
Assessment of the Genetic and Phytochemical Variability of Italian Wild Hop: A Route to Biodiversity Preservation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Genetic Analysis
2.2.1. SSR Analysis
2.2.2. Genetic Data Analysis
2.3. Phytochemical Characterization
2.3.1. Chemicals and Solvents
2.3.2. Bitter Acids Extraction
HPLC-UV Analysis
2.3.3. Essential Oil Extraction and GC/MS Analysis
2.4. Statistical Analysis
3. Results and Discussion
3.1. Genetic Characterization
3.1.1. Microsatellite Markers Polymorphism
3.1.2. Cluster Analysis
3.2. Phytochemical Characterization
3.2.1. Bitter Acids Quantification
3.2.2. Essential Oil Yields
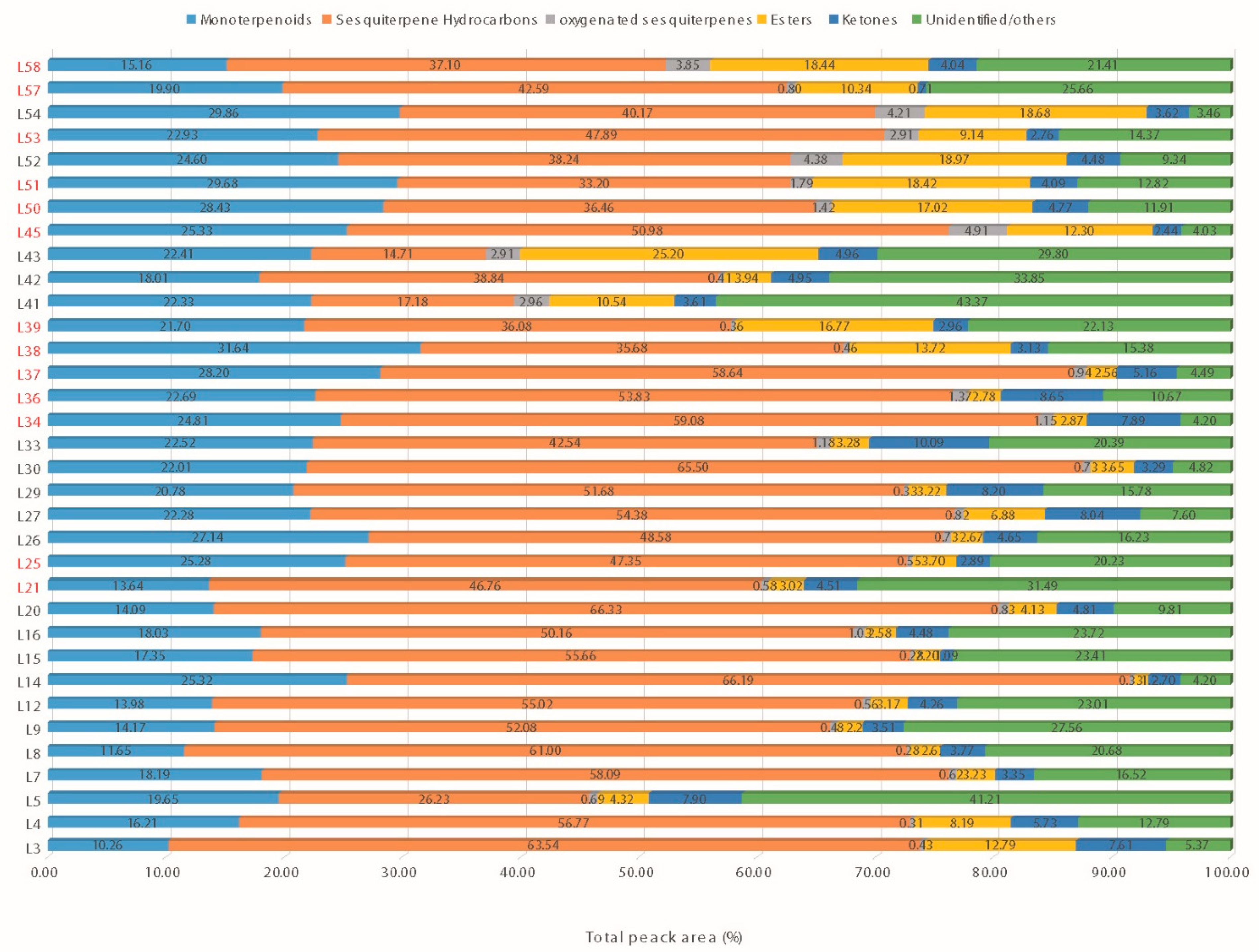
3.2.3. GC-MS Analysis-Essential Oil Profile
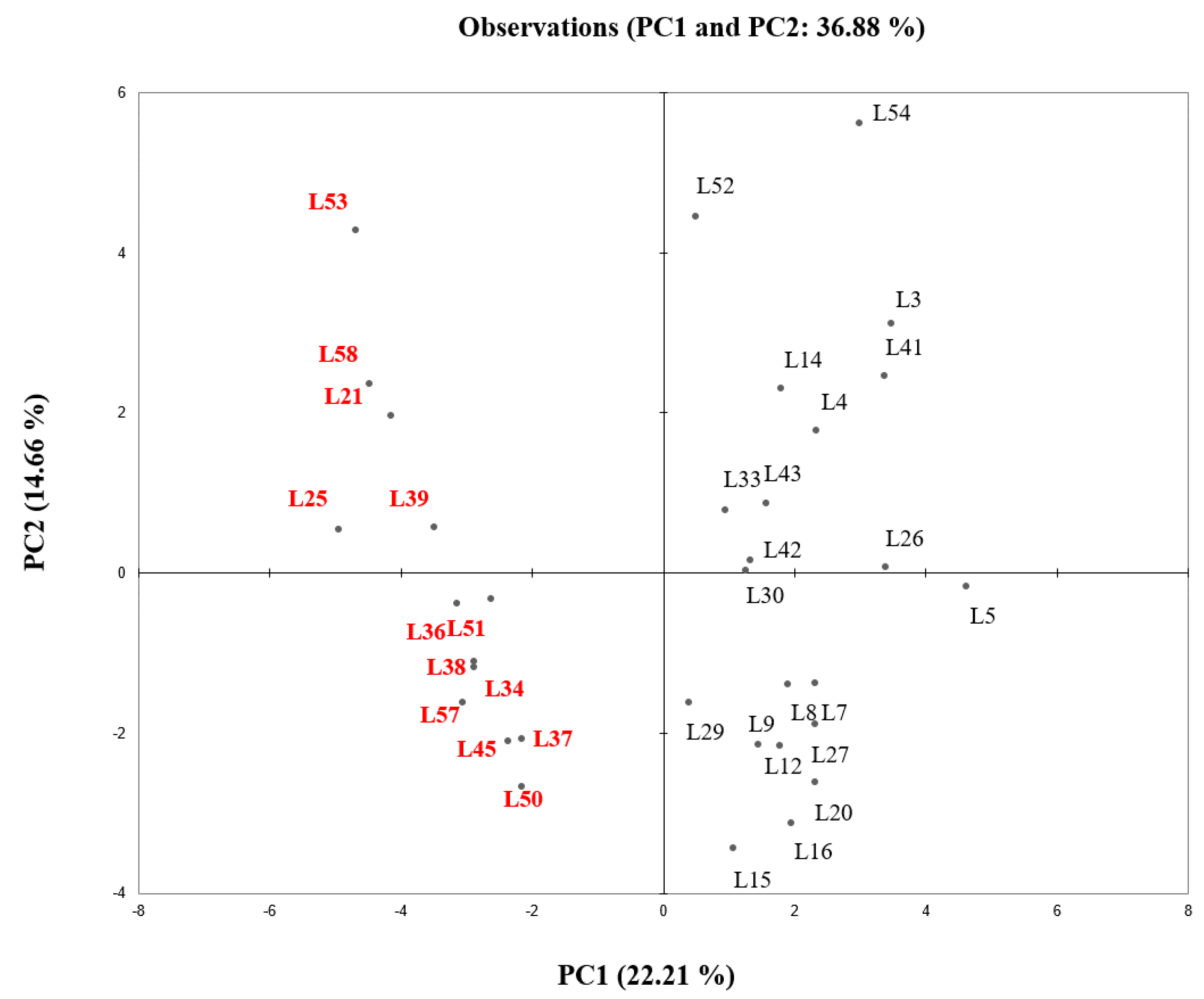
3.2.4. PCA Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson RJ, H.; Roberts, T.R.; Smith, R.J.; Bradley, L.L.; Moir, M. The inherent foam stabilising and lacing properties of some minor, hop-derived constituents of beer. In European Brewery Convention Symposium Beer Foam Quality; Verlag Hans Carl Getränke-Fachverlag: Amsterdam, The Netherlands, 1998; pp. 188–207. [Google Scholar]
- Neve, R.A. Hops; Chapman and Hall: London, UK, 1991. [Google Scholar]
- Falco, F.L.; Feitelson, E.; Dayan, T. Spatial Scale Mismatches in the Eu Agri-Biodiversity Conservation Policy. The Case for a Shift to Landscape-Scale Design. Land 2021, 10, 846. [Google Scholar] [CrossRef]
- Rodolfi, M.; Silvanini, A.; Chiancone, B.; Marieschi, M.; Fabbri, A.; Bruni, R.; Ganino, T. Identification and Genetic Structure of Wild Italian Humulus lupulus L. and Comparison with European and American Hop Cultivars Using Nuclear Microsatellite Markers. Genet. Resour. Crop Evol. 2018, 65, 1405–1422. [Google Scholar] [CrossRef]
- Grdiša, M.; Šatović, Z.; Liber, Z.; Jakše, J.; Varga, F.; Erhatić, R.; Srečec, S. High Genetic Diversity and Low Population Differentiation in Wild Hop (Humulus lupulus L.) from Croatia. Appl. Sci. 2021, 11, 6484. [Google Scholar] [CrossRef]
- Patzak, J.; Nesvadba, V.; Krofta, K.; Henychova, A.; Marzoev, A.I.; Richards, K. Evaluation of Genetic Variability of Wild Hops (Humulus lupulus L.) in Canada and the Caucasus Region by Chemical and Molecular Methods. Genome 2010, 53, 545–557. [Google Scholar] [CrossRef]
- Bassil, N.V.; Gilmore, B.; Oliphant, J.M.; Hummer, K.E.; Henning, J.A. Genic SSRs for European and North American Hop (Humulus lupulus L.). Genet. Resour. Crop Evol. 2008, 55, 959–969. [Google Scholar] [CrossRef]
