HLA-DPB1*03 as Risk Allele and HLA-DPB1*04 as Protective Allele for Both Early- and Adult-Onset Multiple Sclerosis in a Hellenic Cohort

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. HLA-DPB1* and HLA-DRB1* Genotyping
2.3. Statistical Analyses
3. Results
3.1. HWP and Linkage Disequilibrium of the Study’s Sample
3.2. Nongenetic Comparisons between Age of Onset Groups
3.3. HLA-DPB1 Allele Comparisons between the Age of Onset Groups
4. Discussion
5. Conclusions-Limitations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Availability of Data and Material
References
- Ramagopalan, S.V.; Dyment, D.A.; Ebers, G.C. Genetic epidemiology: The use of old and new tools for multiple sclerosis. Trends Neurosci. 2008, 31, 645–652. [Google Scholar] [CrossRef]
- Gruen, J.R.; Weissman, S.M. Human MHC Class III and IV Genes and Disease Associations. Front. Biosci. 2001, 6, D960–D972. [Google Scholar] [CrossRef] [PubMed]
- Katsavos, S.; Anagnostouli, M. Biomarkers in Multiple Sclerosis: An Up-to-Date Overview. Mult. Scler. Int. 2013, 2013, 340508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatelos, P.; Anagnostouli, M. HLA-Genotype in Multiple Sclerosis: The Role in Disease onset, Clinical Course, Cognitive Status and Response to Treatment: A Clear Step Towards Personalized Therapeutics. Immunogenet 2017, 2, 116. [Google Scholar]
- Goodin, D.; Khankhanian, P.; Gourraud, P.A.; Vince, N. Highly conserved extended haplotypes of the major histocompatibility complex and their relationship to multiple sclerosis susceptibility. PLoS ONE 2018, 13, e0190043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozikas, V.P.; Anagnostouli, M.C.; Petrikis, P.; Sitzoglou, C.; Phokas, C.; Tsakanikas, C.; Karavatos, A. Familial bipolar disorder and multiple sclerosis: A three-generation HLA family study. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 835–839. [Google Scholar] [CrossRef]
- International Multiple Sclerosis Genetics Consortium; Wellcome Trust Case Control Consortium 2; Sawcer, S.; Hellenthal, G.; Pirinen, M.; Spencer, C.C.; Patsopoulos, N.A.; Moutsianas, L.; Dilthey, A.; Su, Z.; et al. Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature 2011, 476, 214–219. [Google Scholar]
- Kular, L.; Liu, Y.; Ruhrmann, S.; Zheleznyakova, G.; Marabita, F.; Gomez-Cabrero, D.; James, T.; Ewing, E.; Lindén, M.; Górnikiewicz, B.; et al. DNA Methylation as a Mediator of HLA-DRB1*15:01 and a Protective Variant in Multiple Sclerosis. Nat. Commun. 2018, 9, 2397. [Google Scholar] [CrossRef]
