010, 18, 817–824. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.A.; Handy, J.; Levander, O.A. Host Nutritional Status: The Neglected Virulence Factor. Trends Microbiol. 2004, 12, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Rayman, M.P.; Lv, H.; Schomburg, L.; Cui, B.; Gao, C.; Chen, P.; Zhuang, G.; Zhang, Z.; Peng, X.; et al. Low Population Selenium Status Is Associated With Increased Prevalence of Thyroid Disease. J. Clin. Endocrinol. Metab. 2015, 100, 4037–4047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbaraly, T.N.; Akbaraly, N.T.; Hininger-Favier, I.; Carrière, I.; Arnaud, J.; Gourlet, V.; Roussel, A.-M.; Berr, C. Plasma Selenium over Time and Cognitive Decline in the Elderly. Epidemiol. Camb. Mass 2007, 18, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, D.J.; Fedirko, V.; Jenab, M.; Schomburg, L.; Méplan, C.; Freisling, H.; Bueno-de-Mesquita, H.B.a.; Hybsier, S.; Becker, N.-P.; Czuban, M.; et al. Selenium Status Is Associated with Colorectal Cancer Risk in the European Prospective Investigation of Cancer and Nutrition Cohort. Int. J. Cancer 2015, 136, 1149–1161. [Google Scholar] [CrossRef]
- Morris, J.S.; Crane, S.B. Selenium Toxicity from a Misformulated Dietary Supplement, Adverse Health Effects, and the Temporal Response in the Nail Biologic Monitor. Nutrients 2013, 5, 1024–1057. [Google Scholar] [CrossRef] [Green Version]
- MacFarquhar, J.K.; Broussard, D.L.; Melstrom, P.; Hutchinson, R.; Wolkin, A.; Martin, C.; Burk, R.F.; Dunn, J.R.; Green, A.L.; Hammond, R.; et al. Acute Selenium Toxicity Associated with a Dietary Supplement. Arch. Intern. Med. 2010, 170, 256–261. [Google Scholar] [CrossRef] [Green Version]
- Klayman, D.L.; Günther, W.H.H. Organic Selenium Compounds: Their Chemistry and Biology; The Chemistry of Organometallic Compounds; Wiley-Interscience: New York, NY, USA, 1973; ISBN 978-0-471-49032-6. [Google Scholar]
- Lenardão, E.J.; Santi, C.; Sancineto, L. New Frontiers in Organoselenium Compounds; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-92404-5. [Google Scholar]
- Hansen, D.; Duda, P.J.; Zayed, A.; Terry, N. Selenium Removal by Constructed Wetlands: Role of Biological Volatilization. Environ. Sci. Technol. 1998, 32, 591–597. [Google Scholar] [CrossRef] [Green Version]
- Frankenberger, W.T., Jr. Selenium in the Environment; CRC Press: Boca Raton, FL, USA, 1994; ISBN 978-0-824-478993-0. [Google Scholar]
- Shanker, A.K. Countering UV-B Stress in Plants: Does Selenium Have a Role? Plant Soil 2006, 282, 21–26. [Google Scholar] [CrossRef]
- White, P.J. Selenium Accumulation by Plants. Ann. Bot. 2016, 117, 217–235. [Google Scholar] [CrossRef] [Green Version]
- Mattmiller, S.A.; Carlson, B.A.; Sordillo, L.M. Regulation of Inflammation by Selenium and Selenoproteins: Impact on Eicosanoid Biosynthesis. J. Nutr. Sci. 2013, 2, e28. [Google Scholar] [CrossRef] [Green Version]
- Ogra, Y.; Ishiwata, K.; Takayama, H.; Aimi, N.; Suzuki, K.T. Identification of a Novel Selenium Metabolite, Se-Methyl-N-Acetylselenohexosamine, in Rat Urine by High-Performance Liquid Chromatography--Inductively Coupled Plasma Mass Spectrometry and--Electrospray Ionization Tandem Mass Spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life. Sci. 2002, 767, 301–312. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Ogra, Y.; Ishiwata, K.; Takayama, H.; Aimi, N.; Suzuki, K.T. Selenosugars Are Key and Urinary Metabolites for Selenium Excretion within the Required to Low-Toxic Range. Proc. Natl. Acad. Sci. USA 2002, 99, 15932–15936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuehnelt, D.; Kienzl, N.; Traar, P.; Le, N.H.; Francesconi, K.A.; Ochi, T. Selenium Metabolites in Human Urine after Ingestion of Selenite, L-Selenomethionine, or DL-Selenomethionine: A Quantitative Case Study by HPLC/ICPMS. Anal. Bioanal. Chem. 2005, 383, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Liu, Y.; Chen, X.; Wei, X. Extraction, Characterization and Antioxidant Activities of Se-Enriched Tea Polysaccharides. Int. J. Biol. Macromol. 2015, 77, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Turlo, J.; Gutkowska, B.; Herold, F.; Dawidowski, M.; Slowiński, T.; Zobel, A. Relationship between Selenium Accumulation and Mycelial Cell Composition in Lentinula Edodes (Berk.) Cultures. J. Toxicol. Environ. Health A 2010, 73, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.W.; Stone, F.G.A.; Wilkinson, G. Comprehensive Organometallic Chemistry II: A Review of the Literature 1982–1994; Pergamon: Oxford, UK; New York, NY, USA, 1995; ISBN 978-0-08-040608-4. [Google Scholar]
- Iwaoka, M. Nucleophilic Selenium. In Organoselenium Chemistry; Wirth, T., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2011; pp. 53–109. ISBN 978-3-527-64194-9. [Google Scholar]
- Back, T.G. Selenium: Organoselenium Chemistry. In Encyclopedia of Inorganic Chemistry; King, R.B., Crabtree, R.H., Lukehart, C.M., Atwood, D.A., Scott, R.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2006; p. ia214. ISBN 978-0-470-86078-6. [Google Scholar]
- Bhasin, K.K.; Arora, E.; Mehta, S.K.; Klapoetke, T.M. Preparation and Characterization of Symmetrical Bis[4-Chloro-2-Pyrimidyl] Dichalcogenide (S, Se, Te) and Unsymmetrical 4-Chloro-2-(Arylchalcogenyl) Pyrimidine: X-Ray Crystal Structure of 4-Chloro-2-(Phenylselanyl) Pyrimidine and 2-(p-Tolylselanyl)-4-Chloropyrimidine. J. Organomet. Chem. 2011, 696, 835–840. [Google Scholar] [CrossRef]
- Hodage, A.S.; Phadnis, P.P.; Wadawale, A.; Priyadarsini, K.I.; Jain, V.K. Synthesis, Characterization and Structures of 2-(3,5-Dimethylpyrazol-1-Yl)Ethylseleno Derivatives and Their Probable Glutathione Peroxidase (GPx) like Activity. Org. Biomol. Chem. 2011, 9, 2992–2998. [Google Scholar] [CrossRef]
- Zade, S.S.; Singh, H.B. Synthesis of Organoselenium Compounds. In PATAI’S Chemistry of Functional Groups; Rappoport, Z., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2012; p. pat0706. ISBN 978-0-470-68253-1. [Google Scholar]
- Panda, A. Chemistry of Selena Macrocycles. Coord. Chem. Rev. 2009, 253, 1056–1098. [Google Scholar] [CrossRef]
- Milton, M.D.; Khan, S.; Singh, J.D.; Mishra, V.; Khandelwal, B.L. A Facile Access to Chalcogen and Dichalcogen Bearing Dialkylamines and Diols. Tetrahedron Lett. 2005, 46, 755–758. [Google Scholar] [CrossRef]
- Krafft, F.; Vorster, W. Ueber Umwandlung des Diphenylsulfons in Diphenylsulfid und Diphenylselenid. Berichte Dtsch. Chem. Ges. 1893, 26, 2813–2822. [Google Scholar] [CrossRef] [Green Version]
- Ariyoshi, K.; Aso, Y.; Otsubo, T.; Ogura, F. Application of bis(p-methoxyphenyl)selenoxide as an oxidizing agent of kornblum oxidation. Chem. Lett. 1984, 13, 891–892. [Google Scholar] [CrossRef] [Green Version]
- Kelly, A.M.; Rosini, G.P.; Goldman, A.S. Oxygen Transfer from Organoelement Oxides to Carbon Monoxide Catalyzed by Transition Metal Carbonyls. J. Am. Chem. Soc. 1997, 119, 6115–6125. [Google Scholar] [CrossRef]
- Wessjohann, L.A.; Schneider, A.; Abbas, M.; Brandt, W. Selenium in Chemistry and Biochemistry in Comparison to Sulfur. Biol. Chem. 2007, 388, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Helt, J.-C.P.; Autschbach, J.; Detty, M.R. A New Reaction for Organoselenium Compounds: Alkyl Transfer from Diorganoselenium(IV) Dibromides to Alkenoic Acids To Give γ- and δ-Lactones. Organometallics 2009, 28, 3426–3436. [Google Scholar] [CrossRef]
- Reich, H.J.; Jasperse, C.P. Organoselenium Chemistry. Preparation and Reactions of 2,4,6-Tri-Tert-Butylbenzeneselenenic Acid. J. Org. Chem. 1988, 53, 2389–2390. [Google Scholar] [CrossRef]