- Murakami, A.; Darby, P.; Javornik, B.; Pais, M.S.S.; Seigner, E.; Lutz, A.; Svoboda, P. Microsatellite DNA Analysis of Wild Hops, Humulus lupulus L. Genet. Resour. Crop Evol. 2006, 53, 1553–1562. [Google Scholar] [CrossRef]
- Hrnčič, M.K.; Španinger, E.; Košir, I.J.; Knez, Ž.; Bren, U. Hop Compounds: Extraction Techniques, Chemical Analyses, Antioxidative, Antimicrobial, and Anticarcinogenic Effects. Nutrients 2019, 11, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Keukeleire, D. Fundamentals of beer and hop chemistry. Quim. Nova 2000, 23, 108–112. [Google Scholar] [CrossRef]
- Van Cleemput, M.; Cattoor, K.; De Bosscher, K.; Haegeman, G.; De Keukeleire, D.; Heyerick, A. Hop (Humulus lupulus)-derived bitter acids as multipotent bioactive compounds. J. Nat. Prod. 2009, 72, 1220–1230. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, N.; Yang, A.; Huang, J.; Ren, X.; Xian, M.; Zou, H. Hop Bitter Acids: Resources, Biosynthesis, and Applications. Appl. Microbiol. Biotechnol. 2021, 105, 4343–4356. [Google Scholar] [CrossRef]
- Zanoli, P.; Zavatti, M. Pharmacognostic and Pharmacological Profile of Humulus lupulus L. J. Ethnopharmacol. 2008, 116, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Almaguer, C.; Schönberger, C.; Gastl, M.; Arendt, E.K.; Becker, T. Humulus lupulus—A Story That Begs to Be Told. A Review. J. Inst. Brew. 2014, 120, 289–314. [Google Scholar] [CrossRef]
- Sharpe, F.R.; Laws, D.R.J. The essential oil of hops a review. J. Inst. Brew 1981, 87, 96–107. [Google Scholar] [CrossRef]
- Steenackers, B.; De Cooman, L.; De Vos, D. Chemical Transformations of Characteristic Hop Secondary Metabolites in Relation to Beer Properties and the Brewing Process: A Review. Food Chem. 2015, 172, 742–756. [Google Scholar] [CrossRef] [PubMed]
- Eri, S.; Khoo, B.K.; Lech, J.; Hartman, T.G. Direct Thermal Desorption-Gas Chromatography and Gas Chromatography-Mass Spectrometry Profiling of Hop (Humulus lupulus L.) Essential Oils in Support of Varietal Characterization. J. Agric. Food Chem. 2000, 4, 1140–1149. [Google Scholar] [CrossRef]
- Nance, M.R.; Setzer, W.N. Volatile Components of Aroma Hops (Humulus lupulus L.) Commonly Used in Beer Brewing. J. Brew. Distill. 2011, 2, 16–22. [Google Scholar]
- Kishimoto, T.; Wanikawa, A.; Kono, K.; Shibata, K. Comparison of the odor-active compounds in unhopped beer and beers hopped with different hop varieties. J. Agric. Food Chem. 2006, 54, 8855–8861. [Google Scholar] [CrossRef] [PubMed]
