A Journey to the Conformational Analysis of T-Cell Epitope Peptides Involved in Multiple Sclerosis

 and
and
Abstract
:
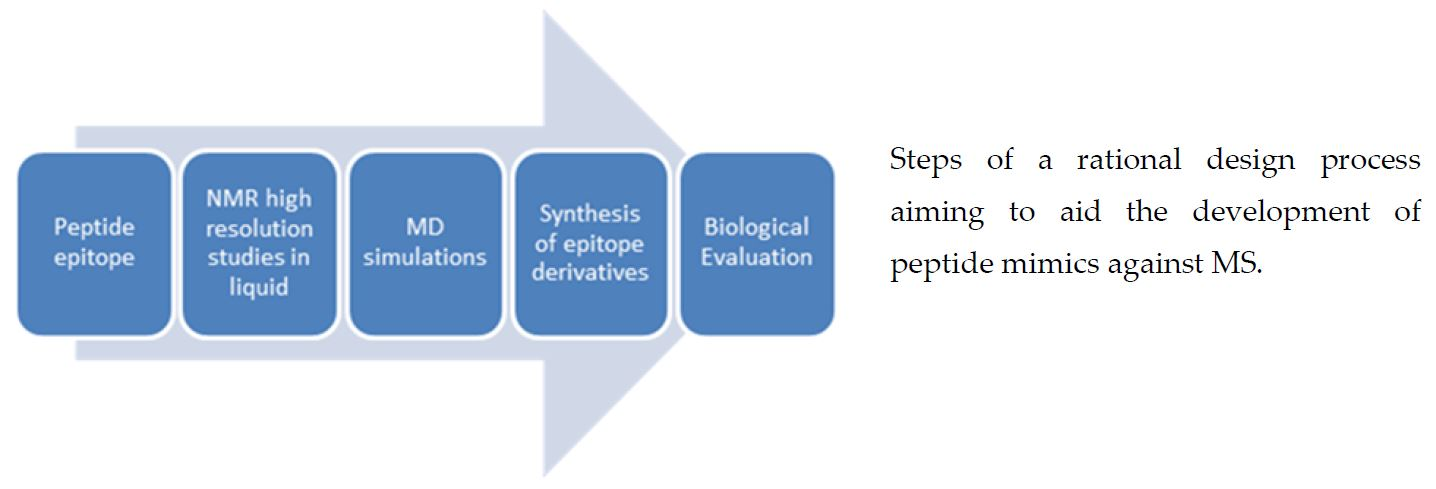{kind=link}
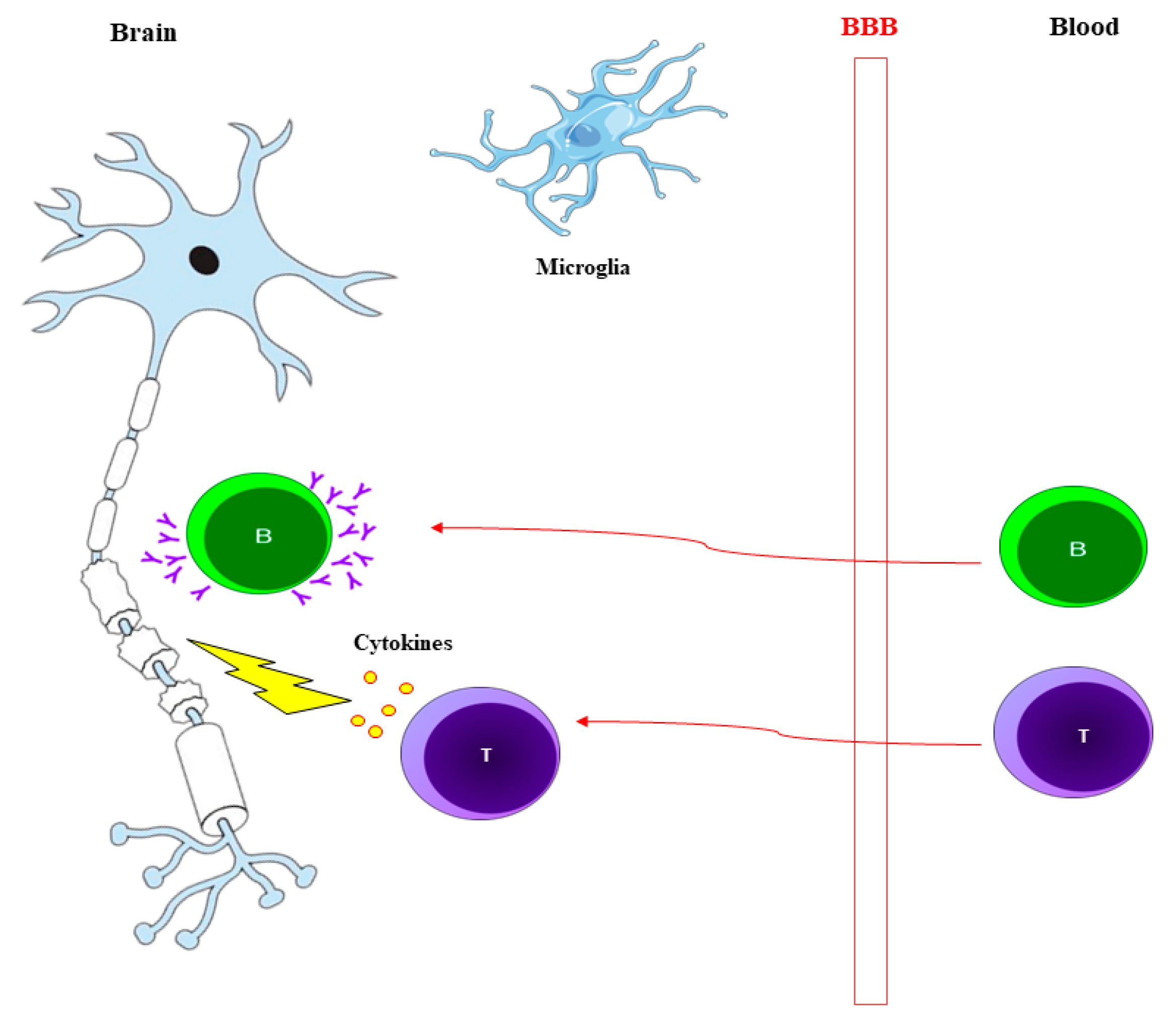{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
- Relapsing-remitting MS (RRMS): This is the most frequently occurring and affects ca. 85% of all MS patients. The patients with RRMS suffer from relapses and remissions of their neurological symptoms.
- Secondary progressive MS (SPMS): This follows the development of RRMS and causes further worsening of the disease.
- Primary progressive MS (PPMS): This affects 8–10% of patients and is characterized by the gradual further worsening of the disease.