- Tanaka, T.; Kohno, H.; Murakami, M.; Kagami, S.; El-Bayoumy, K. Suppressing Effects of Dietary Supplementation of the Organoselenium 1,4-Phenylenebis(Methylene)Selenocyanate and the Citrus Antioxidant Auraptene on Lung Metastasis of Melanoma Cells in Mice. Cancer Res. 2000, 60, 3713–3716. [Google Scholar] [PubMed]
- Rao, C.V.; Cooma, I.; Rodriguez, J.G.; Simi, B.; El-Bayoumy, K.; Reddy, B.S. Chemoprevention of Familial Adenomatous Polyposis Development in the APC(Min) Mouse Model by 1,4-Phenylene Bis(Methylene)Selenocyanate. Carcinogenesis 2000, 21, 617–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, C.V.; Wang, C.Q.; Simi, B.; Rodriguez, J.G.; Cooma, I.; El-Bayoumy, K.; Reddy, B.S. Chemoprevention of Colon Cancer by a Glutathione Conjugate of 1,4-Phenylenebis(Methylene)Selenocyanate, a Novel Organoselenium Compound with Low Toxicity. Cancer Res. 2001, 61, 3647–3652. [Google Scholar]
- Koketsu, M.; Yang, H.O.; Kim, Y.M.; Ichihashi, M.; Ishihara, H. Preparation of 1,4-Oxaselenin from AgNO(3)/LDA-Assisted Reaction of 3-Selena-4-Pentyn-1-One as Potential Antitumor Agents. Org. Lett. 2001, 3, 1705–1707. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Jiang, C.; Kaeck, M.; Ganther, H.; Ip, C.; Thompson, H. Cellular and Metabolic Effects of Triphenylselenonium Chloride in a Mammary Cell Culture Model. Carcinogenesis 1995, 16, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz, E.H.G.; Silvers, M.A.; Jardim, G.A.M.; Resende, J.M.; Cavalcanti, B.C.; Bomfim, I.S.; Pessoa, C.; de Simone, C.A.; Botteselle, G.V.; Braga, A.L.; et al. Synthesis and Antitumor Activity of Selenium-Containing Quinone-Based Triazoles Possessing Two Redox Centres, and Their Mechanistic Insights. Eur. J. Med. Chem. 2016, 122, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Hatfield, D.L. (Ed.) Selenium: Its Molecular Biology and Role in Human Health, 2nd ed.; Springer: New York, NY, USA, 2006; ISBN 978-0-387-33826-2. [Google Scholar]
- Roussyn, I.; Briviba, K.; Masumoto, H.; Sies, H. Selenium-Containing Compounds Protect DNA from Single-Strand Breaks Caused by Peroxynitrite. Arch. Biochem. Biophys. 1996, 330, 216–218. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.S.; Beckman, T.W.; Chen, J.; Marshall, P.A.; Freeman, B.A. Apparent Hydroxyl Radical Production by Peroxynitrite: Implications for Endothelial Injury from Nitric Oxide and Superoxide. Proc. Natl. Acad. Sci. USA 1990, 87, 1620–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.; Gerson, B.; Subramaniam, S. The Role of Copper, Molybdenum, Selenium, and Zinc in Nutrition and Health. Clin. Lab. Med. 1998, 18, 673–685. [Google Scholar] [CrossRef]
- Alpegiani, M.; Bedeschi, A.; Perrone, E.; Franceschi, G. 2-Selenacephems and 1-Dethia-1-Selenapenems. Tetrahedron Lett. 1986, 27, 3041–3044. [Google Scholar] [CrossRef]
- Stadtman, T.C. Selenocysteine. Annu. Rev. Biochem. 1996, 65, 83–100. [Google Scholar] [CrossRef]
- Nozawa, R.; Yokota, T.; Fujimoto, T. Susceptibility of Methicillin-Resistant Staphylococcus Aureus to the Selenium-Containing Compound 2-Phenyl-1,2-Benzoisoselenazol-3(2H)-One (PZ51). Antimicrob. Agents Chemother. 1989, 33, 1388–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Vlamis-Gardikas, A.; Kandasamy, K.; Zhao, R.; Gustafsson, T.N.; Engstrand, L.; Hoffner, S.; Engman, L.; Holmgren, A. Inhibition of Bacterial Thioredoxin Reductase: An Antibiotic Mechanism Targeting Bacteria Lacking Glutathione. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Braga, H.C.; Stefani, H.A.; Paixão, M.W.; Santos, F.W.; Lüdtke, D.S. Synthesis of 5′-Seleno-Xylofuranosides. Tetrahedron 2010, 66, 3441–3446. [Google Scholar] [CrossRef]
- Shakibaie, M.; Forootanfar, H.; Golkari, Y.; Mohammadi-Khorsand, T.; Shakibaie, M.R. Anti-Biofilm Activity of Biogenic Selenium Nanoparticles and Selenium Dioxide against Clinical Isolates of Staphylococcus Aureus, Pseudomonas Aeruginosa, and Proteus Mirabilis. J. Trace Elem. Med. Biol. 2015, 29, 235–241. [Google Scholar] [CrossRef]
- Alberto, E.E.; Rossato, L.L.; Alves, S.H.; Alves, D.; Braga, A.L. Imidazolium Ionic Liquids Containing Selenium: Synthesis and Antimicrobial Activity. Org. Biomol. Chem. 2011, 9, 1001–1003. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. AIDS Trials Ethics Questioned. Science 1997, 276, 520–523. [Google Scholar] [CrossRef]
- Du, J.; Surzhykov, S.; Lin, J.S.; Newton, M.G.; Cheng, Y.C.; Schinazi, R.F.; Chu, C.K. Synthesis, Anti-Human Immunodeficiency Virus and Anti-Hepatitis B Virus Activities of Novel Oxaselenolane Nucleosides. J. Med. Chem. 1997, 40, 2991–2993. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.K.; Ma, L.; Olgen, S.; Pierra, C.; Du, J.; Gumina, G.; Gullen, E.; Cheng, Y.C.; Schinazi, R.F. Synthesis and Antiviral Activity of Oxaselenolane Nucleosides. J. Med. Chem. 2000, 43, 3906–3912. [Google Scholar] [CrossRef] [PubMed]
- Sahu, P.K.; Kim, G.; Yu, J.; Ahn, J.Y.; Song, J.; Choi, Y.; Jin, X.; Kim, J.-H.; Lee, S.K.; Park, S.; et al. Stereoselective Synthesis of 4′-Selenonucleosides via Seleno-Michael Reaction as Potent Antiviral Agents. Org. Lett. 2014, 16, 5796–5799. [Google Scholar] [CrossRef]
- Sartori, G.; Jardim, N.S.; Marcondes Sari, M.H.; Dobrachinski, F.; Pesarico, A.P.; Rodrigues, L.C.; Cargnelutti, J.; Flores, E.F.; Prigol, M.; Nogueira, C.W. Antiviral Action of Diphenyl Diselenide on Herpes Simplex Virus 2 Infection in Female BALB/c Mice. J. Cell. Biochem. 2016, 117, 1638–1648. [Google Scholar] [CrossRef]
- Sahu, P.K.; Umme, T.; Yu, J.; Nayak, A.; Kim, G.; Noh, M.; Lee, J.-Y.; Kim, D.-D.; Jeong, L.S. Selenoacyclovir and Selenoganciclovir: Discovery of a New Template for Antiviral Agents. J. Med. Chem. 2015, 58, 8734–8738. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.-Y.; Xia, X.-M.; Li, P.; Wang, K.-Y. Inhibitory Effect of Sulfated Lentinan and Lentinan against Tobacco Mosaic Virus (TMV) in Tobacco Seedlings. Int. J. Biol. Macromol. 2013, 61, 264–269. [Google Scholar] [CrossRef]
- Mugesh, G.; du Mont, W.W.; Sies, H. Chemistry of Biologically Important Synthetic Organoselenium Compounds. Chem. Rev. 2001, 101, 2125–2179. [Google Scholar] [CrossRef]
- Duntas, L.H.; Benvenga, S. Selenium: An Element for Life. Endocrine 2015, 48, 756–775. [Google Scholar] [CrossRef]
- Jablonska, E.; Vinceti, M. Selenium and Human Health: Witnessing a Copernican Revolution? J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2015, 33, 328–368. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular Pathways and Physiological Roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantinescu-Aruxandei, D.; Frîncu, R.M.; Capră, L.; Oancea, F. Selenium Analysis and Speciation in Dietary Supplements Based on Next-Generation Selenium Ingredients. Nutrients 2018, 10, 1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocourt, C.R.B.; Cheng, W.-H. Selenium Supranutrition: Are the Potential Benefits of Chemoprevention Outweighed by the Promotion of Diabetes and Insulin Resistance? Nutrients 2013, 5, 1349–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, J.; Hu, X.; Ryan Smith, M.; Go, Y.-M.; Jones, D.P. Selenium at the Redox Interface of the Genome, Metabolome and Exposome. Free Radic. Biol. Med. 2018, 127, 215–227. [Google Scholar] [CrossRef]
- Kubachka, K.M.; Hanley, T.; Mantha, M.; Wilson, R.A.; Falconer, T.M.; Kassa, Z.; Oliveira, A.; Landero, J.; Caruso, J. Evaluation of Selenium in Dietary Supplements Using Elemental Speciation. Food Chem. 2017, 218, 313–320. [Google Scholar] [CrossRef]