- Dabbous-Wach, A.; Rodolfi, M.; Paolini, J.; Costa, J.; Ganino, T. Characterization of Wild Corsican Hops and Assessment of the Performances of German Hops in Corsican Environmental Conditions through a Multidisciplinary Approach. Appl. Sci. 2021, 11, 3756. [Google Scholar] [CrossRef]
- Mongelli, A.; Rodolfi, M.; Ganino, T.; Marieschi, M.; Dall’Asta, C.; Bruni, R. Italian Hop Germplasm: Characterization of Wild Humulus lupulus L. Genotypes from Northern Italy by Means of Phytochemical, Morphological Traits and Multivariate Data Analysis. Ind. Crops Prod. 2015, 70, 16–27. [Google Scholar] [CrossRef]
- Kavalier, A.R.; Litt, A.; Ma, C.; Pitra, N.J.; Coles, M.C.; Kennelly, E.J.; Matthews, P.D. Phytochemical and Morphological Characterization of Hop (Humulus lupulus L.) Cones over Five Developmental Stages Using High Performance Liquid Chromatography Coupled to Time-of-Flight Mass Spectrometry, Ultrahigh Performance Liquid Chromatography Photodi. J. Agric. Food Chem. 2011, 59, 4783–4793. [Google Scholar] [CrossRef]
- McCallum, J.L.; Nabuurs, M.H.; Gallant, S.T.; Kirby, C.W.; Mills, A.A.S. Phytochemical Characterization of Wild Hops (Humulus lupulus Ssp. Lupuloides) Germplasm Resources From the Maritimes Region of Canada. Front. Plant Sci. 2019, 10, 1438. [Google Scholar] [CrossRef]
- Yan, D.D.; Wong, Y.F.; Shellie, R.A.; Marriott, P.J.; Whittock, S.P.; Koutoulis, A. Assessment of the Phytochemical Profiles of Novel Hop (Humulus lupulus L.) Cultivars: A Potential Route to Beer Crafting. Food Chem. 2019, 275, 15–23. [Google Scholar] [CrossRef]
- Doyle, J.; Doyle, J. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Stajner, N.; Jakse, J.; Kozjak, P.; Javornik, B. The Isolation and Characterisation of Microsatellites in Hop (Humulus lupulus L.). Plant Sci. 2005, 168, 213–221. [Google Scholar] [CrossRef]
- Kirby, L.T. Probability and Statistical Analysis. In DNA Fingerprinting; Springer: Berlin/Heidelberg, Germany, 1990; pp. 149–177. [Google Scholar] [CrossRef]
- Paetkau, D.; Calvert, W.; Stirling, I.; Strobeck, C. Microsatellite analysis of population structure in Canadian polar bears. Mol. Ecol. 1995, 4, 347–354. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314. [Google Scholar]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising How the Computer Program Cervus Accommodates Genotyping Error Increases Success in Paternity Assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Ganino, T.; Fabbri, A.; Rotondi, A.; Beghè, D. Genetic Resources of Olea europaea L. in the Bologna Province (Italy): SSR Analysis and Identification of Local Germplasm; Firenze University Press: Firenze, Italy, 2008; pp. 1000–1007. [Google Scholar] [CrossRef]