- Gianfrancesco, M.; Stridh, P.; Shao, X.; Rhead, B.; Graves, J.S.; Chitnis, T.; Waldman, A.; Lotze, T.; Schreiner, T.; Belman, A.; et al. Genetic risk factors for pediatric–onset multiple sclerosis. Mult. Scler. 2018, 24, 1825–1834. [Google Scholar] [CrossRef]
- Venkateswaran, S.; Banwell, B. Pediatric multiple sclerosis. Neurologist 2010, 16, 92–105. [Google Scholar] [CrossRef]
- Anagnostouli, M.C.; Manouseli, A.; Artemiadis, A.; Katsavos, S.; Fillipopoulou, C.; Youroukos, S.; Efthimiopoulos, S.; Doxiadis, I. HLA-DRB1* Allele Frequencies in Pediatric, Adolescent and Adult-Onset Multiple Sclerosis Patients, in a Hellenic Sample. Evidence for New and Established Associations. J. Mult. Scler. 2014, 1, 104. [Google Scholar] [CrossRef] [Green Version]
- Odum, N.; Hyldic-Nielsenn, J.J.; Morling, N.; Sandberg-Wollheim, M.; Platz, P.; Svejgaard, A. HLA-DP antigens are involved in the susceptibility to multiple sclerosis. Tissue Antigens 1988, 31, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.M.; Wang, C.; Zhang, K.N.; Lin, A.Y.; Kira, J.; Hu, G.Z.; Qu, X.H.; Xiong, Y.Q.; Cao, W.F.; Gong, L.Y. Association of susceptibility to multiple sclerosis in Southern Han Chinese with HLA-DRB1, -DPB1 alleles and DRB1-DPB1 haplotypes: Distinct from other populations. Mult. Scler. 2009, 15, 1422–1430. [Google Scholar] [CrossRef]
- Fukazawa, T.; Yamasaki, K.; Ito, H.; Kikuchi, S.; Minohara, M.; Horiuchi, I.; Tsukishima, E.; Sasaki, H.; Hamada, T.; Nishimura, Y.; et al. Both the HLA-DPB1 and -DRB1 alleles correlate with risk for multiple sclerosis in Japanese: Clinical phenotypes and gender as important factors. Tissue Antigens 2000, 55, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, T.; Kikuchi, S.; Miyagishi, R.; Miyazaki, Y.; Yabe, I.; Hamada, T.; Sasaki, H. HLA-DPB1*0501 is not uniquely associated with opticospinal multiple sclerosis in Japanese patients. Important role of DPB1*0301. Mult. Scler. 2006, 12, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, S.; Isobe, N.; Yonekawa, T.; Matsushita, T.; Masaki, K.; Sato, S.; Kawano, Y.; Yamamoto, K.; Kira, J.; South Japan Multiple Sclerosis Genetics Consortium. Genetic and infectious profiles of Japanese multiple sclerosis patients. PLoS ONE 2012, 7, e48592. [Google Scholar] [CrossRef]
- Yu, M.; Kinkel, R.P.; Weinstock-Guttman, B.; Cook, D.J.; Tuohy, V.K. HLADP: A class II restriction molecule involved in epitope spreading during the development of multiple sclerosis. Hum. Immunol. 1998, 59, 15–24. [Google Scholar] [CrossRef]