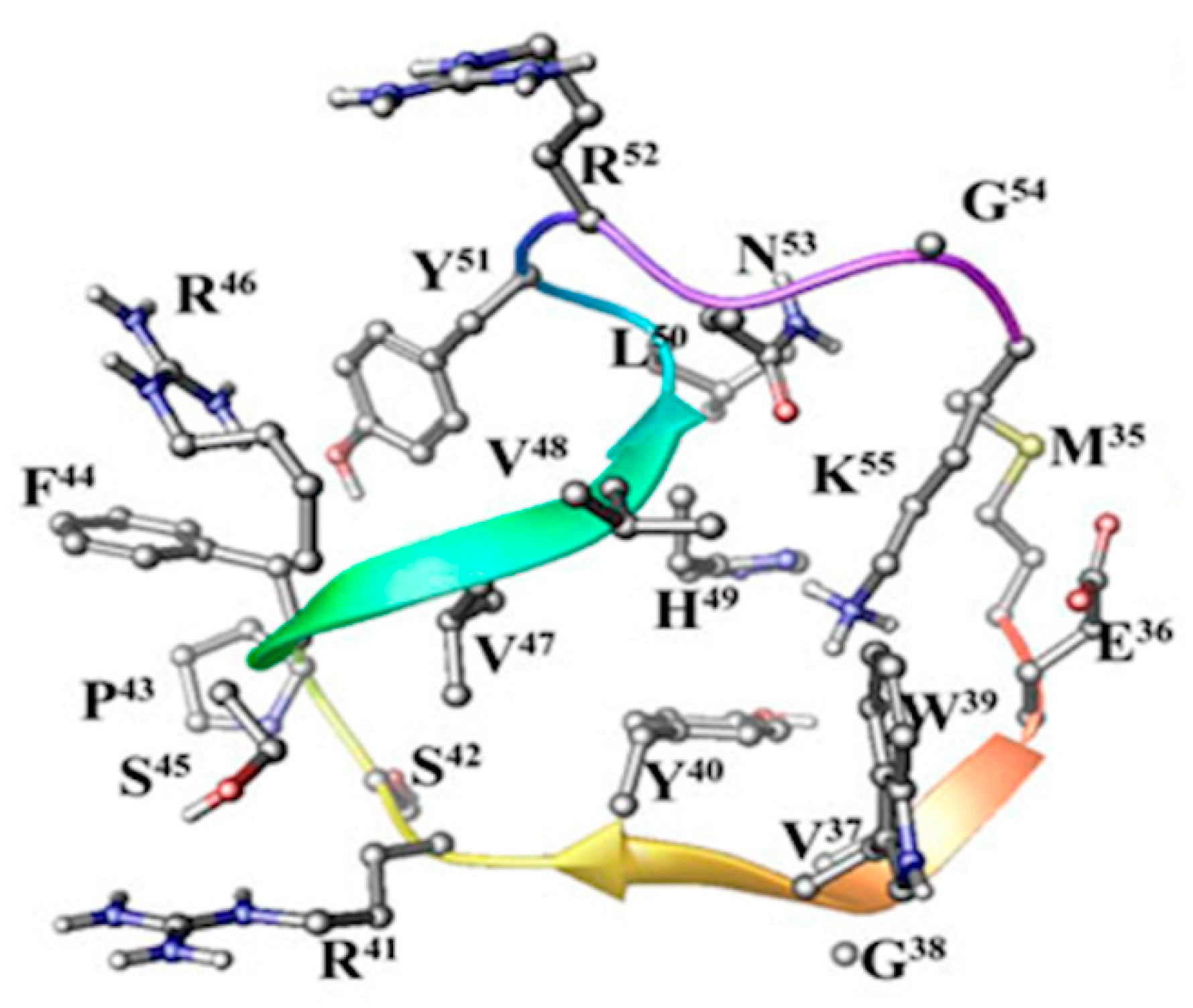
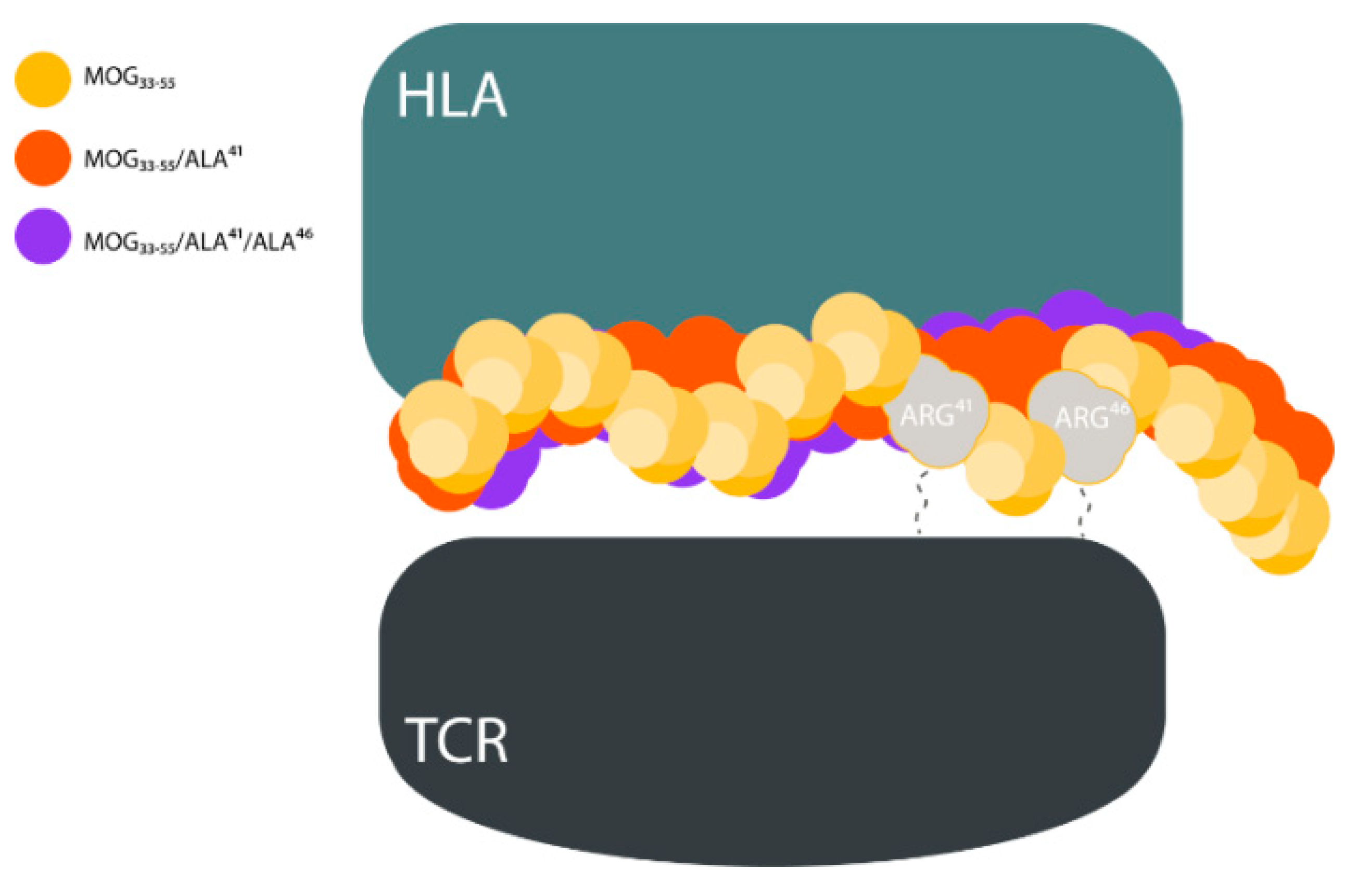
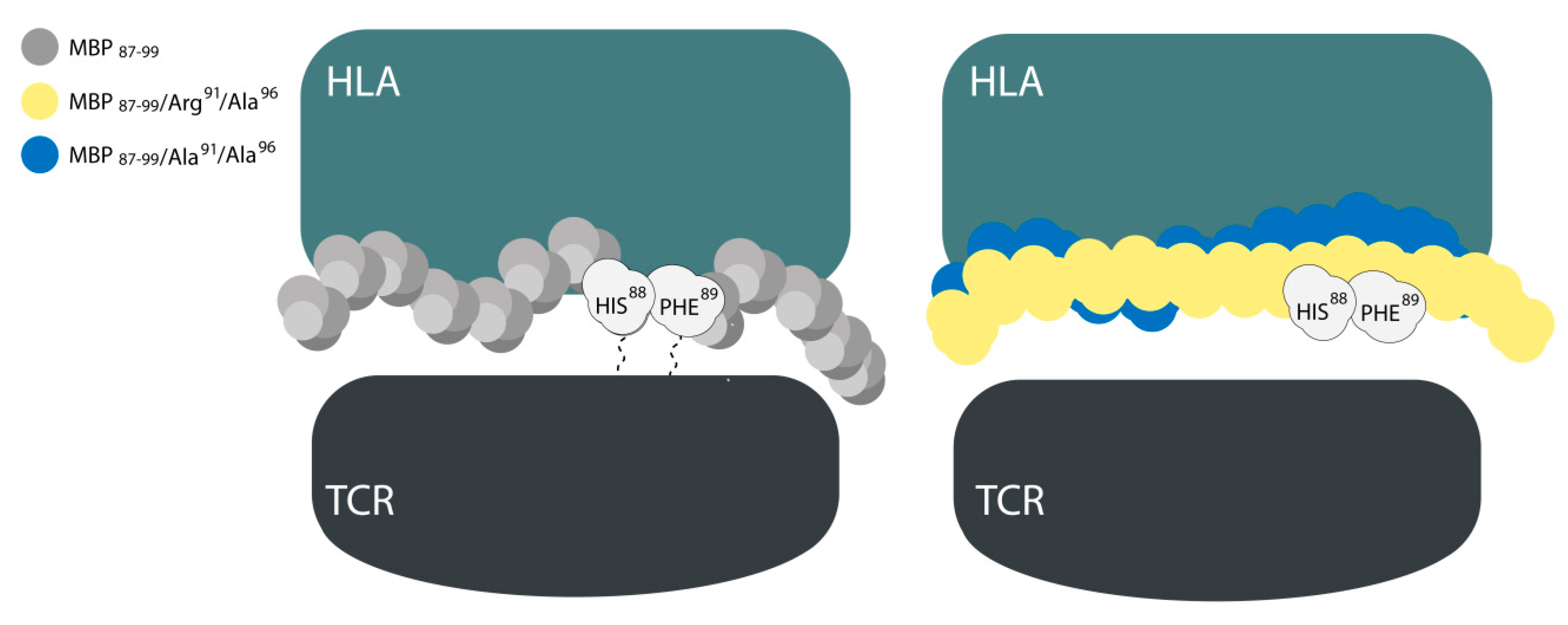
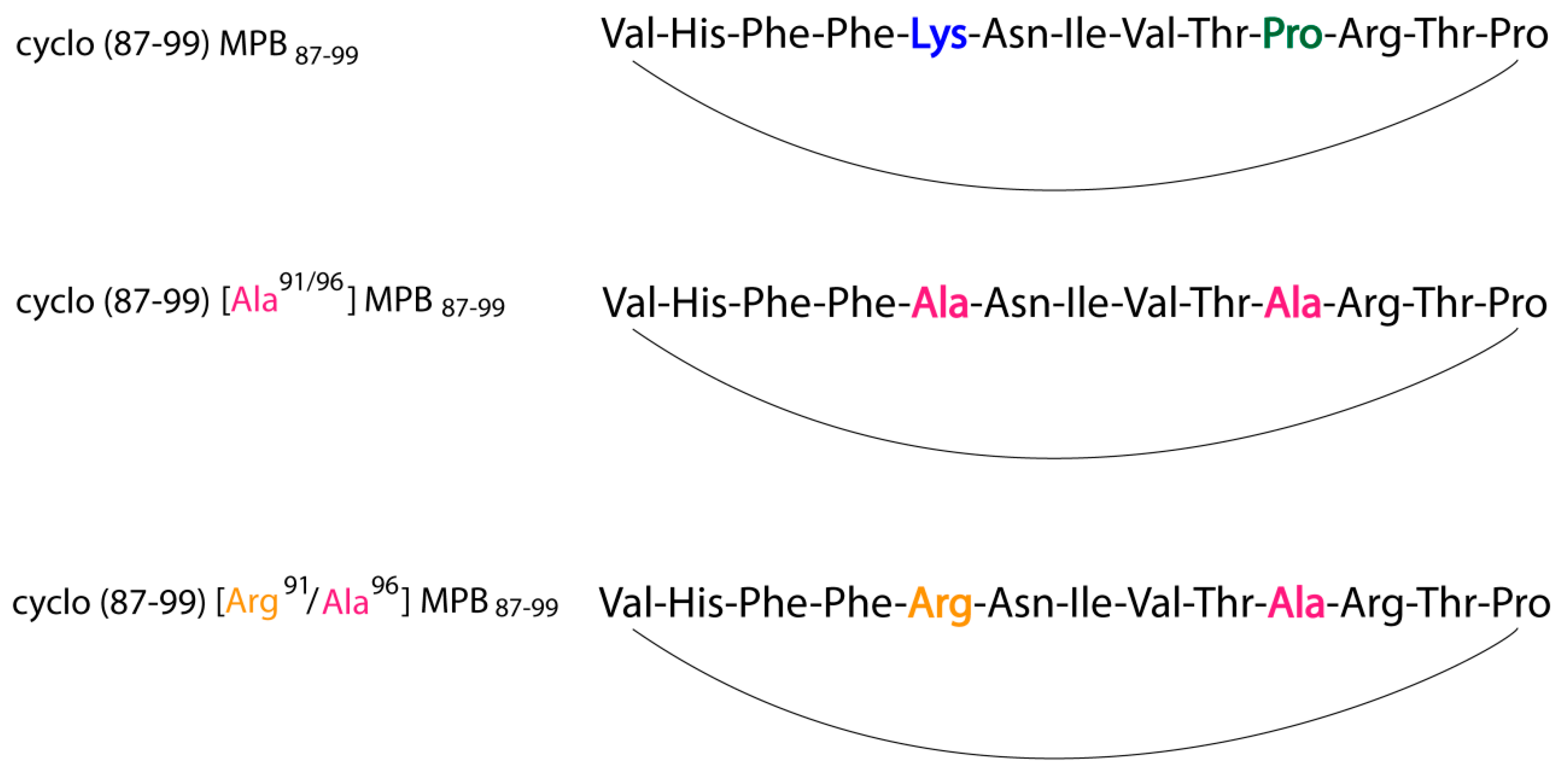
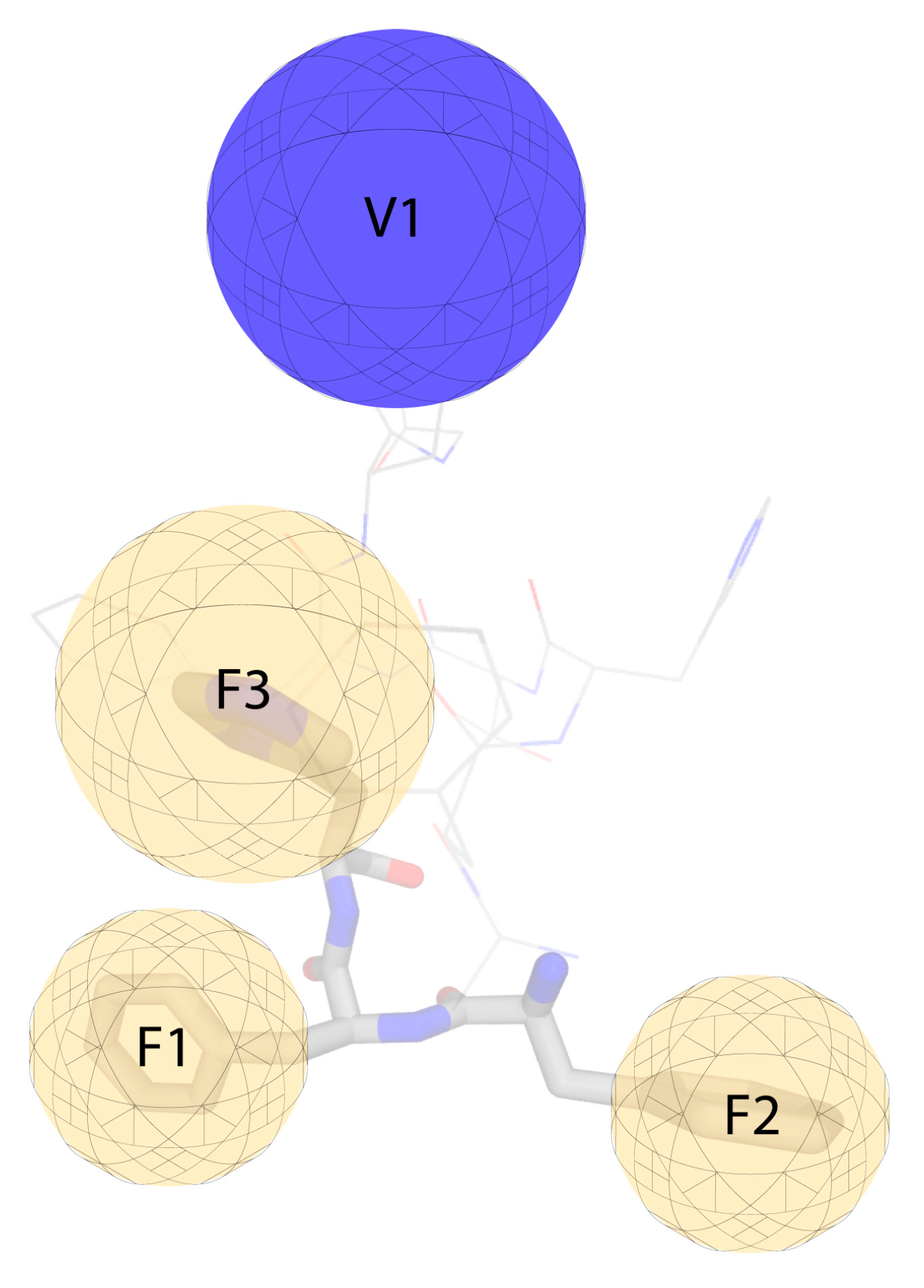
2. Results and Discussion
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wallin, M.T.; Culpepper, W.J.; Campbell, J.D.; Nelson, L.M.; Langer-Gould, A.; Marrie, R.A.; Cutter, G.R.; Kaye, W.E.; Wagner, L.; Tremlett, H.; et al. The prevalence of MS in the United States. Neurology 2019, 92, 1029–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, J.; Trevick, S.; Younger, D.S. Epidemiology of Multiple Sclerosis. Neurol. Clin. 2016, 34, 919–939. [Google Scholar] [CrossRef]
- Dziedzic, A.; Miller, E.; Saluk-Bijak, J.; Bijak, M. The gpr17 receptor—A promising goal for therapy and a potential marker of the neurodegenerative process in multiple sclerosis. Int. J. Mol. Sci. 2020, 21, 1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennum, P.; Wanscher, B.; Frederiksen, J.; Kjellberg, J. The socioeconomic consequences of multiple sclerosis: A controlled national study. Eur. Neuropsychopharmacol. 2012, 22, 36–43. [Google Scholar] [CrossRef]
- Raj, R.; Kaprio, J.; Korja, M.; Mikkonen, E.D.; Jousilahti, P.; Siironen, J. Risk of hospitalization with neurodegenerative disease after moderate-to-severe traumatic brain injury in the working-age population: A retrospective cohort study using the Finnish national health registries. PLoS Med. 2017, 14, e1002316. [Google Scholar] [CrossRef] [PubMed]
- Melchor, G.S.; Khan, T.; Reger, J.F.; Huang, J.K. Remyelination Pharmacotherapy Investigations Highlight Diverse Mechanisms Underlying Multiple Sclerosis Progression. ACS Pharmacol. Transl. Sci. 2019, 2, 372–386. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Amatya, B. Rehabilitation in Multiple Sclerosis: A Systematic Review of Systematic Reviews. Arch. Phys. Med. Rehabil. 2017, 98, 353–367. [Google Scholar] [CrossRef]
- Eckstein, C.; Bhatti, M.T. Currently approved and emerging oral therapies in multiple sclerosis: An update for the ophthalmologist. Surv. Ophthalmol. 2016, 61, 318–332. [Google Scholar] [CrossRef]
- Lublin, F.D.; Reingold, S.C. Defining the clinical course of multiple sclerosis: Results of an international survey. Neurology 1996, 46, 907–911. [Google Scholar] [CrossRef]
- Lublin, F.D.; Baier, M.; Cutter, G. Effect of relapses on development of residual deficit in multiple sclerosis. Neurology 2003, 61, 1528–1532. [Google Scholar] [CrossRef]
- Hemmer, B.; Archelos, J.J.; Hartung, H.P. New concepts in the immunopathogenesis of multiple sclerosis. Nat. Rev. Neurosci. 2002, 3, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Lassmann, H.; Raine, C.S.; Antel, J.; Prineas, J.W. Immunopathology of multiple sclerosis: Report on an international meeting held at the Institute of Neurology of the University of Vienna. J. Neuroimmunol. 1998, 213–217. [Google Scholar] [CrossRef]
- Tzakos, A.; Kursula, P.; Troganis, A.; Theodorou, V.; Tselios, T.; Svarnas, C.; Matsoukas, J.; Apostolopoulos, V.; Gerothanassis, I. Structure and Function of the Myelin Proteins: Current Status and Perspectives in Relation to Multiple Sclerosis. Curr. Med. Chem. 2005, 12, 1569–1587. [Google Scholar] [CrossRef] [PubMed]