- Mongelli, A.; Rodolfi, M.; Ganino, T.; Marieschi, M.; Caligiani, A.; Dall’Asta, C.; Bruni, R. Are Humulus lupulus L. Ecotypes and Cultivars Suitable for the Cultivation of Aromatic Hop in Italy? A Phytochemical Approach. Ind. Crops Prod. 2016, 83, 693–700. [Google Scholar] [CrossRef]
- Rodolfi, M.; Chiancone, B.; Liberatore, C.M.; Fabbri, A.; Cirlini, M.; Ganino, T. Changes in Chemical Profile of Cascade Hop Cones According to the Growing Area. J. Sci. Food Agric. 2019, 99, 6011–6019. [Google Scholar] [CrossRef]
- Mafakheri, M.; Hamidoghli, Y. Micropropagation of Hop (Humulus lupulus L.) via Shoot Tip and Node Culture. Acta Hortic. 2019, 1236, 31–35. [Google Scholar] [CrossRef]
- Jakse, J.; Satovic, Z.; Javornik, B. Microsatellite Variability among Wild and Cultivated Hops (Humulus lupulus L.). Genome 2004, 47, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, C.; Belfiori, B.; Sileoni, V.; Marconi, O.; Perretti, G.; Bellucci, M.; Rubini, A. High Genetic and Chemical Diversity of Wild Hop Populations from Central Italy with Signals of a Genetic Structure Influenced by Both Sexual and Asexual Reproduction. Plant Sci. 2021, 304, 110794. [Google Scholar] [CrossRef]
- Patzak, J.; Nesvadba, V.; Henychová, A.; Krofta, K. Assessment of the Genetic Diversity of Wild Hops (Humulus lupulus L.) in Europe Using Chemical and Molecular Analyses. Biochem. Syst. Ecol. 2010, 38, 136–145. [Google Scholar] [CrossRef]
- Kondi’ckondi’c, D.; Andrejačerenak, A.A.; Košir, I.J.; Ocvirk, M.; Krmpot, T.; Ðurić, G.Ð. Morphological and Biochemical Characterization of Wild Hop (Humulus lupulus L.) Populations from Banja Luka Area (Bosnia and Herzegovina). Agronomy 2021, 11, 239. [Google Scholar] [CrossRef]
- Krofta, K. Comparison of Quality Parameters of Czech and Foreign Hop Varieties. Citeseer 2003, 49, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Van Opstaele, F.; Praet, T.; Aerts, G.; De Cooman, L. Characterization of Novel Single-Variety Oxygenated Sesquiterpenoid Hop Oil Fractions via Headspace Solid-Phase Microextraction and Gas Chromatography-Mass Spectrometry/Olfactometry. J. Agric. Food Chem. 2013, 61, 10555–10564. [Google Scholar] [CrossRef] [PubMed]