- Gosetti, F.; Frascarolo, P.; Polati, S.; Medana, C.; Gianotti, V.; Palma, P.; Aigotti, R.; Baiocchi, C.; Gennaro, M.C. Speciation of Selenium in Diet Supplements by HPLC–MS/MS Methods. Food Chem. 2007, 105, 1738–1747. [Google Scholar] [CrossRef]
- Helzlsouer k, M.S. Acute Selenium Intoxication in the United States. Fed. Proc. 1985, 44, 1670. [Google Scholar]
- Clark, R.F.; Strukle, E.; Williams, S.R.; Manoguerra, A.S. Selenium Poisoning from a Nutritional Supplement. JAMA 1996, 275, 1087–1088. [Google Scholar] [CrossRef]
- Skalickova, S.; Milosavljevic, V.; Cihalova, K.; Horky, P.; Richtera, L.; Adam, V. Selenium Nanoparticles as a Nutritional Supplement. Nutrition 2017, 33, 83–90. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, C.; Tian, C.; Ren, Z.; Song, X.; Wang, X.; Xu, N.; Jing, H.; Li, S.; Liu, W.; et al. Characterization, Antioxidation, Anti-Inflammation and Renoprotection Effects of Selenized Mycelia Polysaccharides from Oudemansiella Radicata. Carbohydr. Polym. 2018, 181, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Jing, H.; Zhang, J.; Che, G.; Zhou, M.; Gao, Z.; Li, S.; Ren, Z.; Hao, L.; Liu, Y.; et al. Optimization of Mycelia Selenium Polysaccharide Extraction from Agrocybe Cylindracea SL-02 and Assessment of Their Antioxidant and Anti-Ageing Activities. PLoS ONE 2016, 11, e0160799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangiavacchi, F.; Coelho Dias, I.F.; Di Lorenzo, I.; Grzes, P.; Palomba, M.; Rosati, O.; Bagnoli, L.; Marini, F.; Santi, C.; Lenardao, E.J.; et al. Sweet Selenium: Synthesis and Properties of Selenium-Containing Sugars and Derivatives. Pharmaceuticals 2020, 13, 211. [Google Scholar] [CrossRef] [PubMed]
- Storkey, C.; Davies, M.J.; White, J.M.; Schiesser, C.H. Synthesis and Antioxidant Capacity of 5-Selenopyranose Derivatives. Chem. Commun. Camb. Engl. 2011, 47, 9693–9695. [Google Scholar] [CrossRef] [PubMed]
- Storkey, C.; Pattison, D.I.; White, J.M.; Schiesser, C.H.; Davies, M.J. Preventing Protein Oxidation with Sugars: Scavenging of Hypohalous Acids by 5-Selenopyranose and 4-Selenofuranose Derivatives. Chem. Res. Toxicol. 2012, 25, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Lucas, M.A.; Nguyen, O.T.K.; Schiesser, C.H.; Zheng, S.-L. Preparation of 5-Selenopentopyranose Sugars from Pentose Starting Materials by Samarium(II) Iodide or (Phenylseleno)Formate Mediated Ring Closures. Tetrahedron 2000, 56, 3995–4000. [Google Scholar] [CrossRef]
- Ng, H.H.; Leo, C.H.; O’Sullivan, K.; Alexander, S.-A.; Davies, M.J.; Schiesser, C.H.; Parry, L.J. 1,4-Anhydro-4-Seleno-d-Talitol (SeTal) Protects Endothelial Function in the Mouse Aorta by Scavenging Superoxide Radicals under Conditions of Acute Oxidative Stress. Biochem. Pharmacol. 2017, 128, 34–45. [Google Scholar] [CrossRef]
- Carroll, L.; Pattison, D.I.; Fu, S.; Schiesser, C.H.; Davies, M.J.; Hawkins, C.L. Catalytic Oxidant Scavenging by Selenium-Containing Compounds: Reduction of Selenoxides and N-Chloramines by Thiols and Redox Enzymes. Redox Biol. 2017, 12, 872–882. [Google Scholar] [CrossRef]
- Kötzler, M.P.; Hancock, S.M.; Withers, S.G. Glycosidases: Functions, Families and Folds. In Encyclopedia of Life Sciences (eLS); John Wiley & Sons, Ltd.: Chichester, UK, 2014; p. a0020548. ISBN 978-0-470-01590-2. [Google Scholar]
- Gerber-Lemaire, S.; Juillerat-Jeanneret, L. Glycosylation Pathways as Drug Targets for Cancer: Glycosidase Inhibitors. Mini Rev. Med. Chem. 2006, 6, 1043–1052. [Google Scholar] [CrossRef]
- Kónya, Z.; Bécsi, B.; Kiss, A.; Tamás, I.; Lontay, B.; Szilágyi, L.; Kövér, K.E.; Erdődi, F. Aralkyl Selenoglycosides and Related Selenosugars in Acetylated Form Activate Protein Phosphatase-1 and -2A. Bioorg. Med. Chem. 2018, 26, 1875–1884. [Google Scholar] [CrossRef]
- Davies, M.J.; Schiesser, C.H. 1,4-Anhydro-4-Seleno-d-Talitol (SeTal): A Remarkable Selenium-Containing Therapeutic Molecule. New J. Chem. 2019, 43, 9759–9765. [Google Scholar] [CrossRef]
- Jeong, L.S.; Tosh, D.K.; Choi, W.J.; Lee, S.K.; Kang, Y.-J.; Choi, S.; Lee, J.H.; Lee, H.; Lee, H.W.; Kim, H.O. Discovery of a New Template for Anticancer Agents: 2′-Deoxy-2′-Fluoro-4′-Selenoarabinofuranosyl-Cytosine (2′-F-4′-Seleno-Ara-C). J. Med. Chem. 2009, 52, 5303–5306. [Google Scholar] [CrossRef]
- Jeong, L.S.; Choi, Y.N.; Tosh, D.K.; Choi, W.J.; Kim, H.O.; Choi, J. Design and Synthesis of Novel 2′,3′-Dideoxy-4′-Selenonucleosides as Potential Antiviral Agents. Bioorg. Med. Chem. 2008, 16, 9891–9897. [Google Scholar] [CrossRef]
- Kumar, A.A.; Illyés, T.Z.; Kövér, K.E.; Szilágyi, L. Convenient Syntheses of 1,2-Trans Selenoglycosides Using Isoselenuronium Salts as Glycosylselenenyl Transfer Reagents. Carbohydr. Res. 2012, 360, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.; Mario Pinto, B. Phenylselenoglycosides as Novel, Versatile Glycosyl Donors. Selective Activation over Thioglycosides. Tetrahedron Lett. 1991, 32, 4435–4438. [Google Scholar] [CrossRef]
- Mehta, S.; Pinto, B.M. Novel Glycosidation Methodology. The Use of Phenyl Selenoglycosides as Glycosyl Donors and Acceptors in Oligosaccharide Synthesis. J. Org. Chem. 1993, 58, 3269–3276. [Google Scholar] [CrossRef]
- Nanami, M.; Ando, H.; Kawai, Y.; Kokets, M.; Ishihara, H. Stereoselective Synthesis of Various α-Selenoglycosides Using in Situ Production of α-Selenolate Anion. Tetrahedron Lett. 2007, 48, 1113–1116. [Google Scholar] [CrossRef]
- Braga, A.L.; Severo Filho, W.A.; Schwab, R.S.; Rodrigues, O.E.D.; Dornelles, L.; Braga, H.C.; Lüdtke, D.S. Synthesis of Selenium- and Tellurium-Containing Nucleosides Derived from Uridine. Tetrahedron Lett. 2009, 50, 3005–3007. [Google Scholar] [CrossRef]
- Braga, H.C.; Wouters, A.D.; Zerillo, F.B.; Lüdtke, D.S. Synthesis of Seleno-Carbohydrates Derived from D-Galactose. Carbohydr. Res. 2010, 345, 2328–2333. [Google Scholar] [CrossRef]
- Vargas, J.P.; Pinto, L.M.; Savegnago, L.; Lüdtke, D.S. Synthesis of Alkylseleno-Carbohydrates and Evaluation of Their Antioxidant Properties. J. Braz. Chem. Soc. 2015. [Google Scholar] [CrossRef]
- Yue, C.; Chen, J.; Hou, R.; Liu, J.; Li, X.; Gao, Z.; Liu, C.; Wang, D.; Lu, Y.; Li, H.; et al. Effects of Selenylation Modification on Antioxidative Activities of Schisandra Chinensis Polysaccharide. PLoS ONE 2015, 10, e0134363. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, B.; Wang, X.; Yao, J.; Zhang, J. Synthesis of Selenium-Containing Polysaccharides and Evaluation of Antioxidant Activity in Vitro. Int. J. Biol. Macromol. 2012, 51, 987–991. [Google Scholar] [CrossRef]
- Wang, L.; Li, C.; Huang, Q.; Fu, X. Biofunctionalization of Selenium Nanoparticles with a Polysaccharide from Rosa Roxburghii Fruit and Their Protective Effect against H2O2-Induced Apoptosis in INS-1 Cells. Food Funct. 2019, 10, 539–553. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, J.; Rao, S.; Su, Y.; Li, J.; Li, C.; Xu, S.; Yang, Y. Antidiabetic Activity of Mycelia Selenium-Polysaccharide from Catathelasma Ventricosum in STZ-Induced Diabetic Mice. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 62, 285–291. [Google Scholar] [CrossRef]
- Jin, Y.; Zhang, L.; Zhang, M.; Chen, L.; Cheung, P.C.K.; Oi, V.E.C.; Lin, Y. Antitumor Activities of Heteropolysaccharides of Poria Cocos Mycelia from Different Strains and Culture Media. Carbohydr. Res. 2003, 338, 1517–1521. [Google Scholar] [CrossRef]
- Gan, D.; Ma, L.; Jiang, C.; Xu, R.; Zeng, X. Production, Preliminary Characterization and Antitumor Activity in Vitro of Polysaccharides from the Mycelium of Pholiota Dinghuensis Bi. Carbohydr. Polym. 2011, 84, 997–1003. [Google Scholar] [CrossRef]