- Kouskoff, V.; Lacaud, G.; Nemazee, D. T cell-independent rescue of B lymphocytes from peripheral immune tolerance. Science 2000, 287, 2501–2513. [Google Scholar] [CrossRef]
- Tuusa, J.; Raasakka, A.; Ruskamo, S.; Kursula, P. Myelin-derived and putative molecular mimic peptides share structural properties in aqueous and membrane-like environments. Mult. Scler. Demyelinating Disord. 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Bielekova, B.; Sung, M.-H.; Kadom, N.; Simon, R.; McFarland, H.; Martin, R. Expansion and Functional Relevance of High-Avidity Myelin-Specific CD4 + T Cells in Multiple Sclerosis. J. Immunol. 2004, 172, 3893–3904. [Google Scholar] [CrossRef] [Green Version]
- Greer, J.M.; Csurhes, P.A.; Cameron, K.D.; McCombe, P.A.; Good, M.F.; Pender, M.P. Increased immunoreactivity to two overlapping peptides of myelin proteolipid protein in multiple sclerosis. Brain 1997, 120, 1447–1460. [Google Scholar] [CrossRef]
- Mendel, I.; De Rosbo, N.K.; Ben-Nun, A. Delineation of the minimal encephalitogenic epitope within the immunodominant region of myelin oligodendrocyte glycoprotein: Diverse Vβ gene usage by T cells recognizing the core epitope encephalitogenic for T cell receptor Vβb and T cell receptor Vβa H-2. Eur. J. Immunol. 1996, 26, 2470–2479. [Google Scholar] [CrossRef]
- De Rosbo, N.K.; Kaye, J.F.; Eisenstein, M.; Mendel, I.; Hoeftberger, R.; Lassmann, H.; Milo, R.; Ben-Nun, A. The Myelin-Associated Oligodendrocytic Basic Protein Region MOBP15–36 Encompasses the Immunodominant Major Encephalitogenic Epitope(s) for SJL/J Mice and Predicted Epitope(s) for Multiple Sclerosis-Associated HLA-DRB1*1501. J. Immunol. 2004, 173, 1426–1435. [Google Scholar] [CrossRef] [Green Version]
- Holz, A.; Bielekova, B.; Martin, R.; Oldstone, M.B.A. Myelin-Associated Oligodendrocytic Basic Protein: Identification of an Encephalitogenic Epitope and Association with Multiple Sclerosis. J. Immunol. 2000, 164, 1103–1109. [Google Scholar] [CrossRef] [Green Version]
- Denic, A.; Johnson, A.J.; Bieber, A.J.; Warrington, A.E.; Rodriguez, M.; Pirko, I. The relevance of animal models in multiple sclerosis research. Pathophysiology 2011, 18, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gholamzad, M.; Ebtekar, M.; Ardestani, M.S.; Azimi, M.; Mahmodi, Z.; Mousavi, M.J.; Aslani, S. A comprehensive review on the treatment approaches of multiple sclerosis: Currently and in the future. Inflamm. Res. 2019, 68, 25–38. [Google Scholar] [CrossRef]
- Shahrizaila, N.; Yuki, N. Guillain-Barré syndrome animal model: The first proof of molecular mimicry in human autoimmune disorder. J. Biomed. Biotechnol. 2011, 2011, 829129. [Google Scholar] [CrossRef] [Green Version]
- Moise, L.; Beseme, S.; Tassone, R.; Liu, R.; Kibria, F.; Terry, F.; Martin, W.; De Groot, A.S. T cell epitope redundancy: Cross-conservation of the TCR face between pathogens and self and its implications for vaccines and autoimmunity. Expert Rev. Vaccines 2016, 15, 607–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsoukas, J.; Apostolopoulos, V.; Mavromoustakos, T. Designing Peptide Mimetics for the Treatment of Multiple Sclerosis. Mini Rev. Med. Chem. 2005, 1, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef]