- Praet, T.; Van Opstaele, F.; Steenackers, B.; De Vos, D.; Aerts, G.; De Cooman, L. Flavor Activity of Sesquiterpene Oxidation Products, Formed upon Lab-Scale Boiling of a Hop Essential Oil-Derived Sesquiterpene Hydrocarbon Fraction (Cv. Saaz). J. Am. Soc. Brew. Chem. 2016, 74, 65–76. [Google Scholar] [CrossRef]
- Eyres, G.T.; Marriott, P.J.; Dufour, J.P. Comparison of Odor-Active Compounds in the Spicy Fraction of Hop (Humulus lupulus L.) Essential Oil from Four Different Varieties. J. Agric. Food Chem. 2007, 55. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| ID | Name/Location | Region | ID | Name/Location | Region |
|---|---|---|---|---|---|
| L1 | Adriano | Lombardy (IT) | L31 | Lodi Fico 1 | Lombardy (IT) |
| L2 | Arcore 1 | Lombardy (IT) | L32 | Lodi Fico 2 | Lombardy (IT) |
| L3 | Arcore 2 | Lombardy (IT) | L33 | Maddalena | Lombardy (IT) |
| L4 | Arcore 3 | Lombardy (IT) | L34 | Marter | Trentino Alto Adige (IT) |
| L5 | Asnago | Lombardy (IT) | L35 | Martinengo | Lombardy (IT) |
| L6 | Bevera | Lombardy (IT) | L36 | Masi | Trentino Alto Adige (IT) |
| L7 | Bigli | Lombardy (IT) | L37 | Masi Vigo | Trentino Alto Adige (IT) |
| L8 | Bigli 2 | Lombardy (IT) | L38 | Mezzano | Trentino Alto Adige (IT) |
| L9 | Bogno | Lombardy (IT) | L39 | Molina di Fiemme | Trentino Alto Adige (IT) |
| L10 | Bovisio | Lombardy (IT) | L40 | Mortara | Lombardy (IT) |
| L11 | Canardo | Lombardy (IT) | L41 | Mozzanico | Lombardy (IT) |
| L12 | Cantù | Lombardy (IT) | L42 | Oca | Lombardy (IT) |
| L13 | Casalnovo | Lombardy (IT) | L43 | Oggiono | Lombardy (IT) |
| L14 | Casalzunigo | Lombardy (IT) | L44 | Paradiso | Lombardy (IT) |
| L15 | Cascina Padregnano 1 | Lombardy (IT) | L45 | Ponte di Portolo | Trentino Alto Adige (IT) |
| L16 | Cascina Padregnano 2 | Lombardy (IT) | L46 | Pusiano | Lombardy (IT) |
| L17 | Cascina Padregnano 2b | Lombardy (IT) | L47 | Raimondi | Lombardy (IT) |
| L18 | Cascina V. | Lombardy (IT) | L48 | Riviera | Lombardy (IT) |
| L19 | Centa 1 | Trentino Alto Adige (IT) | L49 | S. Antonino | Lombardy (IT) |
| L20 | Cocquo | Lombardy (IT) | L50 | Santa Lucia | Trentino Alto Adige (IT) |
| L21 | Doiano | Trentino Alto Adige (IT) | L51 | Soravana | Trentino Alto Adige (IT) |
| L22 | Ello | Lombardy (IT) | L52 | Ticino di M. | Lombardy (IT) |
| L23 | Erba 1 | Lombardy (IT) | L53 | Transacqua | Trentino Alto Adige (IT) |
| L24 | Erba 2 | Lombardy (IT) | L54 | Trecate | Lombardy (IT) |
| L25 | Fai | Trentino Alto Adige (IT) | L55 | Trucazzano 1 | Lombardy (IT) |
| L26 | Gola Secca | Lombardy (IT) | L56 | Trucazzano 2 | Lombardy (IT) |