- Field, J.; Browning, S.R.; Johnson, L.J.; Danoy, P.; Varney, M.D.; Tait, B.D.; Gandhi, K.S.; Charlesworth, J.C.; Heard, R.N.; Australia and New Zealand Multiple Sclerosis Genetics Consortium; et al. A polymorphism in the HLA-DPB1 gene is associated with susceptibility to multiple sclerosis. PLoS ONE 2010, 5, e13454. [Google Scholar] [CrossRef] [Green Version]
- Dekker, J.W.; Easteal, S.; Jakobsen, I.B.; Gao, X.; Stewart, G.J.; Buhler, M.M.; Hawkins, B.R.; Higgins, D.A.; Yu, Y.L.; Serjeantson, S.W. HLA-DPB1 alleles correlate with risk for multiple sclerosis in Caucasoid and Cantonese patients lacking the high-risk DQB1*0602 allele. Tissue Antigens 1993, 41, 31–36. [Google Scholar] [CrossRef]
- Roth, M.P.; Coppin, H.; Descoins, P.; Ruidavets, J.B.; Cambon-Thomsen, A.; Clanet, M. HLA-DPB1 gene polymorphism and multiple sclerosis: A large case-control study in the southwest of France. J. Neuroimmunol. 1991, 34, 215–222. [Google Scholar] [CrossRef]
- Marrosu, M.G.; Cocco, E.; Costa, G.; Murru, M.R.; Mancosu, C.; Murru, R.; Lai, M.; Sardu, C.; Contu, P. Interaction of loci within the HLA region influences multiple sclerosis course in the Sardinian population. J. Neurol. 2006, 253, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Patsopoulos, N.A.; Barcellos, L.F.; Hintzen, R.Q.; Schaefer, C.; van Duijn, C.M.; Noble, J.A.; Raj, T.; IMSGC; ANZgene; Gourraud, P.A.; et al. Fine-mapping the genetic association of the major histocompatibility complex in multiple sclerosis: HLA and non-HLA effects. PLoS Genet. 2013, 9, e1003926. [Google Scholar]
- Gontika, M.P.; Anagnostouli, M.C. Human leukocyte antigens immunogenetics of neuromyelitis optica or Devic’s disease and the impact on the immunopathogenesis, diagnosis and treatment: A critical review. Neuroimmunol. Neuroinflamm. 2014, 1, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Hersh, A.O.; Prahalad, S. Immunogenetics of juvenile idiopathic arthritis: A comprehensive review. J. Autoimmun. 2015, 64, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimaki, K.; Kawamura, T.; Inada, H.; Yagawa, K.; Nose, Y.; Nabeya, N.; Isshiki, G.; Tatsumi, N.; Niihira, S. HLA DPB1*0201 gene confers disease susceptibility in Japanese with childhood onset type I diabetes, independent of HLA-DR and DQ genotypes. Diabetes Res. Clin. Pract. 2000, 47, 49–55. [Google Scholar] [CrossRef]
- Sato, S.; Isobe, N.; Yoshimura, S.; Kanamori, Y.; Masaki, K.; Matsushita, T.; Kira, J. HLA-DPB1*0201 is associated with susceptibility to atopic myelitis in Japanese. J. Neuroimmunol. 