- Davis, M.M.; Boniface, J.J.; Reich, Z.; Lyons, D.; Hampl, J.; Arden, B.; Chien, Y. Ligand Recognition By alpha beta T Cell Receptors. Annu. Rev. Immunol. 1998, 16, 523–544. [Google Scholar] [CrossRef] [PubMed]
- Wootla, B.; Eriguchi, M.; Rodriguez, M. Is multiple sclerosis an autoimmune disease? Autoimmune Dis. 2012, 2012, 969657. [Google Scholar] [CrossRef] [PubMed]
- Hare, B.J.; Wyss, D.F.; Osburne, M.S.; Kern, P.S.; Reinherz, E.L.; Wagner, G. Structure, specificity and CDR mobility of a class II restricted single- chain T-cell receptor. Nat. Struct. Biol. 1999, 6, 574–581. [Google Scholar] [CrossRef]
- Rudolph, M.G.; Wilson, I.A. The specificity of TCR/pMHC interaction. Curr. Opin. Immunol. 2002, 14, 52–65. [Google Scholar] [CrossRef]
- Rudolph, M.G.; Stanfield, R.L.; Wilson, I.A. How TCRS Bind MHCS, Peptides, And Coreceptors. Annu. Rev. Immunol. 2006, 24, 419–466. [Google Scholar] [CrossRef] [PubMed]
- Lassmann, H. Axonal and neuronal pathology in multiple sclerosis: What have we learnt from animal models. Exp. Neurol. 2010, 225, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Reuter, E.; Gollan, R.; Grohmann, N.; Paterka, M.; Salmon, H.; Birkenstock, J.; Richers, S.; Leuenberger, T.; Brandt, A.U.; Kuhlmann, T.; et al. Cross-Recognition of a myelin peptide by CD8+ T cells in the CNS is not sufficient to promote neuronal damage. J. Neurosci. 2015, 35, 4837–4850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madden, D.R. The Three-Dimensional Structure of Peptide-MHC Complexes. Annu. Rev. Immunol. 1995, 13, 587–622. [Google Scholar] [CrossRef]
- Adams, E.J.; Luoma, A.M. The Adaptable Major Histocompatibility Complex (MHC) Fold: Structure and Function of Nonclassical and MHC Class I–Like Molecules. Annu. Rev. Immunol. 2013, 31, 529–561. [Google Scholar] [CrossRef]
- Yannakakis, M.P.; Tzoupis, H.; Michailidou, E.; Mantzourani, E.; Simal, C.; Tselios, T. Molecular dynamics at the receptor level of immunodominant myelin oligodendrocyte glycoprotein 35–55 epitope implicated in multiple sclerosis. J. Mol. Graph. Model. 2016, 68, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Mouzaki, A.; Tselios, T.; Papathanassopoulos, P.; Matsoukas, I.; Chatzantoni, K. Immunotherapy for Multiple Sclerosis: Basic Insights for New Clinical Strategies. Curr. Neurovasc. Res. 2005, 1, 325–340. [Google Scholar] [CrossRef]
- Tselios, T.; Daliani, I.; Deraos, S.; Thymianou, S.; Matsoukas, E.; Troganis, A.; Gerothanassis, I.; Mouzaki, A.; Mavromoustakos, T.; Probert, L.; et al. Treatment of experimental allergic encephalomyelitis (EAE) by a rationally designed cyclic analogue of myelin basic protein (MBP) epitope 72-85. Bioorg. Med. Chem. Lett. 2000, 10, 2713–2717. [Google Scholar] [CrossRef]
- Tselios, T.; Apostolopoulos, V.; Daliani, I.; Deraos, S.; Grdadolnik, S.; Mavromoustakos, T.; Melachrinou, M.; Thymianou, S.; Probert, L.; Mouzaki, A.; et al. Antagonistic effects of human cyclic MBP87-99 altered peptide ligands in experimental allergic encephalomyelitis and human T-cell proliferation. J. Med. Chem. 2002, 45, 275–283. [Google Scholar] [CrossRef]
- Ntountaniotis, D.; Vanioti, Μ.; Kordopati, G.G.; Kellici, T.F.; Marousis, K.D.; Mavromoustakos, T.; Spyroulias, G.A.; Golic Grdadolnik, S.; Tselios, T.V. A combined NMR and molecular dynamics simulation study to determine the conformational properties of rat/mouse 35-55 myelin oligodendrocyte glycoprotein epitope implicated in the induction of experimental autoimmune encephalomyelitis. J. Biomol. Struct. Dyn. 2017, 35, 1559–1567. [Google Scholar] [CrossRef]