- Savic, M.; Klimaszewska, M.; Bamburowicz-Klimkowska, M.; Suchocki, P.; Niksic, M.; Szutowski, M.; Wroczynski, P.; Turlo, J. A Search for the Optimum Selenium Source to Obtain Mushroom-Derived Chemopreventive Preparations. Int. J. Med. Mushrooms 2016, 18, 279–289. [Google Scholar] [CrossRef]
- Xu, C.L.; Wang, Y.Z.; Jin, M.L.; Yang, X.Q. Preparation, Characterization and Immunomodulatory Activity of Selenium-Enriched Exopolysaccharide Produced by Bacterium Enterobacter Cloacae Z0206. Bioresour. Technol. 2009, 100, 2095–2097. [Google Scholar] [CrossRef]
- Li, Z.-Y.; Guo, S.-Y.; Li, L. Bioeffects of Selenite on the Growth of Spirulina Platensis and Its Biotransformation. Bioresour. Technol. 2003, 89, 171–176. [Google Scholar] [CrossRef]
- Turlo, J.; Gutkowska, B.; Herold, F. Effect of Selenium Enrichment on Antioxidant Activities and Chemical Composition of Lentinula Edodes (Berk.) Pegl. Mycelial Extracts. Food Chem. Toxicol. 2010, 48, 1085–1091. [Google Scholar] [CrossRef]
- Turło, J.; Gutkowska, B.; Herold, F.; Klimaszewska, M.; Suchocki, P. Optimization of Selenium-Enriched Mycelium of Lentinula Edodes (Berk.) Pegler as a Food Supplement. Food Biotechnol. 2010, 24, 180–196. [Google Scholar] [CrossRef]
- Turło, J.; Gutkowska, B.; Herold, F.; Gajzlerska, W.; Dawidowski, M.; Dorociak, A.; Zobel, A. Biological Availability and Preliminary Selenium Speciation in Selenium-Enriched Mycelium of Lentinula Edodes (Berk.). Food Biotechnol. 2011, 25, 16–29. [Google Scholar] [CrossRef]
- Liu, J.; Chen, X.; Yue, C.; Hou, R.; Chen, J.; Lu, Y.; Li, X.; Li, R.; Liu, C.; Gao, Z.; et al. Effect of Selenylation Modification on Immune-Enhancing Activity of Atractylodes Macrocephala Polysaccharide. Int. J. Biol. Macromol. 2015, 72, 1435–1440. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, F.-X.; Liu, Z.-W.; Zhang, S.-T.; Zhang, Y.-Y.; Liang, J.-Y.; Wang, Y.-P. Synthesis and Characterisation of Seleno-Cynomorium Songaricum Rupr. Polysaccharide. Nat. Prod. Res. 2009, 23, 1641–1651. [Google Scholar] [CrossRef]
- Wang, J.; Li, Q.; Bao, A.; Liu, X.; Zeng, J.; Yang, X.; Yao, J.; Zhang, J.; Lei, Z. Synthesis of Selenium-Containing Artemisia Sphaerocephala Polysaccharides: Solution Conformation and Anti-Tumor Activities in Vitro. Carbohydr. Polym. 2016, 152, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Chen, J.; Qiu, S.; Li, Y.; Wang, D.; Liu, C.; Li, X.; Hou, R.; Yue, C.; Liu, J.; et al. Optimization of Selenylation Modification for Garlic Polysaccharide Based on Immune-Enhancing Activity. Carbohydr. Polym. 2016, 136, 560–569. [Google Scholar] [CrossRef]
- Liu, L.; Pan, D.; Zeng, X.; Li, H. Effect of Selenium-Enriched Exopolysaccharide Produced by Lactococcus Lactis Subsp. Lactis on Signaling Molecules in Mouse Spleen Lymphocytes. Food Funct. 2013, 4, 1489–1495. [Google Scholar] [CrossRef]
- Pan, D.; Liu, J.; Zeng, X.; Liu, L.; Li, H.; Guo, Y. Immunomodulatory Activity of Selenium Exopolysaccharide Produced by Lactococcus lactis Subsp. Lactis. Food Agric. Immunol. 2015, 26, 248–259. [Google Scholar] [CrossRef]
- Huang, G.-C.; Lee, C.-J.; Wang, K.-T.; Weng, B.-C.; Chien, T.-Y.; Tseng, S.-H.; Wang, C.-C. Immunomodulatory Effects of Hedysarum Polybotrys Extract in Mice Macrophages, Splenocytes and Leucopenia. Molecules 2013, 18, 14862–14875. [Google Scholar] [CrossRef]
- Zhu, Z.-Y.; Liu, F.; Gao, H.; Sun, H.; Meng, M.; Zhang, Y.-M. Synthesis, Characterization and Antioxidant Activity of Selenium Polysaccharide from Cordyceps Militaris. Int. J. Biol. Macromol. 2016, 93, 1090–1099. [Google Scholar] [CrossRef]
- Liu, Y.; You, Y.; Li, Y.; Zhang, L.; Yin, L.; Shen, Y.; Li, C.; Chen, H.; Chen, S.; Hu, B.; et al. The Characterization, Selenylation and Antidiabetic Activity of Mycelial Polysaccharides from Catathelasma Ventricosum. Carbohydr. Polym. 2017, 174, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Duan, K.; Shan, H. Selenylation Modification of Degraded Polysaccharide from Enteromorpha Prolifera and Its Biological Activities. J. Ocean Univ. China 2018, 17, 445–450. [Google Scholar] [CrossRef]
- Surhio, M.M.; Wang, Y.; Xu, P.; Shah, F.; Li, J.; Ye, M. Antihyperlipidemic and Hepatoprotective Properties of Selenium Modified Polysaccharide from Lachnum Sp. Int. J. Biol. Macromol. 2017, 99, 88–95. [Google Scholar] [CrossRef]
- Ren, G.; Li, K.; Hu, Y.; Yu, M.; Qu, J.; Xu, X. Optimization of Selenizing Conditions for Seleno-Lentinan and Its Characteristics. Int. J. Biol. Macromol. 2015, 81, 249–258. [Google Scholar] [CrossRef]
- Hou, R.; Chen, J.; Yue, C.; Li, X.; Liu, J.; Gao, Z.; Liu, C.; Lu, Y.; Wang, D.; Li, H.; et al. Modification of Lily Polysaccharide by Selenylation and the Immune-Enhancing Activity. Carbohydr. Polym. 2016, 142, 73–81. [Google Scholar] [CrossRef]
- Chen, W.; Chen, J.; Wu, H.; Gou, Y.; Hu, F.; Liu, L.; Gao, X.; Zhang, P. Optimization of Selenylation Conditions for a Pectic Polysaccharide and Its Structural Characteristic. Int. J. Biol. Macromol. 2014, 69, 244–251. [Google Scholar] [CrossRef]
- Qiu, S.; Chen, J.; Chen, X.; Fan, Q.; Zhang, C.; Wang, D.; Li, X.; Chen, X.; Chen, X.; Liu, C.; et al. Optimization of Selenylation Conditions for Lycium Barbarum Polysaccharide Based on Antioxidant Activity. Carbohydr. Polym. 2014, 103, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Hamid, M.; Liu, D.; Abdulrahim, Y.; Liu, Y.; Qian, G.; Khan, A.; Gan, F.; Huang, K. Amelioration of CCl4-Induced Liver Injury in Rats by Selenizing Astragalus Polysaccharides: Role of Proinflammatory Cytokines, Oxidative Stress and Hepatic Stellate Cells. Res. Vet. Sci. 2017, 114, 202–211. [Google Scholar] [CrossRef]
- Qin, T.; Ren, Z.; Huang, Y.; Song, Y.; Lin, D.; Li, J.; Ma, Y.; Wu, X.; Qiu, F.; Xiao, Q. Selenizing Hericium Erinaceus Polysaccharides Induces Dendritic Cells Maturation through MAPK and NF-ΚB Signaling Pathways. Int. J. Biol. Macromol. 2017, 97, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Lian, K.-X.; Zhu, X.-Q.; Chen, J.; Liu, G.; Gu, X.-L. Selenylation Modification: Enhancement of the Antioxidant Activity of a Glycyrrhiza Uralensis Polysaccharide. Glycoconj. J. 2018, 35, 243–253. [Google Scholar] [CrossRef]
- Li, X.; Hou, R.; Yue, C.; Liu, J.; Gao, Z.; Chen, J.; Lu, Y.; Wang, D.; Liu, C.; Hu, Y. The Selenylation Modification of Epimedium Polysaccharide and Isatis Root Polysaccharide and the Immune-Enhancing Activity Comparison of Their Modifiers. Biol. Trace Elem. Res. 2016, 171, 224–234. [Google Scholar] [CrossRef]
- Ji, Y.-B.; Dong, F.; Lang, L.; Zhang, L.-W.; Miao, J.; Liu, Z.-F.; Jin, L.-N.; Hao, Y. Optimization of Synthesis, Characterization and Cytotoxic Activity of Seleno-Capparis Spionosa L. Polysaccharide. Int. J. Mol. Sci. 2012, 13, 17275–17289. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhang, B.; Fu, X.; Liu, R.H. A Novel Polysaccharide Isolated from Mulberry Fruits (Murus alba L.) and its Selenide Derivative: Structural Characterization and Biological Activities. Food Funct. 2016, 7, 2886–2897. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Wang, C.; Zhang, S.; Li, S.; Zhou, G.; Wang, S.; Zhang, J. Extraction, Selenylation Modification and Antitumor Activity of the Glucan from Castanea Mollissima Blume. Glycoconj. J. 2017, 34, 207–217. [Google Scholar] [CrossRef]
- Ding, G.-B.; Nie, R.-H.; Lv, L.-H.; Wei, G.-Q.; Zhao, L.-Q. Preparation and Biological Evaluation of a Novel Selenium-Containing Exopolysaccharide from Rhizobium Sp. N613. Carbohydr. Polym. 2014, 109, 28–34. [Google Scholar] [CrossRef]