| L27 | Gola Secca 2 | Lombardy (IT) | L57 | Val di Non | Trentino Alto Adige (IT) |
| L28 | Gorgonzola | Lombardy (IT) | L58 | Vervo | Trentino Alto Adige (IT) |
| L29 | Ispra | Lombardy (IT) | L59 | Vigevano | Lombardy (IT) |
| L30 | Latur | Lombardy (IT) | L60 | Voghera | Lombardy (IT) |
| Primer | For 5′→3′ | Rev 5′→3′ | Repeat | Size (bp) | T (°C) |
|---|---|---|---|---|---|
| HlGA31 | CY5-CAAACTTGGTGCTCTAAGATGAA | CGTTTTCCCAACACCTAGTTC | (GA)17 | 163 | 55 |
| HlGT14 | CY5-GGCATGGCTAACTCTATATGC | AAATAGAAGTGCCATAACTGA | (TA)3A2(TA)3CATGC(GT)12 | 165 | 54 |
| HlGT16 | CY5-CCGTGATACAAATCTACCCAAA | CTCCAGTCAGCAATCTCTTCAA | (AC)21(AT)8 | 228 | 54 |
| HlGT17 | CY5-GGTCCTTAGTCACTTGCCAAT | GACTGTTCGAAGCACAATCAA | (GT)15 | 182 | 54 |
| HlACA3 | CY5-CAAGTTGTTGGTTGATTTCACAT | CTCCTTCCTGTGTTCACCAC | (CAA)15 | 215 | 52 |
| HlAGA6 | CY5-GTTAGAATCTCGTTGGCAA | TCTGAAACTTCACTAATCATC | (GAA)15N3(GAA)2 | 192 | 55 |
| HlAGA7 | CY5-ACAAGCAGTAATGATGAGGA | TCCAAGTCTCTCAATTAGGAA | (CAT)3N3(CTT)9N3(CTT)3 | 180 | 54 |
| HlGA23 | CY5-AAGCACGAAAACTGACTTG | GTTGCCCAAAATCACTGTT | (CT)24 | 245 | 54 |
| Locus | Locus | Locus | Locus | Locus | Locus | Locus | Locus | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HlAGA7 | f | HlGT16 | f | HlGT17 | f | HlACA3 | f | HlAGA6 | f | HlGA23 | f | HlGA31 | f | HlGT14 | f | |
| a | 160 | 0.025 | 211 | 0.108 | 175 | 0.167 | 204 | 0.008 | 170 | 0.167 | 239 | 0.008 | 160 | 0.342 | 161 | 0.075 |
| b | 166 | 0.133 | 213 | 0.058 | 177 | 0.133 | 206 | 0.050 | 172 | 0.025 | 244 | 0.258 | 162 | 0.033 | 165 | 0.283 |
| c | 171 | 0.008 | 227 | 0.067 | 179 | 0.092 | 210 | 0.517 | 174 | 0.042 | 247 | 0.05 | 164 | 0.350 | 167 | 0.608 |
| d | 178 | 0.025 | 229 | 0.325 | 181 | 0.042 | 220 | 0.133 | 176 | 0.025 | 251 | 0.008 | 166 | 0.217 | 169 | 0.017 |
| e | 181 | 0.058 | 231 | 0.033 | 183 | 0.017 | 222 | 0.067 | 182 | 0.217 | 255 | 0.067 | 168 | 0.017 | 175 | 0.017 |
| f | 183 | 0.017 | 233 | 0.283 | 185 | 0.308 | 224 | 0.017 | 184 | 0.008 | 257 | 0.033 | 188 | 0.042 | ||
| g | 185 | 0.050 | 235 | 0.033 | 187 | 0.158 | 228 | 0.066 | 186 | 0.125 | 265 | 0.008 | ||||
| h | 187 | 0.558 | 237 | 0.075 | 189 | 0.033 | 230 | 0.042 | 188 | 0.217 | 267 | 0.033 | ||||
| j | 190 | 0.108 | 239 | 0.017 | 191 | 0.008 | 232 | 0.017 | 191 | 0.133 | 269 | 0.008 | ||||
| k | 199 | 0.017 | 193 | 0.017 | 235 | 0.042 | 193 | 0.008 | 271 | 0.008 | ||||||
| l | 195 | 0.025 | 238 | 0.042 | 199 | 0.025 | 273 | 0.008 | ||||||||