- Mantzourani, E.D.; Platts, J.A.; Brancale, A.; Mavromoustakos, T.M.; Tselios, T.V. Molecular dynamics at the receptor level of immunodominant myelin basic protein epitope 87-99 implicated in multiple sclerosis and its antagonists altered peptide ligands: Triggering of immune response. J. Mol. Graph. Model. 2007, 26, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Willcox, B.E.; Gao, G.F.; Wyer, J.R.; Ladbury, J.E.; Bell, J.I.; Jakobsen, B.K.; Van der Merwe, P.A. TCR binding to peptide-MHC stabilizes a flexible recognition interface. Immunity 1999, 10, 357–365. [Google Scholar] [CrossRef] [Green Version]
- Matsoukas, J.; Apostolopoulos, V.; Kalbacher, H.; Papini, A.M.; Tselios, T.; Chatzantoni, K.; Biagioli, T.; Lolli, F.; Deraos, S.; Papathanassopoulos, P.; et al. Design and synthesis of a novel potent myelin basic protein epitope 87-99 cyclic analogue: Enhanced stability and biological properties of mimics render them a potentially new class of immunomodulators. J. Med. Chem. 2005, 48, 1470–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emmanouil, M.; Tseveleki, V.; Triantafyllakou, I.; Nteli, A.; Tselios, T.; Probert, L. A cyclic altered peptide analogue based on myelin basic protein 87-99 provides lasting prophylactic and therapeutic protection against acute experimental autoimmune encephalomyelitis. Molecules 2018, 23, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spyranti, Z.; Dalkas, G.A.; Spyroulias, G.A.; Mantzourani, E.D.; Mavromoustakos, T.; Friligou, I.; Matsoukas, J.M.; Tselios, T.V. Putative bioactive conformations of amide linked cyclic myelin basic protein peptide analogues associated with experimental autoimmune encephalomyelitis. J. Med. Chem. 2007, 50, 6039–6047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deraos, G.; Chatzantoni, K.; Matsoukas, M.T.; Tselios, T.; Deraos, S.; Katsara, M.; Papathanasopoulos, P.; Vynios, D.; Apostolopoulos, V.; Mouzaki, A.; et al. Citrullination of linear and cyclic altered peptide ligands from myelin basic protein (MBP87-99) epitope elicits a Th1 polarized response by T cells isolated from multiple sclerosis patients: Implications in triggering disease. J. Med. Chem. 2008, 51, 7834–7842. [Google Scholar] [CrossRef] [PubMed]
- Matsoukas, J.; Apostolopoulos, V.; Lazoura, E.; Deraos, G.; Matsoukas, M.-T.; Katsara, M.; Tselios, T.; Deraos, S. Round and Round we Go: Cyclic Peptides in Disease. Curr. Med. Chem. 2006. [Google Scholar] [CrossRef]
- Tzakos, A.G.; Fuchs, P.; Van Nuland, N.A.J.; Troganis, A.; Tselios, T.; Deraos, S.; Matsoukas, J.; Gerothanassis, I.P.; Bonvin, A.M.J.J. NMR and molecular dynamics studies of an autoimmune myelin basic protein peptide and its antagonist: Structural implications for the MHC II (I-A u)-peptide complex from docking calculations. Eur. J. Biochem. 2004, 271, 3399–3413. [Google Scholar] [CrossRef]
- Wraith, D.C.; Smilek, D.E.; Mitchell, D.J.; Steinman, L.; McDevitt, H.O. Antigen recognition in autoimmune encephalomyelitis and the potential for peptide-mediated immunotherapy. Cell 1989, 59, 247–255. [Google Scholar] [CrossRef]