- Zhao, B.; Zhang, J.; Yao, J.; Song, S.; Yin, Z.; Gao, Q. Selenylation Modification Can Enhance Antioxidant Activity of Potentilla Anserina L. Polysaccharide. Int. J. Biol. Macromol. 2013, 58, 320–328. [Google Scholar] [CrossRef]
- Qiu, S.; Chen, J.; Qin, T.; Hu, Y.; Wang, D.; Fan, Q.; Zhang, C.; Chen, X.; Chen, X.; Liu, C.; et al. Effects of Selenylation Modification on Immune-Enhancing Activity of Garlic Polysaccharide. PLoS ONE 2014, 9, e86377. [Google Scholar] [CrossRef]
- Wang, D.; Zhao, Y.; Sun, Y.; Yang, X. Protective Effects of Ziyang Tea Polysaccharides on CCl4-Induced Oxidative Liver Damage in Mice. Food Chem. 2014, 143, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, Y.; He, X.; Wei, X. Preparation, Structural Characterization and Bioactivities of Se-Containing Polysaccharide: A Review. Int. J. Biol. Macromol. 2018, 120, 82–92. [Google Scholar] [CrossRef]
- Shi, L.; Xun, W.; Yue, W.; Zhang, C.; Ren, Y.; Shi, L.; Wang, Q.; Yang, R.; Lei, F. Effect of Sodium Selenite, Se-Yeast and Nano-Elemental Selenium on Growth Performance, Se Concentration and Antioxidant Status in Growing Male Goats. Small Rumin. Res. 2011, 96, 49–52. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, J.; Yu, H. Elemental Selenium at Nano Size Possesses Lower Toxicity without Compromising the Fundamental Effect on Selenoenzymes: Comparison with Selenomethionine in Mice. Free Radic. Biol. Med. 2007, 42, 1524–1533. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Xu, T. Elemental Selenium at Nano Size (Nano-Se) as a Potential Chemopreventive Agent with Reduced Risk of Selenium Toxicity: Comparison with Se-Methylselenocysteine in Mice. Toxicol. Sci. Off. J. Soc. Toxicol. 2008, 101, 22–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.S.; Gao, X.Y.; Zhang, L.D.; Bao, Y.P. Biological Effects of a Nano Red Elemental Selenium. BioFactors Oxf. Engl. 2001, 15, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Vert, M.; Doi, Y.; Hellwich, K.-H.; Hess, M.; Hodge, P.; Kubisa, P.; Rinaudo, M.; Schué, F. Terminology for Biorelated Polymers and Applications (IUPAC Recommendations 2012). Pure Appl. Chem. 2012, 84, 377–410. [Google Scholar] [CrossRef]
- Torres, S.K.; Campos, V.L.; León, C.G.; Rodríguez-Llamazares, S.M.; Rojas, S.M.; González, M.; Smith, C.; Mondaca, M.A. Biosynthesis of Selenium Nanoparticles by Pantoea Agglomerans and Their Antioxidant Activity. J. Nanoparticle Res. 2012, 14, 1236. [Google Scholar] [CrossRef]
- Desai, M.P.; Labhasetwar, V.; Walter, E.; Levy, R.J.; Amidon, G.L. The Mechanism of Uptake of Biodegradable Microparticles in Caco-2 Cells Is Size Dependent. Pharm. Res. 1997, 14, 1568–1573. [Google Scholar] [CrossRef]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, B.; Peng, Q.; Baron, M.; Melcova, M.; Opatrilova, R.; Zidkova, J.; et al. Nano-Selenium and Its Nanomedicine Applications: A Critical Review. Int. J. Nanomed. 2018, 13, 2107–2128. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhai, X.; Zhao, G.; Ren, F.; Leng, X. Synthesis, Characterization, and Controlled Release of Selenium Nanoparticles Stabilized by Chitosan of Different Molecular Weights. Carbohydr. Polym. 2015, 134, 158–166. [Google Scholar] [CrossRef]
- Chen, W.; Li, Y.; Yang, S.; Yue, L.; Jiang, Q.; Xia, W. Synthesis and Antioxidant Properties of Chitosan and Carboxymethyl Chitosan-Stabilized Selenium Nanoparticles. Carbohydr. Polym. 2015, 132, 574–581. [Google Scholar] [CrossRef]
- Sun, Y.; Nie, Y.; Wang, Z.; Hua, C.; Wang, R.; Gao, J. Biomacromolecule-Directed Synthesis and Characterization of Selenium Nanoparticles and Their Compatibility with Bacterial and Eukaryotic Cells. Nanosci. Nanotechnol. Lett. 2017, 9, 1987–1991. [Google Scholar] [CrossRef]
- Xu, D.; Yang, L.; Wang, Y.; Wang, G.; Rensing, C.; Zheng, S. Proteins Enriched in Charged Amino Acids Control the Formation and Stabilization of Selenium Nanoparticles in Comamonas Testosteroni S44. Sci. Rep. 2018, 8, 4766. [Google Scholar] [CrossRef]
- Liu, C.; Fu, Y.; Li, C.-E.; Chen, T.; Li, X. Phycocyanin-Functionalized Selenium Nanoparticles Reverse Palmitic Acid-Induced Pancreatic β Cell Apoptosis by Enhancing Cellular Uptake and Blocking Reactive Oxygen Species (ROS)-Mediated Mitochondria Dysfunction. J. Agric. Food Chem. 2017, 65, 4405–4413. [Google Scholar] [CrossRef]
- Zhu, C.; Zhang, S.; Song, C.; Zhang, Y.; Ling, Q.; Hoffmann, P.R.; Li, J.; Chen, T.; Zheng, W.; Huang, Z. Selenium Nanoparticles Decorated with Ulva Lactuca Polysaccharide Potentially Attenuate Colitis by Inhibiting NF-ΚB Mediated Hyper Inflammation. J. Nanobiotechnol. 2017, 15, 20. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Yu, Z.; Lin, Z.; Lei, Z.; Ning, Z.; Regenstein, J.M.; Yang, J.; Ren, J. Biofunctionalization of Selenium Nanoparticle with Dictyophora Indusiata Polysaccharide and Its Antiproliferative Activity through Death-Receptor and Mitochondria-Mediated Apoptotic Pathways. Sci. Rep. 2015, 5, 18629. [Google Scholar] [CrossRef] [PubMed]
- Ping, Z.; Liu, T.; Xu, H.; Meng, Y.; Li, W.; Xu, X.; Zhang, L. Construction of Highly Stable Selenium Nanoparticles Embedded in Hollow Nanofibers of Polysaccharide and Their Antitumor Activities. Nano Res. 2017, 10, 3775–3789. [Google Scholar] [CrossRef]
- Wu, H.; Zhu, H.; Li, X.; Liu, Z.; Zheng, W.; Chen, T.; Yu, B.; Wong, K.-H. Induction of Apoptosis and Cell Cycle Arrest in A549 Human Lung Adenocarcinoma Cells by Surface-Capping Selenium Nanoparticles: An Effect Enhanced by Polysaccharide-Protein Complexes from Polyporus Rhinocerus. J. Agric. Food Chem. 2013, 61, 9859–9866. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Li, X.; Liu, W.; Chen, T.; Li, Y.; Zheng, W.; Man, C.W.-Y.; Wong, M.-K.; Wong, K.-H. Surface Decoration of Selenium Nanoparticles by Mushroom Polysaccharides–Protein Complexes to Achieve Enhanced Cellular Uptake and Antiproliferative Activity. J. Mater. Chem. 2012, 22, 9602. [Google Scholar] [CrossRef]
- Hong, A.; Rao, L.; Zhuang, M.; Luo, T.; Wang, Y.; Ma, Y. Chitosan-Decorated Selenium Nanoparticles as Proteins Carriers to Improve the in Vivo Half-Life of the Peptide Therapeutic BAY 55-9837 for Type 2 Diabetes Mellitus. Int. J. Nanomed. 2014, 4819. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Wang, Y.; Zhou, Y.; Li, W.; Zheng, W. Modification and Modulation of Saccharides on Elemental Selenium Nanoparticles in Liquid Phase. Mater. Lett. 2008, 62, 2311–2314. [Google Scholar] [CrossRef]
- Zhang, S.-Y.; Zhang, J.; Wang, H.-Y.; Chen, H.-Y. Synthesis of Selenium Nanoparticles in the Presence of Polysaccharides. Mater. Lett. 2004, 58, 2590–2594. [Google Scholar] [CrossRef]
- Shoeibi, S.; Mashreghi, M. Biosynthesis of Selenium Nanoparticles Using Enterococcus Faecalis and Evaluation of Their Antibacterial Activities. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. GMS 2017, 39, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Ramamurthy, C.; Sampath, K.S.; Arunkumar, P.; Kumar, M.S.; Sujatha, V.; Premkumar, K.; Thirunavukkarasu, C. Green Synthesis and Characterization of Selenium Nanoparticles and Its Augmented Cytotoxicity with Doxorubicin on Cancer Cells. Bioprocess Biosyst. Eng. 2013, 36, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Stevanović, M.; Filipović, N.; Djurdjević, J.; Lukić, M.; Milenković, M.; Boccaccini, A. 45S5Bioglass®-Based Scaffolds Coated with Selenium Nanoparticles or with Poly(Lactide-Co-Glycolide)/Selenium Particles: Processing, Evaluation and Antibacterial Activity. Colloids Surf. B Biointerfaces 2015, 132, 208–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanhua, W.; Hao, H.; Li, Y.; Zhang, S. Selenium-Substituted Hydroxyapatite Nanoparticles and Their in Vivo Antitumor Effect on Hepatocellular Carcinoma. Colloids Surf. B Biointerfaces 2016, 140, 297–306. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, W.; Zhao, Z.; Li, N.; Mou, Z.; Sun, D.; Cai, Y.; Wang, W.; Lin, Y. Quercetin Loading CdSe/ZnS Nanoparticles as Efficient Antibacterial and Anticancer Materials. J. Inorg. Biochem. 2017, 167, 36–48. [Google Scholar] [CrossRef]