| m | 205 | 0.008 | 279 | 0.025 | ||||||||||||
| n | 281 | 0.017 | ||||||||||||||
| o | 283 | 0.050 | ||||||||||||||
| p | 285 | 0.017 | ||||||||||||||
| q | 287 | 0.067 | ||||||||||||||
| r | 289 | 0.058 | ||||||||||||||
| s | 293 | 0.025 | ||||||||||||||
| t | 297 | 0.017 | ||||||||||||||
| u | 299 | 0.058 | ||||||||||||||
| v | 301 | 0.008 | ||||||||||||||
| w | 303 | 0.067 | ||||||||||||||
| x | 305 | 0.025 | ||||||||||||||
| y | 307 | 0.008 | ||||||||||||||
| z | 309 | 0.008 | ||||||||||||||
| aa | 311 | 0.008 | ||||||||||||||
| ab | 313 | 0.017 | ||||||||||||||
| ac | 315 | 0.008 | ||||||||||||||
| ad | 317 | 0.008 | ||||||||||||||
| ae | 319 | 0.008 | ||||||||||||||
| af | 323 | 0.008 | ||||||||||||||
| N | 10 | 9 | 11 | 11 | 12 | 31 | 6 | 5 | ||||||||
| He | 0.661 | 0.793 | 0.829 | 0.705 | 0.850 | 0.910 | 0.717 | 0.550 | ||||||||
| Ho | 0.617 | 0.700 | 0.783 | 0.717 | 0.683 | 0.767 | 0.533 | 0.550 | ||||||||
| PIC | 0.634 | 0.759 | 0.8 | 0.68 | 0.824 | 0.897 | 0.658 | 0.482 | ||||||||
| ID | α-Acids % (DM) | β-Acids % (DM) | COH% in Tot α-Acids | Oil Yield % |
|---|---|---|---|---|
| L3 | 1.777 ± 0.032 f–k | 1.323 ± 0.001 f–i | 19.032 ± 0.493 d–h | 0.329 ± 0.030 d–i |
| L4 | 2.216 ± 0.273 d–j | 1.659 ± 0.139 d–i | 20.806 ± 0.371 d–h | 0.395 ± 0.142 b–i |
| L5 | 2.262 ± 0.332 d–j | 1.445 ± 0.215 f–i | 24.215 ± 0.102 b–e | 0.351 ± 0.001 d–i |
| L7 | 4.105 ± 0.082 a | 1.735 ± 0.048 d–i | 25.864 ± 0.451 a–d | 0.630 ± 0.051 a,b,c |
| L8 | 1.197 ± 0.006 h–l | 2.022 ± 0.035 c–i | 18.421 ± 0.267 e–h | 0.514 ± 0.093 a–f |
| L9 | 2.193 ± 0.058 d–j | 1.627 ± 0.022 d–i | 19.911 ± 0.048 d–h | 0.364 ± 0.032 c–i |
| L12 | 1.695 ± 0.097 f–k | 2.798 ± 0.295 c–i | 16.730 ± 0.923 g,h,i | 0.253 ± 0.004 f–i |
| L14 | 0.000 ± 0.000 l | 2.514 ± 0.352 c–i | 0.000 ± 0.000 k | 0.294 ± 0.141 e–i |
| L15 | 1.982 ± 0.489 e–k | 1.873 ± 0.470 d–i | 28.391 ± 0.319 a,b,c | 0.434 ± 0.053 a–i |
| L16 | 2.853 ± 0.109 a–g | 1.743 ± 0.073 d–i | 29.163 ± 0.221 a,b | 0.425 ± 0.035 b–i |
| L20 | 3.377 ± 0.614 a–e | 3.890 ± 0.576 b,c | 16.634 ± 0.492 g–j | 0.347 ± 0.004 d–i |
| L21 | 4.125 ± 0.969 a | 3.283 ± 0.573 b–f | 18.778 ± 0.482 e–h | 0.454 ± 0.061 a–i |
| L25 | 2.361 ± 0.008 d–j | 2.656 ± 0.060 c–i | 22.424 ± 1.157 b–g | 0.472 ± 0.052 a–h |
| L26 | 3.848 ± 0.194 a,b,c | 2.202 ± 0.095 c–i | 23.853 ± 0.104 b,f | 0.344 ± 0.076 d–i |
| L27 | 2.437 ± 0.016 c–i | 1.543 ± 0.172 e–i | 22.582 ± 0.477 b–g | 0.436 ± 0.067 a–i |
| L29 | 1.945 ± 0.153 e–k | 1.974 ± 0.790 c–i | 14.432 ± 0.896 h,i,j | 0.452 ± 0.002 a–i |