- Lee, C.; Liang, M.N.; Tate, K.M.; Rabinowitz, J.D.; Beeson, C.; Jones, P.P.; McConnell, H.M. Evidence that the autoimmune antigen myelin basic protein (MBP) Ac1-9 binds towards one end of the major histocompatibility complex (MHC) cleft. J. Exp. Med. 1998, 187, 1505–1516. [Google Scholar] [CrossRef] [Green Version]
- He, X.L.; Radu, C.; Sidney, J.; Sette, A.; Ward, E.S.; Garcia, K.C. Structural snapshot of aberrant antigen presentation linked to autoimmunity: The immunodominant epitope of MBP complexed with I-Au. Immunity 2002, 17, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Fugger, L.; Liang, J.; Gautam, A.; Rothbard, J.B.; McDevitt, H.O. Quantitative analysis of peptides from myelin basic protein binding to the MHC class II protein, I-A(u), which confers susceptibility to experimental allergic encephalomyelitis. Mol. Med. 1996, 2, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzler, B.; Wraith, D.C. Inhibition of experimental autoimmune encephalomyelitis by inhalation but not oral administration of the encephalitogenic peptide: Influence of MHC binding affinity. Int. Immunol. 1993, 5, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Laimou, D.; Lazoura, E.; Troganis, A.N.; Matsoukas, M.T.; Deraos, S.N.; Katsara, M.; Matsoukas, J.; Apostolopoulos, V.; Tselios, T.V. Conformational studies of immunodominant myelin basic protein 1-11 analogues using NMR and molecular modeling. J. Comput. Aided. Mol. Des. 2011, 25, 1019–1032. [Google Scholar] [CrossRef]
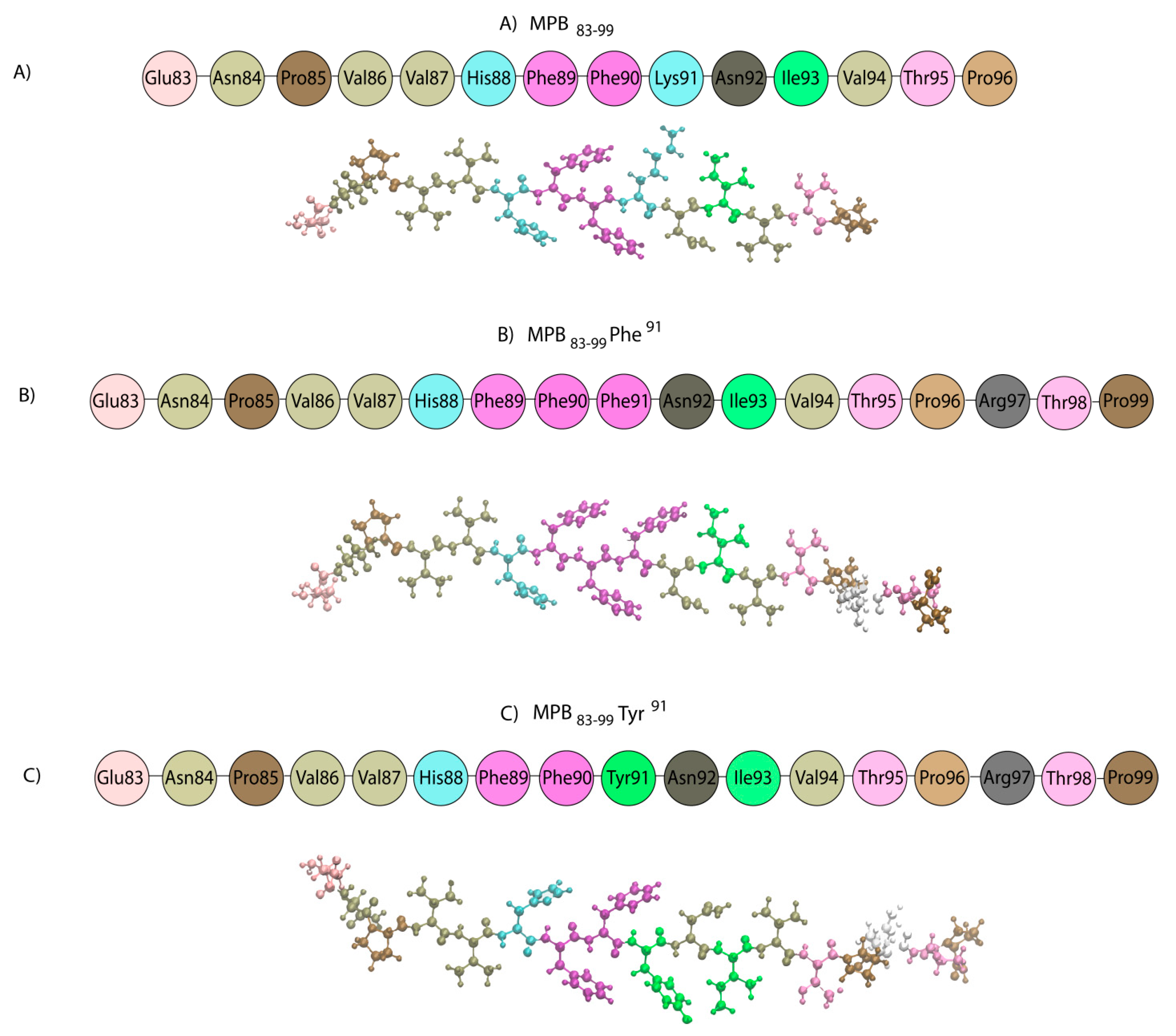
- Spyranti, Z.; Tselios, T.; Deraos, G.; Matsoukas, J.; Spyroulias, G.A. NMR structural elucidation of myelin basic protein epitope 83-99 implicated in multiple sclerosis. Amino Acids 2010, 38, 929–936. [Google Scholar] [CrossRef]
- Mantzourani, E.; Mavromoustakos, T.; Platts, J.; Matsoukas, J.; Tselios, T. Structural Requirements for Binding of Myelin Basic Protein (MBP) Peptides to MHC II: Effects on Immune Regulation. Curr. Med. Chem. 2005, 12, 1521–1535. [Google Scholar] [CrossRef]
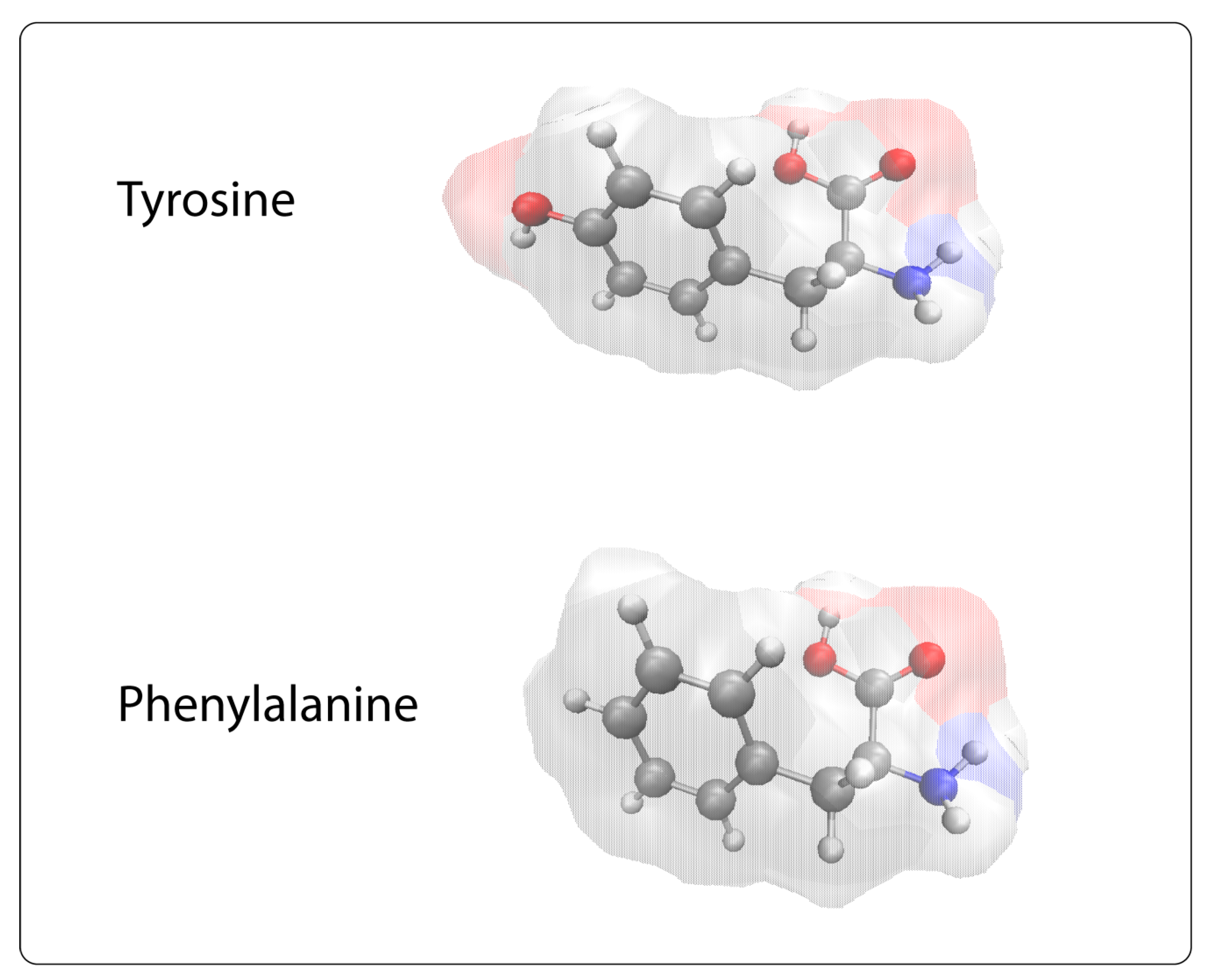
- Mantzourani, E.D.; Tselios, T.V.; Grdadolnik, S.G.; Platts, J.A.; Brancale, A.; Deraos, G.N.; Matsoukas, J.M.; Mavromoustakos, T.M. Comparison of proposed putative active conformations of myelin basic protein epitope 87-99 linear altered peptide ligands by spectroscopic and modelling studies: The role of positions 91 and 96 in T-cell receptor activation. J. Med. Chem. 2006, 49, 6683–6691. [Google Scholar] [CrossRef]
- Mantzourani, E.D.; Tselios, T.V.; Grdadolnik, S.G.; Brancale, A.; Platts, J.A.; Matsoukas, J.M.; Mavromoustakos, T.M. A putative bioactive conformation for the altered peptide ligand of myelin basic protein and inhibitor of experimental autoimmune encephalomyelitis [Arg91, Ala96] MBP87-99. J. Mol. Graph. Model. 2006, 25, 17–29. [Google Scholar] [CrossRef]
- Mantzourani, E.; Laimou, D.; Matsoukas, M.; Tselios, T. Peptides as Therapeutic Agents or Drug Leads for Autoimmune, Hormone Dependent and Cardiovascular Diseases. Antiinflamm. Antiallergy Agents Med. Chem. 2008, 7, 294–306. [Google Scholar] [CrossRef]
- Mantzourani, E.D.; Blokar, K.; Tselios, T.V.; Matsoukas, J.M.; Platts, J.A.; Mavromoustakos, T.M.; Grdadolnik, S.G. A combined NMR and molecular dynamics simulation study to determine the conformational properties of agonists and antagonists against experimental autoimmune encephalomyelitis. Bioorg. Med. Chem. 2008, 16, 2171–2182. [Google Scholar] [CrossRef]