- Yin, J.; Hou, Y.; Yin, Y.; Song, X. Selenium-Coated Nanostructured Lipid Carriers Used for Oral Delivery of Berberine to Accomplish a Synergic Hypoglycemic Effect. Int. J. Nanomed. 2017, 12, 8671–8680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.S.; Kulkarni, A.; Khurana, A.; Kaur, J.; Tikoo, K. Selenium Nanoparticles Involve HSP-70 and SIRT1 in Preventing the Progression of Type 1 Diabetic Nephropathy. Chem. Biol. Interact. 2014, 223, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, S.; Welling, M.N.; Mantri, S.B.; Desai, K. In Vitro and in Vivo Antioxidant, Cytotoxic, and Anti-Chronic Inflammatory Arthritic Effect of Selenium Nanoparticles. J. Biomed. Mater. Res. B Appl. Biomater. 2016, 104, 993–1003. [Google Scholar] [CrossRef]
- Ramya, S.; Shanmugasundaram, T.; Balagurunathan, R. Biomedical Potential of Actinobacterially Synthesized Selenium Nanoparticles with Special Reference to Anti-Biofilm, Anti-Oxidant, Wound Healing, Cytotoxic and Anti-Viral Activities. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. GMS 2015, 32, 30–39. [Google Scholar] [CrossRef]
- Wu, X.; Yao, J.; Yang, Z.; Yue, W.; Ren, Y.; Zhang, C.; Liu, X.; Wang, H.; Zhao, X.; Yuan, S.; et al. Improved Fetal Hair Follicle Development by Maternal Supplement of Selenium at Nano Size (Nano-Se). Livest. Sci. 2011, 142, 270–275. [Google Scholar] [CrossRef]
- Shi, L.; Yang, R.; Yue, W.; Xun, W.; Zhang, C.; Ren, Y.; Shi, L.; Lei, F. Effect of Elemental Nano-Selenium on Semen Quality, Glutathione Peroxidase Activity, and Testis Ultrastructure in Male Boer Goats. Anim. Reprod. Sci. 2010, 118, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Majhi, S.; Das, D. Chemical Derivatization of Natural Products: Semisynthesis and Pharmacological Aspects-A Decade Update. Tetrahedron 2021, 78, 131801. [Google Scholar] [CrossRef]
- Armaly, A.M.; DePorre, Y.C.; Groso, E.J.; Riehl, P.S.; Schindler, C.S. Discovery of Novel Synthetic Methodologies and Reagents during Natural Product Synthesis in the Post-Palytoxin Era. Chem. Rev. 2015, 115, 9232–9276. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Sorensen, E.J. Classics in Total Synthesis: Targets, Strategies, Methods; VCH: Weinheim, Germany; New York, NY, USA, 1996; ISBN 978-3-527-29284-4. [Google Scholar]
- Xiao, R.; Grinstaff, M.W. Chemical Synthesis of Polysaccharides and Polysaccharide Mimetics. Prog. Polym. Sci. 2017, 74, 78–116. [Google Scholar] [CrossRef]
- Ren, Y.; Bai, Y.; Zhang, Z.; Cai, W.; Del Rio Flores, A. The Preparation and Structure Analysis Methods of Natural Polysaccharides of Plants and Fungi: A Review of Recent Development. Molecules 2019, 24, 3122. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Chen, J.; Li, J.; Yan, L.; Li, S.; Ye, X.; Liu, D.; Ding, T.; Linhardt, R.J.; Orfila, C.; et al. Extraction and Characterization of RG-I Enriched Pectic Polysaccharides from Mandarin Citrus Peel. Food Hydrocoll. 2018, 79, 579–586. [Google Scholar] [CrossRef]
- Song, Y.-R.; Sung, S.-K.; Jang, M.; Lim, T.-G.; Cho, C.-W.; Han, C.-J.; Hong, H.-D. Enzyme-Assisted Extraction, Chemical Characteristics, and Immunostimulatory Activity of Polysaccharides from Korean Ginseng (Panax Ginseng Meyer). Int. J. Biol. Macromol. 2018, 116, 1089–1097. [Google Scholar] [CrossRef]
- Zhang, M.; Cui, S.W.; Cheung, P.C.K.; Wang, Q. Antitumor Polysaccharides from Mushrooms: A Review on Their Isolation Process, Structural Characteristics and Antitumor Activity. Trends Food Sci. Technol. 2007, 18, 4–19. [Google Scholar] [CrossRef]
- e Silva, A.d.S.; de Magalhães, W.T.; Moreira, L.M.; Rocha, M.V.P.; Bastos, A.K.P. Microwave-Assisted Extraction of Polysaccharides from Arthrospira (Spirulina) Platensis Using the Concept of Green Chemistry. Algal Res. 2018, 35, 178–184. [Google Scholar] [CrossRef]
- Sukhithasri, V.; Nisha, N.; Biswas, L.; Anil Kumar, V.; Biswas, R. Innate Immune Recognition of Microbial Cell Wall Components and Microbial Strategies to Evade Such Recognitions. Microbiol. Res. 2013, 168, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Choong, Y.-K.; Ellan, K.; Chen, X.-D.; Azuar Mohamad, S. Extraction and Fractionation of Polysaccharides from a Selected Mushroom Species, Ganoderma lucidum: A Critical Review. In Fractionation; Al-Haj Ibrahim, H., Ed.; IntechOpen: London, UK, 2019; ISBN 978-1-78984-964-6. [Google Scholar]
- Segneanu, A.-E.; Sziple, F.; Vlazan, P.; Sfarloaga, P.; Grozesku, I.; Daniel, V. Biomass Extraction Methods. In Biomass Now—Sustainable Growth and Use; Matovic, M.D., Ed.; InTech: Rijeka, Croatia, 2013; ISBN 978-953-51-1105-4. [Google Scholar]
- Ruthes, A.C.; Smiderle, F.R.; Iacomini, M. D-Glucans from Edible Mushrooms: A Review on the Extraction, Purification and Chemical Characterization Approaches. Carbohydr. Polym. 2015, 117, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Du, B.; Bian, Z.; Xu, B. Beta-Glucans from Edible and Medicinal Mushrooms: Characteristics, Physicochemical and Biological Activities. J. Food Compos. Anal. 2015, 41, 165–173. [Google Scholar] [CrossRef]
- Villares, A.; Mateo-Vivaracho, L.; Guillamón, E. Structural Features and Healthy Properties of Polysaccharides Occurring in Mushrooms. Agriculture 2012, 2, 452–471. [Google Scholar] [CrossRef] [Green Version]
- Zou, P.; Yang, X.; Huang, W.-W.; Zhao, H.-T.; Wang, J.; Xu, R.-B.; Hu, X.-L.; Shen, S.-Y.; Qin, D. Characterization and Bioactivity of Polysaccharides Obtained from Pine Cones of Pinus Koraiensis by Graded Ethanol Precipitation. Molecules 2013, 18, 9933–9948. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.-H.; Shen, M.-Y.; Nie, S.-P.; Liu, X.; Zhang, H.; Xie, M.-Y. Analysis of Monosaccharide Composition of Cyclocarya Paliurus Polysaccharide with Anion Exchange Chromatography. Carbohydr. Polym. 2013, 98, 976–981. [Google Scholar] [CrossRef]
- Turło, J. The Biotechnology of Higher Fungi—Current State and Perspectives. Folia Biol. Oecologica 2014, 10, 49–65. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhong, J. Production of Ginseng and Its Bioactive Components in Plant Cell Culture: Current Technological and Applied Aspects. J. Biotechnol. 1999, 68, 89–99. [Google Scholar] [CrossRef]
- Sun, Y.; Zhou, C.; Huang, S.; Jiang, C. Selenium Polysaccharide SPMP-2a from Pleurotus Geesteranus Alleviates H2O2-Induced Oxidative Damage in HaCaT Cells. BioMed Res. Int. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.; Zhang, J.; Liu, Z.; Zhang, Y.; Li, J.; Li, Y.O. Biosynthesis of Selenium Rich Exopolysaccharide (Se-EPS) by Pseudomonas PT-8 and Characterization of Its Antioxidant Activities. Carbohydr. Polym. 2016, 142, 230–239. [Google Scholar] [CrossRef]
- Xing, H.; Du, R.; Zhao, F.; Han, Y.; Xiao, H.; Zhou, Z. Optimization, Chain Conformation and Characterization of Exopolysaccharide Isolated from Leuconostoc Mesenteroides DRP105. Int. J. Biol. Macromol. 2018, 112, 1208–1216. [Google Scholar] [CrossRef]
- Cumpstey, I. Chemical Modification of Polysaccharides. ISRN Org. Chem. 2013, 2013, 1–27. [Google Scholar] [CrossRef]