| L30 | 0.596 ± 0.059 k,l | 1.217 ± 0.073 h,i | 16.506 ± 0.597 g–j | 0.223 ± 0.104 g,h,i |
| L33 | 3.449 ± 0.071 a–d | 1.846 ± 0.003 d–i | 22.061 ± 0.249 c–g | 0.530 ± 0.033 a–e |
| L34 | 1.571 ± 0.779 g–k | 3.088 ± 0.794 b–h | 24.559 ± 8.922 b–e | 0.286 ± 0.020 e–i |
| L36 | 3.049 ± 0.470 a–f | 4.774 ± 0.597 a,b | 19.819 ± 1.517 d–h | 0.660 ± 0.120 a,b |
| L37 | 1.091 ± 0.083 i–l | 1.426 ± 0.121 f–i | 32.046 ± 1.665 a | 0.507 ± 0.079 a–f |
| L38 | 1.463 ± 0.756 j,k,l | 2.633 ± 1.781 c–i | 15.988 ± 0.175 g–j | 0.211 ± 0.032 h,i |
| L39 | 3.229 ± 0.031 a–e | 3.520 ± 0.021 b,c,d | 15.097 ± 1.174 h,i,j | 0.420 ± 0.003 b–i |
| L41 | 2.949 ± 0.040 a–g | 3.265 ± 0.039 b–g | 17.078 ± 0.035 f–i | 0.202 ± 0.002 i |
| L42 | 1.105 ± 0.074 i–l | 3.228 ± 0.349 b–g | 10.802 ± 0.163 i,j | 0.345 ± 0.066 d–i |
| L43 | 1.303 ± 0.216 h–l | 3.473 ± 0.455 b–e | 16.106 ± 0.013 g–j | 0.267 ± 0.065 e–i |
| L45 | 1.687 ± 0.345 f–k | 2.030 ± 0.519 c–i | 9.785 ± 0.215 j | 0.345 ± 0.059 d–i |
| L50 | 2.836 ± 0.096 a–g | 2.258 ± 0.078 c–i | 20.337 ± 1.059 d–h | 0.455 ± 0.078 a–i |
| L51 | 2.184 ± 0.501 d–j | 2.658 ± 0.563 c–i | 18.425 ± 0.875 e–h | 0.567 ± 0.034 a–d |
| L52 | 2.636 ± 0.309 b–h | 2.184 ± 0.178 c–i | 16.137 ± 0.225 g–j | 0.490 ± 0.007 a–g |
| L53 | 4.043 ± 0.285 a,b | 6.462 ± 0.355 a | 16.841 ± 0.220 g,h,i | 0.700 ± 0.106 a |
| L54 | 2.369 ± 0.088 d–j | 1.064 ± 0.009 i | 20.468 ± 0.264 d–h | 0.290 ± 0.002 e–i |
| L57 | 1.071 ± 0.011 i–l | 3.198 ± 0.084 b–g | 14.883 ± 0.337 h,i,j | 0.634 ± 0.056 a,b,c |
| L58 | 2.134 ± 0.467 d–j | 3.261 ± 0.641 b–g | 15.805 ± 0.753 g–j | 0.508 ± 0.054 a–f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodolfi, M.; Marieschi, M.; Chiancone, B.; Ganino, T. Assessment of the Genetic and Phytochemical Variability of Italian Wild Hop: A Route to Biodiversity Preservation. Appl. Sci. 2022, 12, 5751. https://0-doi-org.brum.beds.ac.uk/10.3390/app12115751
Rodolfi M, Marieschi M, Chiancone B, Ganino T. Assessment of the Genetic and Phytochemical Variability of Italian Wild Hop: A Route to Biodiversity Preservation. Applied Sciences. 2022; 12(11):5751. https://0-doi-org.brum.beds.ac.uk/10.3390/app12115751
Chicago/Turabian StyleRodolfi, Margherita, Matteo Marieschi, Benedetta Chiancone, and Tommaso Ganino. 2022. "Assessment of the Genetic and Phytochemical Variability of Italian Wild Hop: A Route to Biodiversity Preservation" Applied Sciences 12, no. 11: 5751. https://0-doi-org.brum.beds.ac.uk/10.3390/app12115751