- Martel, P.; Makriyannis, A.; Mavromoustakos, T.; Kelly, K.; Jeffrey, K.R. Topography of tetrahydrocannabinol in model membranes using neutron diffraction. BBA Biomembr. 1993, 1151, 51–58. [Google Scholar] [CrossRef]
- Mavromoustakos, T.; Daliani, I. Effects of cannabinoids in membrane bilayers containing cholesterol. Biochim. Biophys. Acta Biomembr. 1999, 1420, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Mavromoustakos, T.; Theodoropoulou, E. A combined use of 13C-cross polarization/magic angle spinning, 13C-magic angle spinning and 31P-nuclear magnetic resonance spectroscopy with differential scanning calorimetry to study cannabinoid-membrane interactions. Chem. Phys. Lipids 1998, 92, 37–52. [Google Scholar] [CrossRef]
- Mavromoustakos, T.; De-Ping, Y.; Broderick, W.; Fournier, D.; Makriyannis, A. Small angle X-ray diffraction studies on the topography of cannabinoids in synaptic plasma membranes. Pharmacol. Biochem. Behav. 1991, 40, 547–552. [Google Scholar] [CrossRef]
- Mavromoustakos, T.; Yang, D.P.; Makriyannis, A. Topography of alphaxalone and Δ16-alphaxalone in membrane bilayers containing cholesterol. BBA Biomembr. 1994, 1194, 69–74. [Google Scholar] [CrossRef]
- Mavromoustakos, T.; Yang, D.P.; Makriyannis, A. Small angle X-ray diffraction and differential scanning calorimetric studies on O-methyl-(-)-Δ8-tetrahydrocannabinol and its 5′ iodinated derivative in membrane bilayers. BBA Biomembr. 1995, 1237, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Mavromoustakos, T.; Yang, D.P.; Makriyannis, A. Effects of the anesthetic steroid alphaxalone and its inactive Δ16-analog on the thermotropic properties of membrane bilayers. A model for membrane perturbation. BBA Biomembr. 1995, 1239, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Mavromoustakos, T.; Yang, D.P.; Makriyannis, A. Topography and thermotropic properties of cannabinoids in brain sphingomyelin bilayers. Life Sci. 1996, 59, 1969–1979. [Google Scholar] [CrossRef]
- Mavromoustakos, T.; Theodoropoulou, E.; Papahatjis, D.; Kourouli, T.; Yang, D.P.; Trumbore, M.; Makriyannis, A. Studies on the thermotropic effects of cannabinoids on phosphatidylcholine bilayers using differential scanning calorimetry and small angle X-ray diffraction. Biochim. Biophys. Acta Biomembr. 1996, 1281, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Mavromoustakos, T.; Theodoropoulou, E.; Yang, D.P. The use of high-resolution solid-state NMR spectroscopy and differential scanning calorimetry to study interactions of anaesthetic steroids with membrane. Biochim. Biophys. Acta Biomembr. 1997, 1328, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Mavromoustakos, T.; Papahatjis, D.; Laggner, P. Differential membrane fluidization by active and inactive cannabinoid analogues. Biochim. Biophys. Acta Biomembr. 