- Yalpani, M. A Survey of Recent Advances in Selective Chemical and Enzymic Polysaccharide Modifications. Tetrahedron 1985, 41, 2957–3020. [Google Scholar] [CrossRef]
- Lee, J.-H.; Lee, Y.-K.; Chang, Y.H. Effects of Selenylation Modification on Structural and Antioxidant Properties of Pectic Polysaccharides Extracted from Ulmus Pumila L. Int. J. Biol. Macromol. 2017, 104, 1124–1132. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Chen, J.; Wang, D.; Hu, Y.; Wang, M.; Zhang, J.; Nguyen, T.L.; Liu, C.; Liu, X. Optimization of Selenylation Conditions for Chinese Angelica Polysaccharide Based on Immune-Enhancing Activity. Carbohydr. Polym. 2013, 92, 645–650. [Google Scholar] [CrossRef]
- Gao, Z.; Liu, K.; Tian, W.; Wang, H.; Liu, Z.; Li, Y.; Li, E.; Liu, C.; Li, X.; Hou, R.; et al. Effects of Selenizing Angelica Polysaccharide and Selenizing Garlic Polysaccharide on Immune Function of Murine Peritoneal Macrophage. Int. Immunopharmacol. 2015, 27, 104–109. [Google Scholar] [CrossRef]
- Yuan, B.; Yang, X.-Q.; Kou, M.; Lu, C.-Y.; Wang, Y.-Y.; Peng, J.; Chen, P.; Jiang, J.-H. Selenylation of Polysaccharide from the Sweet Potato and Evaluation of Antioxidant, Antitumor, and Antidiabetic Activities. J. Agric. Food Chem. 2017, 65, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, X.; Bao, A.; Liu, X.; Zeng, J.; Liu, X.; Yao, J.; Zhang, J.; Lei, Z. Microwave-Assisted Synthesis, Structure and Anti-Tumor Activity of Selenized Artemisia Sphaerocephala Polysaccharide. Int. J. Biol. Macromol. 2017, 95, 1108–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Zhao, Q.; Wu, Q.; Zhang, H.; Kong, W.; Liang, J.; Yao, J.; Zhang, J.; Wang, J. Efficient Synthesis of Polysaccharide with High Selenium Content Mediated by Imidazole-Based Acidic Ionic Liquids. Carbohydr. Polym. 2019, 203, 157–166. [Google Scholar] [CrossRef]
- Chen, L.; Huang, G. The Antiviral Activity of Polysaccharides and Their Derivatives. Int. J. Biol. Macromol. 2018, 115, 77–82. [Google Scholar] [CrossRef]
- Huang, S.; Yang, W.; Huang, G. Preparation and Activities of Selenium Polysaccharide from Plant Such as Grifola Frondosa. Carbohydr. Polym. 2020, 242, 116409. [Google Scholar] [CrossRef] [PubMed]
- Quintana, M.; Haro-Poniatowski, E.; Morales, J.; Batina, N. Synthesis of Selenium Nanoparticles by Pulsed Laser Ablation. Appl. Surf. Sci. 2002, 195, 175–186. [Google Scholar] [CrossRef]
- Jain, R.; Gonzalez-Gil, G.; Singh, V.; Hullebusch, E.D.V.; Farges, F.; Lens, P.N.L. Biogenic Selenium Nanoparticles: Production, Characterization and Challenges. In Biotechnology; Studium Press LLC.: New Delhi, India, 2014; Volume 10, pp. 365–394. [Google Scholar] [CrossRef]
- Yu, B.; You, P.; Song, M.; Zhou, Y.; Yu, F.; Zheng, W. A Facile and Fast Synthetic Approach to Create Selenium Nanoparticles with Diverse Shapes and Their Antioxidation Ability. New J. Chem. 2016, 40, 1118–1123. [Google Scholar] [CrossRef]
- Li, Y.; Lin, Z.; Guo, M.; Xia, Y.; Zhao, M.; Wang, C.; Xu, T.; Chen, T.; Zhu, B. Inhibitory Activity of Selenium Nanoparticles Functionalized with Oseltamivir on H1N1 Influenza Virus. Int. J. Nanomed. 2017, 12, 5733–5743. [Google Scholar] [CrossRef] [Green Version]
- Cremonini, E.; Zonaro, E.; Donini, M.; Lampis, S.; Boaretti, M.; Dusi, S.; Melotti, P.; Lleo, M.M.; Vallini, G. Biogenic Selenium Nanoparticles: Characterization, Antimicrobial Activity and Effects on Human Dendritic Cells and Fibroblasts. Microb. Biotechnol. 2016, 9, 758–771. [Google Scholar] [CrossRef]
- Piacenza, E.; Presentato, A.; Zonaro, E.; Lemire, J.A.; Demeter, M.; Vallini, G.; Turner, R.J.; Lampis, S. Antimicrobial Activity of Biogenically Produced Spherical Se-Nanomaterials Embedded in Organic Material against Pseudomonas aeruginosa and Staphylococcus aureus Strains on Hydroxyapatite-Coated Surfaces. Microb. Biotechnol. 2017, 10, 804–818. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Webster, T.J. Nanostructured Selenium for Preventing Biofilm Formation on Polycarbonate Medical Devices. J. Biomed. Mater. Res. A 2012, 100A, 3205–3210. [Google Scholar] [CrossRef]
- Yan, J.-K.; Qiu, W.-Y.; Wang, Y.-Y.; Wang, W.-H.; Yang, Y.; Zhang, H.-N. Fabrication and Stabilization of Biocompatible Selenium Nanoparticles by Carboxylic Curdlans with Various Molecular Properties. Carbohydr. Polym. 2018, 179, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wong, K.H.; Wu, H.L.; Zheng, W.; Wong, M.K.; Man, W.Y.C. Pleurotus Tuber-Regium Polysaccharide Functionalized Nano-Selenium Hydrosol with Anti-Tumor Activity and Preparation Method Thereof. U.S. Patent 9,072,669, 7 July 2015. [Google Scholar]
- Husen, A.; Siddiqi, K.S. Plants and Microbes Assisted Selenium Nanoparticles: Characterization and Application. J. Nanobiotechnology 2014, 12, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezhuthupurakkal, P.B.; Polaki, L.R.; Suyavaran, A.; Subastri, A.; Sujatha, V.; Thirunavukkarasu, C. Selenium Nanoparticles Synthesized in Aqueous Extract of Allium Sativum Perturbs the Structural Integrity of Calf Thymus DNA through Intercalation and Groove Binding. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 74, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Sowndarya, P.; Ramkumar, G.; Shivakumar, M.S. Green Synthesis of Selenium Nanoparticles Conjugated Clausena Dentata Plant Leaf Extract and Their Insecticidal Potential against Mosquito Vectors. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1490–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Wong, Y.-S.; Zheng, W.; Bai, Y.; Huang, L. Selenium Nanoparticles Fabricated in Undaria Pinnatifida Polysaccharide Solutions Induce Mitochondria-Mediated Apoptosis in A375 Human Melanoma Cells. Colloids Surf. B Biointerfaces 2008, 67, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.S.; Selvaraj, K. Biogenic Synthesis of Selenium Nanoparticles and Their Effect on As(III)-Induced Toxicity on Human Lymphocytes. Biol. Trace Elem. Res. 2014, 157, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Song, J.Y.; Kim, B.S. Rapid Biological Synthesis of Silver Nanoparticles Using Plant Leaf Extracts. Bioprocess Biosyst. Eng. 2009, 32, 79–84. [Google Scholar] [CrossRef]
- Suresh, K.; Prabagaran, S.R.; Sengupta, S.; Shivaji, S. Bacillus indicus Sp. Nov., an Arsenic-Resistant Bacterium Isolated from an Aquifer in West Bengal, India. Int. J. Syst. Evol. Microbiol. 2004, 54, 1369–1375. [Google Scholar] [CrossRef] [Green Version]
- Mandal, D.; Bolander, M.E.; Mukhopadhyay, D.; Sarkar, G.; Mukherjee, P. The Use of Microorganisms for the Formation of Metal Nanoparticles and Their Application. Appl. Microbiol. Biotechnol. 2006, 69, 485–492. [Google Scholar] [CrossRef]
- Wang, T.; Yang, L.; Zhang, B.; Liu, J. Extracellular Biosynthesis and Transformation of Selenium Nanoparticles and Application in H2O2 Biosensor. Colloids Surf. B Biointerfaces 2010, 80, 94–102. [Google Scholar] [CrossRef]
- Ingale, A.G. Biogenic Synthesis of Nanoparticles and Potential Applications: An Eco-Friendly Approach. J. Nanomed. Nanotechnol. 2013, 4. [Google Scholar] [CrossRef]
- Oremland, R.S.; Herbel, M.J.; Blum, J.S.; Langley, S.; Beveridge, T.J.; Ajayan, P.M.; Sutto, T.; Ellis, A.V.; Curran, S. Structural and Spectral Features of Selenium Nanospheres Produced by Se-Respiring Bacteria. Appl. Environ. Microbiol. 2004, 70, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kora, A.J.; Rastogi, L. Bacteriogenic Synthesis of Selenium Nanoparticles by Escherichia Coli ATCC 35218 and Its Structural Characterisation. IET Nanobiotechnol. 2017, 11, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Li, X.; Cheng, Y.; Xiao, X.; Lu, Z.; Wang, Y.; Wang, F. Aerobic Biogenesis of Selenium Nanoparticles by Enterobacter Cloacae Z0206 as a Consequence of Fumarate Reductase Mediated Selenite Reduction. Sci. Rep. 2017, 7, 3239. [Google Scholar] [CrossRef] [PubMed]