2012, 251, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, M.R.; Ramagopalan, S.V.; Chao, M.J.; Herrera, B.M.; Deluca, G.C.; Orton, S.M.; Dyment, D.A.; Sadovnick, A.D.; Ebers, G.C. Epistasis among HLA-DRB1, HLA-DQA1, and HLA-DQB1 loci determines multiple sclerosis susceptibility. Proc. Natl. Acad. Sci. USA 2009, 106, 7542–7547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moutsianas, L.; Jostins, L.; Beecham, A.H.; Dilthey, A.T.; Xifara, D.K.; Ban, M.; Shah, T.S.; Patsopoulos, N.A.; Alfredsson, L.; Anderson, C.A.; et al. Class II HLA interactions modulate genetic risk for multiple sclerosis. Nat. Genet. 2015, 47, 1107–1113. [Google Scholar]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar] [CrossRef] [Green Version]
- Marsh, S.G. Nomenclature for factors of the HLA system, update June 2012. Tissue Antigens 2012, 80, 289–293. [Google Scholar] [CrossRef]
- Lancaster, A.K.; Single, R.M.; Solberg, O.D.; Nelson, M.P.; Thomson, G. PyPop update—A software pipeline for large-scale multilocus population genomics. Tissue Antigens 2007, 69 (Suppl. 1), 192–197. [Google Scholar] [CrossRef] [Green Version]
- Papassavas, E.C.; Spyropoulou-Vlachou, M.; Papassavas, A.C.; Schipper, R.F.; Doxiadis, I.N.; Stavropoulos-Giokas, C. MHC class I and class II phenotype, gene, and haplotype frequencies in Greeks using molecular typing data. Hum. Immunol. 2000, 61, 615–623. [Google Scholar] [CrossRef]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Narum, S.R. Beyond Bonferroni: Less conservative analyses for conservation genetics. Conserv. Genet. 2006, 7, 783–787. [Google Scholar] [CrossRef]
- Hadley, D.; Hagopian, W.; Liu, E.; She, J.X.; Simell, O.; Akolkar, B.; Ziegler, A.G.; Rewers, M.; Krischer, J.P.; Chen, W.M.; et al. HLA-DPB1*04:01 Protects Genetically Susceptible Children from Celiac Disease Autoimmunity in the TEDDY Study. Am. J. Gastroenterol. 2015, 110, 915–920. [Google Scholar] [CrossRef] [Green Version]
- Ollila, H.M.; Ravel, J.M.; Han, F.; Faraco, J.; Lin, L.; Zheng, X.; Plazzi, G.; Dauvilliers, Y.; Pizza, F.; Hong, S.C.; et al. HLA-DPB1 and HLA class I confer risk of and protection from narcolepsy. Am. J. Hum. Genet. 2015, 96, 852. [Google Scholar] [CrossRef] [Green Version]
- Misicka, E.; Sept, C.; Briggs, F.B.S. Predicting onset of secondary-progressive multiple sclerosis using genetic and non-genetic factors. J. Neurol. 2020. [Google Scholar] [CrossRef]
- Gontika, M.P.; Anagnostouli, M.C. Anti-Myelin Oligodendrocyte Glycoprotein and Human Leukocyte Antigens as Markers in Pediatric and Adolescent Multiple Sclerosis: On Diagnosis, Clinical Phenotypes, and Therapeutic Responses. Mult. Scler. Int. 2018, 2018, 8487471. [Google Scholar] [CrossRef] [Green Version]