2001, 1512, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Koukoulitsa, C.; Durdagi, S.; Siapi, E.; Villalonga-Barber, C.; Alexi, X.; Steele, B.R.; Micha-Screttas, M.; Alexis, M.N.; Tsantili-Kakoulidou, A.; Mavromoustakos, T. Comparison of thermal effects of stilbenoid analogs in lipid bilayers using differential scanning calorimetry and molecular dynamics: Correlation of thermal effects and topographical position with antioxidant activity. Eur. Biophys. J. 2011, 40, 865–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, J.N.H.; Illés, Z.; Reddy, J.; Keskin, D.B.; Fridkis-Hareli, M.; Kuchroo, V.K.; Strominger, J.L. Peptide 15-mers of defined sequence that substitute for random amino acid copolymers in amelioration of experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 2005, 102, 1620–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kant, R.; Pasi, S.; Surolia, A. Homo-β-amino acid containing MBP(85-99) analogs alleviate experimental autoimmune encephalomyelitis. Sci. Rep. 2015, 3, 8205. [Google Scholar] [CrossRef] [Green Version]
- Yannakakis, M.P.; Simal, C.; Tzoupis, H.; Rodi, M.; Dargahi, N.; Prakash, M.; Mouzaki, A.; Platts, J.A.; Apostolopoulos, V.; Tselios, T.V. Design and synthesis of non-peptide mimetics mapping the immunodominant myelin basic protein (MBP83-96) epitope to function as T-cell receptor antagonists. Int. J. Mol. Sci. 2017, 18, 1215. [Google Scholar] [CrossRef]
- Karin, N.; Mitchell, D.J.; Brocke, S.; Ling, N.; Steinman, L. Reversal of experimental autoimmune encephalomyelitis by a soluble peptide variant of a myelin basic protein epitope: T cell receptor antagonism and reduction of interferon γ and tumor necrosis factor α production. J. Exp. Med. 1994, 180, 2227–2237. [Google Scholar] [CrossRef] [Green Version]
- Katsara, M.; Matsoukas, J.; Deraos, G.; Apostolopoulos, V. Towards immunotherapeutic drugs and vaccines against multiple sclerosis. Acta Biochim. Biophys. Sin. 2008, 40, 636–642. [Google Scholar] [CrossRef] [Green Version]
- Aharoni, R.; Teitelbaum, D.; Arnon, R.; Sela, M. Copolymer 1 acts against the immunodominant epitope 82-100 of myelin basic protein by T cell receptor antagonism in addition to major histocompatibility complex blocking. Proc. Natl. Acad. Sci. USA 1999, 96, 634–639. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi-Milasi, F.; Mahnam, K.; Shakhsi-Niaei, M. In silico study of the association of the HLA-A*31:01 allele (human leucocyte antigen allele 31:01) with neuroantigenic epitopes of PLP (proteolipid protein), MBP (myelin basic protein) and MOG proteins (Myelin oligodendrocyte glycoprotein) for studying t. J. Biomol. Struct. Dyn. 2020, 1–17. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koukoulitsa, C.; Chontzopoulou, E.; Kiriakidi, S.; Tzakos, A.G.; Mavromoustakos, T. A Journey to the Conformational Analysis of T-Cell Epitope Peptides Involved in Multiple Sclerosis. Brain Sci. 2020, 10, 356. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060356
Koukoulitsa C, Chontzopoulou E, Kiriakidi S, Tzakos AG, Mavromoustakos T. A Journey to the Conformational Analysis of T-Cell Epitope Peptides Involved in Multiple Sclerosis. Brain Sciences. 2020; 10(6):356. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060356
Chicago/Turabian StyleKoukoulitsa, Catherine, Eleni Chontzopoulou, Sofia Kiriakidi, Andreas G. Tzakos, and Thomas Mavromoustakos. 2020. "A Journey to the Conformational Analysis of T-Cell Epitope Peptides Involved in Multiple Sclerosis" Brain Sciences 10, no. 6: 356. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060356