- Kora, A.J.; Rastogi, L. Biomimetic Synthesis of Selenium Nanoparticles by Pseudomonas Aeruginosa ATCC 27853: An Approach for Conversion of Selenite. J. Environ. Manag. 2016, 181, 231–236. [Google Scholar] [CrossRef]
- Fesharaki, P.J.; Nazari, P.; Shakibaie, M.; Rezaie, S.; Banoee, M.; Abdollahi, M.; Shahverdi, A.R. Biosynthesis of Selenium Nanoparticles Using Klebsiella Pneumoniae and Their Recovery by a Simple Sterilization Process. Braz. J. Microbiol. 2010, 41, 461–466. [Google Scholar] [CrossRef]
- Srivastava, N.; Mukhopadhyay, M. Biosynthesis and Structural Characterization of Selenium Nanoparticles Mediated by Zooglea Ramigera. Powder Technol. 2013, 244, 26–29. [Google Scholar] [CrossRef]
- Cavalu, S.; Prokisch, J.; Laslo, V.; Vicas, S. Preparation, Structural Characterisation and Release Study of Novel Hybrid Microspheres Entrapping Nanoselenium, Produced by Green Synthesis. IET Nanobiotechnol. 2017, 11, 426–432. [Google Scholar] [CrossRef]
- Tan, Y.; Yao, R.; Wang, R.; Wang, D.; Wang, G.; Zheng, S. Reduction of Selenite to Se(0) Nanoparticles by Filamentous Bacterium Streptomyces Sp. ES2-5 Isolated from a Selenium Mining Soil. Microb. Cell Factories 2016, 15, 157. [Google Scholar] [CrossRef] [Green Version]
- Zinicovscaia, I.; Chiriac, T.; Cepoi, L.; Rudi, L.; Culicov, O.; Frontasyeva, M.; Rudic, V. Selenium Uptake and Assessment of the Biochemical Changes in Arthrospira (Spirulina) Platensis Biomass during the Synthesis of Selenium Nanoparticles. Can. J. Microbiol. 2017, 63, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gharieb, M.M.; Wilkinson, S.C.; Gadd, G.M. Reduction of Selenium Oxyanions by Unicellular, Polymorphic and Filamentous Fungi: Cellular Location of Reduced Selenium and Implications for Tolerance. J. Ind. Microbiol. 1995, 14, 300–311. [Google Scholar] [CrossRef]
- Zhang, L.; Li, D.; Gao, P. Expulsion of Selenium/Protein Nanoparticles through Vesicle-like Structures by Saccharomyces Cerevisiae under Microaerophilic Environment. World J. Microbiol. Biotechnol. 2012, 28, 3381–3386. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.R.L.d.M.; Menolli, R.A.; Osaku, E.F.; Tramontina, R.; de Melo, R.H.; do Amaral, A.E.; Duarte, P.A.D.; de Carvalho, M.M.; Smiderle, F.R.; Silva, J.L. da C.; et al. Exopolysaccharides from Aspergillus Terreus: Production, Chemical Elucidation and Immunoactivity. Int. J. Biol. Macromol. 2019, 139, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Ortiz, E.J.; Gonzalez-Gil, G.; Saikaly, P.E.; van Hullebusch, E.D.; Lens, P.N.L. Effects of Selenium Oxyanions on the White-Rot Fungus Phanerochaete Chrysosporium. Appl. Microbiol. Biotechnol. 2015, 99, 2405–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetchinkina, E.; Loshchinina, E.; Kursky, V.; Nikitina, V. Reduction of Organic and Inorganic Selenium Compounds by the Edible Medicinal Basidiomycete Lentinula Edodes and the Accumulation of Elemental Selenium Nanoparticles in Its Mycelium. J. Microbiol. 2013, 51, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Pan, D.; Li, H.; Sun, Y.; Zeng, X.; Yan, B. Antioxidant and Immunomodulatory Activity of Selenium Exopolysaccharide Produced by Lactococcus Lactis Subsp. Lactis. Food Chem. 2013, 138, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Long, H.; Wang, C.; Zhu, Z.; Yu, L.; Yang, W.; Ren, X.; Liu, X. Characterization of Selenium-Containing Polysaccharide from Spirulina Platensis and Its Protective Role against Cd-Induced Toxicity. Int. J. Biol. Macromol. 2020, 164, 2465–2476. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zheng, W.; Wong, Y.-S.; Yang, F.; Bai, Y. Accumulation of Selenium in Mixotrophic Culture of Spirulina Platensis on Glucose. Bioresour. Technol. 2006, 97, 2260–2265. [Google Scholar] [CrossRef]
- Bannu, S.M.; Lomada, D.; Gulla, S.; Chandrasekhar, T.; Reddanna, P.; Reddy, M.C. Potential Therapeutic Applications of C-Phycocyanin. Curr. Drug Metab. 2020, 20, 967–976. [Google Scholar] [CrossRef]
- Hao, S.; Yan, Y.; Li, S.; Zhao, L.; Zhang, C.; Liu, L.; Wang, C. The In Vitro Anti-Tumor Activity of Phycocyanin against Non-Small Cell Lung Cancer Cells. Mar. Drugs 2018, 16, 178. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Ling, Q.; Cai, Z.; Wang, Y.; Zhang, Y.; Hoffmann, P.R.; Zheng, W.; Zhou, T.; Huang, Z. Selenium-Containing Phycocyanin from Se-Enriched Spirulina platensis Reduces Inflammation in Dextran Sulfate Sodium-Induced Colitis by Inhibiting NF-ΚB Activation. J. Agric. Food Chem. 2016, 64, 5060–5070. [Google Scholar] [CrossRef]
- Ji, Y.-B.; Dong, F.; Yu, M.; Qin, L.; Liu, D. Optimization of Synthesis of Seleno-Sargassum fusiforme (Harv.) Setch. Polysaccharide by Response Surface Methodology, Its Characterization, and Antioxidant Activity. J. Chem. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zhong, Y.; Luo, H.; Yang, Y. Selenium-Containing Polysaccharide-Protein Complex in Se-Enriched Ulva Fasciata Induces Mitochondria-Mediated Apoptosis in A549 Human Lung Cancer Cells. Mar. Drugs 2017, 15, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araie, H.; Shiraiwa, Y. Selenium Utilization Strategy by Microalgae. Molecules 2009, 14, 4880–4891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, D.; Li, Y.; Wang, C.; Wang, X.; Yu, Z.; Fu, X. A Novel Polysaccharide from Se-Enriched Ganoderma Lucidum Induces Apoptosis of Human Breast Cancer Cells. Oncol. Rep. 2011, 25, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Li, X. Preparation, physicochemical property and in vitro antioxidant activity of zinc-Hohenbuehelia serotina polysaccharides complex. Int. J. Biol. Macromol. 2019, 121, 862–869. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, C.; Wang, J.; Kumar, V.; Anwar, F.; Xiao, F.; Mushtaq, G.; Liu, Y.; Kamal, M.A.; Yuan, D. Inhibition on the Growth of Human MDA-MB-231 Breast Cancer Cells in Vitro and Tumor Growth in a Mouse Xenograft Model by Se-Containing Polysaccharides from Pyracantha Fortuneana. Nutr. Res. 2016, 36, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Liu, G.; Wang, M.; Lv, Z.; Du, P. A Selenium Polysaccharide from Platycodon Grandiflorum Rescues PC12 Cell Death Caused by H2O2 via Inhibiting Oxidative Stress. Int. J. Biol. Macromol. 2017, 104, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Chen, L.; Yang, Q.; Wang, Y.; Wei, X. Antitumor Activity of Se-Containing Tea Polysaccharides against Sarcoma 180 and Comparison with Regular Tea Polysaccharides and Se-Yeast. Int. J. Biol. Macromol. 2018, 120, 853–858. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Górska, S.; Maksymiuk, A.; Turło, J. Selenium-Containing Polysaccharides—Structural Diversity, Biosynthesis, Chemical Modifications and Biological Activity. Appl. Sci. 2021, 11, 3717. https://0-doi-org.brum.beds.ac.uk/10.3390/app11083717
Górska S, Maksymiuk A, Turło J. Selenium-Containing Polysaccharides—Structural Diversity, Biosynthesis, Chemical Modifications and Biological Activity. Applied Sciences. 2021; 11(8):3717. https://0-doi-org.brum.beds.ac.uk/10.3390/app11083717
Chicago/Turabian StyleGórska, Sandra, Anna Maksymiuk, and Jadwiga Turło. 2021. "Selenium-Containing Polysaccharides—Structural Diversity, Biosynthesis, Chemical Modifications and Biological Activity" Applied Sciences 11, no. 8: 3717. https://0-doi-org.brum.beds.ac.uk/10.3390/app11083717