- Uher, T.; McComb, M.; Galkin, S.; Srpova, B.; Oechtering, J.; Barro, C.; Tyblova, M.; Bergsland, N.; Krasensky, J.; Dwyer, M.; et al. Neurofilament levels are associated with blood-brain barrier integrity, lymphocyte extravasation, and risk factors following the first demyelinating event in multiple sclerosis. Mult. Scler. 2020. [Google Scholar] [CrossRef]
- Warabi, Y.; Matsumoto, Y.; Hayashi, H. Interferon beta1b exacerbates multiple sclerosis with severe optic nerve and spinal cord demyelination. J. Neurol. Sci. 2007, 252, 57–61. [Google Scholar] [CrossRef]
- Werneck, L.C.; Lorenzoni, P.J.; Kay, C.S.K.; Scola, R.H. Multiple sclerosis: Disease modifying therapy and the human leukocyte antigen. Arq. Neuropsiquiatr. 2018, 76, 697–704. [Google Scholar] [CrossRef] [PubMed]


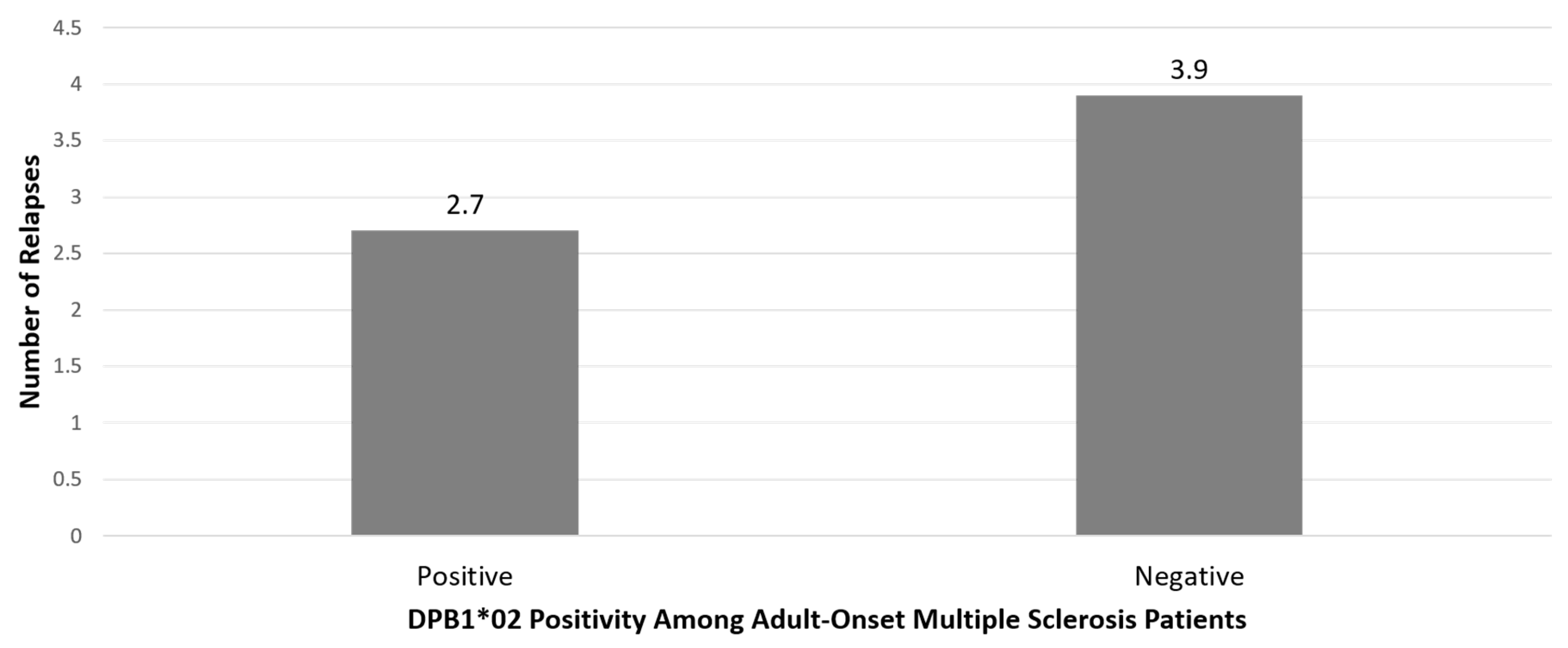{kind=link}
| Characteristics | EOMS | AOMS | Sig 1 |
|---|---|---|---|
| Females | 19/28 (67.9%) | 43/72 (59.7%) | 0.499 |
| Age (years old) | 29.9 ± 9.8 | 39.8 ± 10.8 | 0.001 * |
| Duration of MS (months) | 148.1 ± 105.4 | 81.3 ± 69.1 | 0.011 * |
| EDSS | 3.1 ± 1.7 | 3.3 ± 1.6 | 0.414 |
| Primary Progressive | 3/28 (10.7%) | 7/72 (9.7%) | 0.917 |
| Relapses since onset | 5.2 ± 6.1 | 3.7 ± 2.5 | 0.393 |
| IgG index 2 | 1.3 ± 0.7 | 0.8 ± 0.4 | 0.004 * |
| Presence of OCBs 2 | 13/15 (86.7%) | 31/45 (68.9%) | 0.312 |
| Subcortical lesions 2 | 12/17 (70.6%) | 27/43 (62.8%) | 0.765 |
| Periventricular lesions 2 | 16/17 (94.1%) | 42/43 (97.7%) | 0.49 |
| Infratentorial lesions 2 | 12/17 (70.6%) | 30/43 (69.8%) | 1.000 |
| Spinal cord lesions 2 | 12/14 (85.7%) | 29/31 (93.5%) | 0.578 |
| Early MS | Adult MS | HCs | Early vs. Adult MS | Early MS vs. HCs | Adult MS vs. HCs | |
|---|---|---|---|---|---|---|
| (N = 28) | (N = 72) | (N = 246) | Sig 1 | Sig 2 | Sig 2 | |
| HLA-DPB1*01 | 3.6 | 4.2 | 4.5 | 0.85 (0.09–8.55) 1.000 | 0.79 (0.1–6.37) 0.500 | 0.92 (0.25–3.42) 0.500 |
| HLA-DPB1*02 | 39.3 | 22.2 | 36.6 | 2.27 (0.89–5.80) 0.131 | 1.12 (0.5–2.5) 0.461 | 0.5 (0.23–0.91) 0.008 ** |
| HLA-DPB1*03 | 17.9 | 23.6 | 13.4 | 0.7 (0.23–2.13) 0.602 | 1.4 (0.50–3.95) 0.339 | 2.0 (1.04–3.84) 0.009 ** |
| HLA-DPB1*04 | 64.3 | 79.2 | 92.7 | 0.47 (0.18–1.24) 0.132 | 0.14 (0.06–0.35) <0.001 ** | 0.3 (0.14–0.63) <0.001 ** |
| HLA-DPB1*05 | 3.6 | 2.8 | 2.4 | 1.3 (0.11–14.89) 1.000 | 1.48 (0.17–12.77) 0.500 | 1.14 (0.23–5.79) 0.500 |
| HLA-DPB1*06 | 0 | 1.4 | 0.8 | - 1.000 | - 0.500 | 1.72 (0.15–19.23) 0.500 |
| HLA-DPB1*09 | 0 | 1.4 | 2.8 | - 1.000 | - 0.372 | 0.48 (0.06-3.97) 0.356 |
| HLA-DPB1*10 | 10.7 | 9.7 | 4.9 | 1.11 (0.27–4.65) 1.000 | 2.34 (0.62–8.85) 0.162 | 2.1 (0.8–5.55) 0.052 |
| HLA-DPB1*11 | 0 | 1.4 | - | - 1.000 | - | - |
| HLA-DPB1*13 | 3.6 | 2.8 | 6.1 | 1.3 (0.11–14.89) 1.000 | 0.57 (0.07–4.49) 0.435 | 0.44 (0.1–1.97) 0.176 |
| HLA-DPB1*14 | 7.1 | 6.9 | 3.7 | 1 (0.18–5.48) 1.000 | 2.03 (0.42–9.88) 0.321 | 1.97 (0.64–6.06) 0.126 |
| HLA-DPB1*15 | 0 | 4.2 | 2.0 | - 0.557 | - 0.468 | 2.1 (0.49–8.99) 0.186 |
| HLA-DPB1*19 | 0 | 1.4 | 0.4 | - 1.000 | - 0.500 | 3.45 (0.21–55.87) 0.346 |
| HLA-DPB1*22 | 3.6 | 0 | - | - 0.280 | - | - |
| HLA-DPB1*23 | 3.6 | 0 | 2.4 | - 0.280 | 1.48 (0.17–12.77) 0.500 | - 0.172 |
| HLA-DPB1*32 | 3.6 | 0 | 0.8 | - 0.280 | 4.52 (0.4–51.49) 0.279 | - 0.460 |
| HLA-DPB1*33 | 3.6 | 2.8 | 0.4 | 1.3 (0.11–14.89) 1.000 | 9.07 (0.55–149.24) 0.123 | 7.0 (0.63–78.34) 0.130 |
| HLA-DPB1*34 | 0 | 1.4 | - | - 1.000 | - | - |
| HLA-DPB1*35 | 0 | 4.2 | 0.8 | - 0.557 | - 0.500 | 10.65 (1.09–104.03) 0.005 ** |
| HLA-DPB1*38 | 3.6 | 0 | - | - 0.280 | - | - |
| HLA-DPB1*46 | 3.6 | 0 | - | - 0.280 | - | - |
| HLA-DPB1*50 | 0 | 1.4 | 0.4 | - 1.000 | - 0.500 | 3.45 (0.21–55.87) 0.346 |
| HLA-DPB1*56 | 0 | 1.4 | - | - 1.000 | - | - |
| Total Sample of MS Patients | ||
| HLA-DPB1* Genotype | HLA-DRB1*15 Positive | Sig 1 |
| HLA-DPB1*03 | 0/22 (0%) | 0.001 * |
| HLA-DPB1*04 | 19/63 (30.2%) | 0.048 * |
| HLA-DPB1*14 | 5/7 (71.4%) | 0.008 * |
| Adult-Onset MS | ||
| HLA-DPB1* Genotype | HLA-DRB1*15 Positive | Sig 1 |
| HLA-DPB1*03 | 0/17 (0%) | 0.003 * |
| Early-Onset MS | ||
| HLA-DPB1* Genotype | HLA-DRB1*15 Positive | Sig 1 |
| HLA-DPB1*14 | 2/2 (100%) | 0.036 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anagnostouli, M.; Artemiadis, A.; Gontika, M.; Skarlis, C.; Markoglou, N.; Katsavos, S.; Kilindireas, K.; Doxiadis, I.; Stefanis, L. HLA-DPB1*03 as Risk Allele and HLA-DPB1*04 as Protective Allele for Both Early- and Adult-Onset Multiple Sclerosis in a Hellenic Cohort. Brain Sci. 2020, 10, 374. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060374
Anagnostouli M, Artemiadis A, Gontika M, Skarlis C, Markoglou N, Katsavos S, Kilindireas K, Doxiadis I, Stefanis L. HLA-DPB1*03 as Risk Allele and HLA-DPB1*04 as Protective Allele for Both Early- and Adult-Onset Multiple Sclerosis in a Hellenic Cohort. Brain Sciences. 2020; 10(6):374. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060374
Chicago/Turabian StyleAnagnostouli, Maria, Artemios Artemiadis, Maria Gontika, Charalampos Skarlis, Nikolaos Markoglou, Serafeim Katsavos, Konstantinos Kilindireas, Ilias Doxiadis, and Leonidas Stefanis. 2020. "HLA-DPB1*03 as Risk Allele and HLA-DPB1*04 as Protective Allele for Both Early- and Adult-Onset Multiple Sclerosis in a Hellenic Cohort" Brain Sciences 10, no. 6: 374. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060374