Neurochemical Correlates of Brain Atrophy in Fibromyalgia Syndrome: A Magnetic Resonance Spectroscopy and Cortical Thickness Study

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
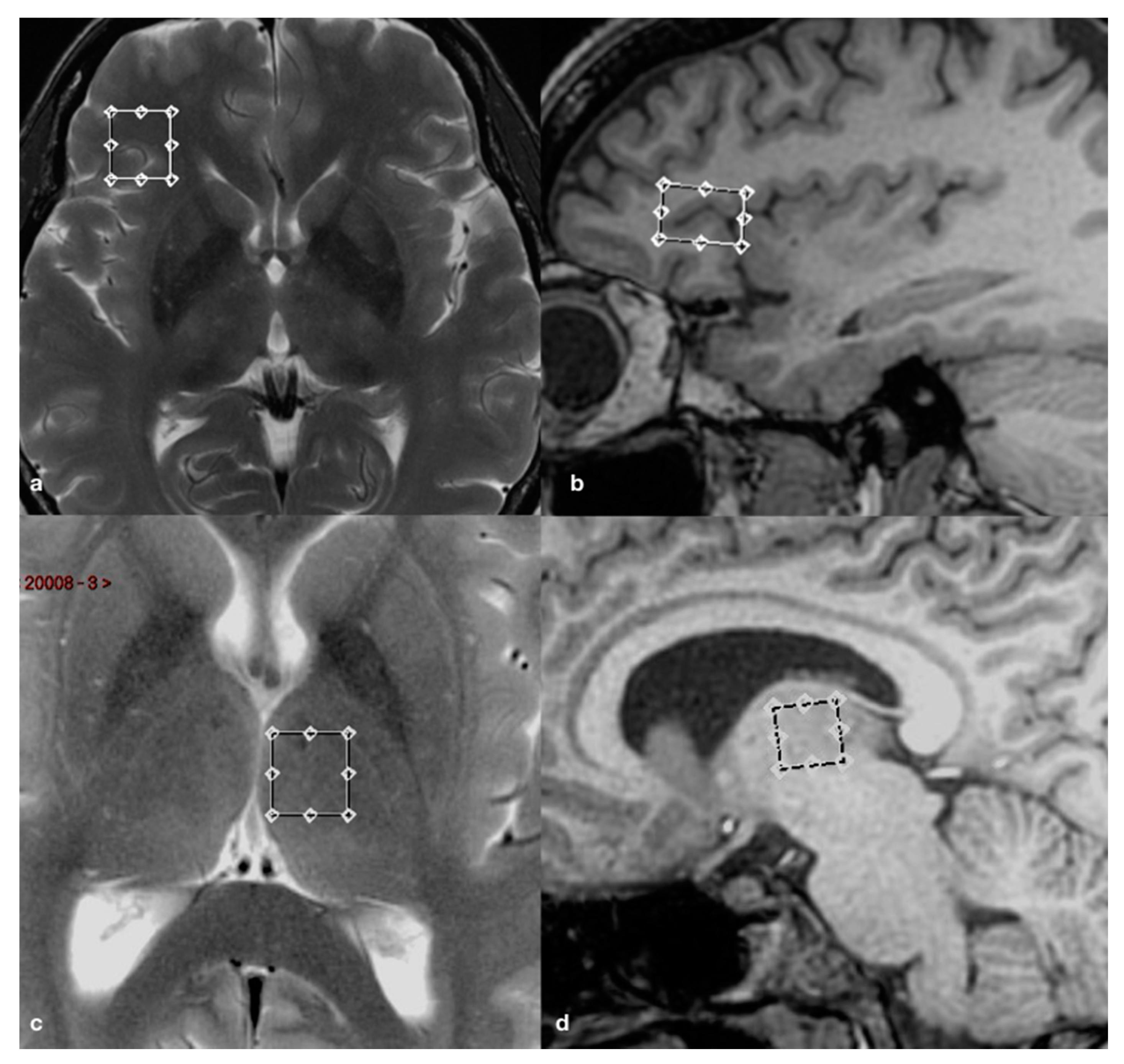
2.2. Brain Imaging and Spectroscopy
2.3. Cortical and Subcortical Segmentations
2.4. Statistical Analysis
3. Results
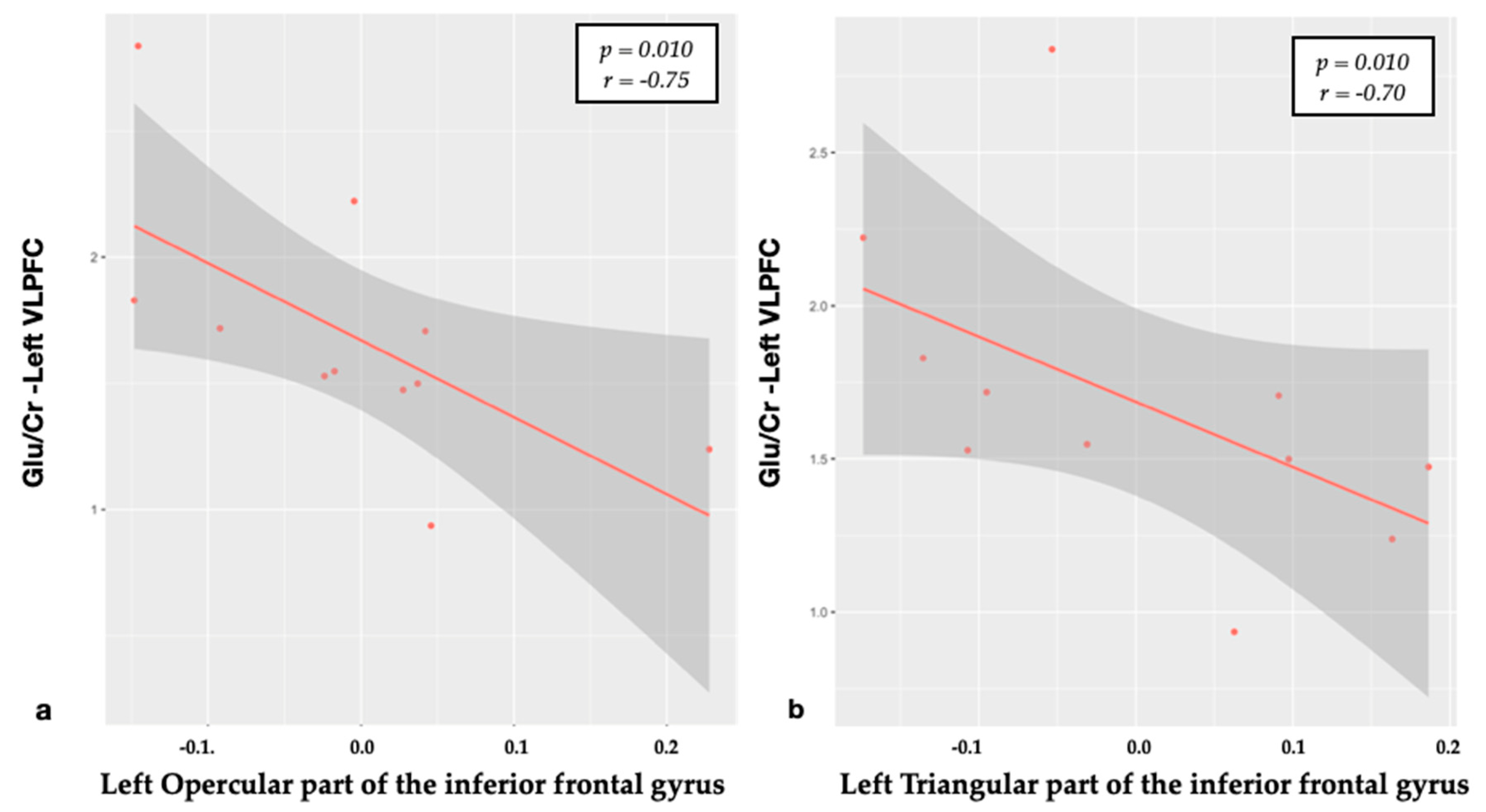
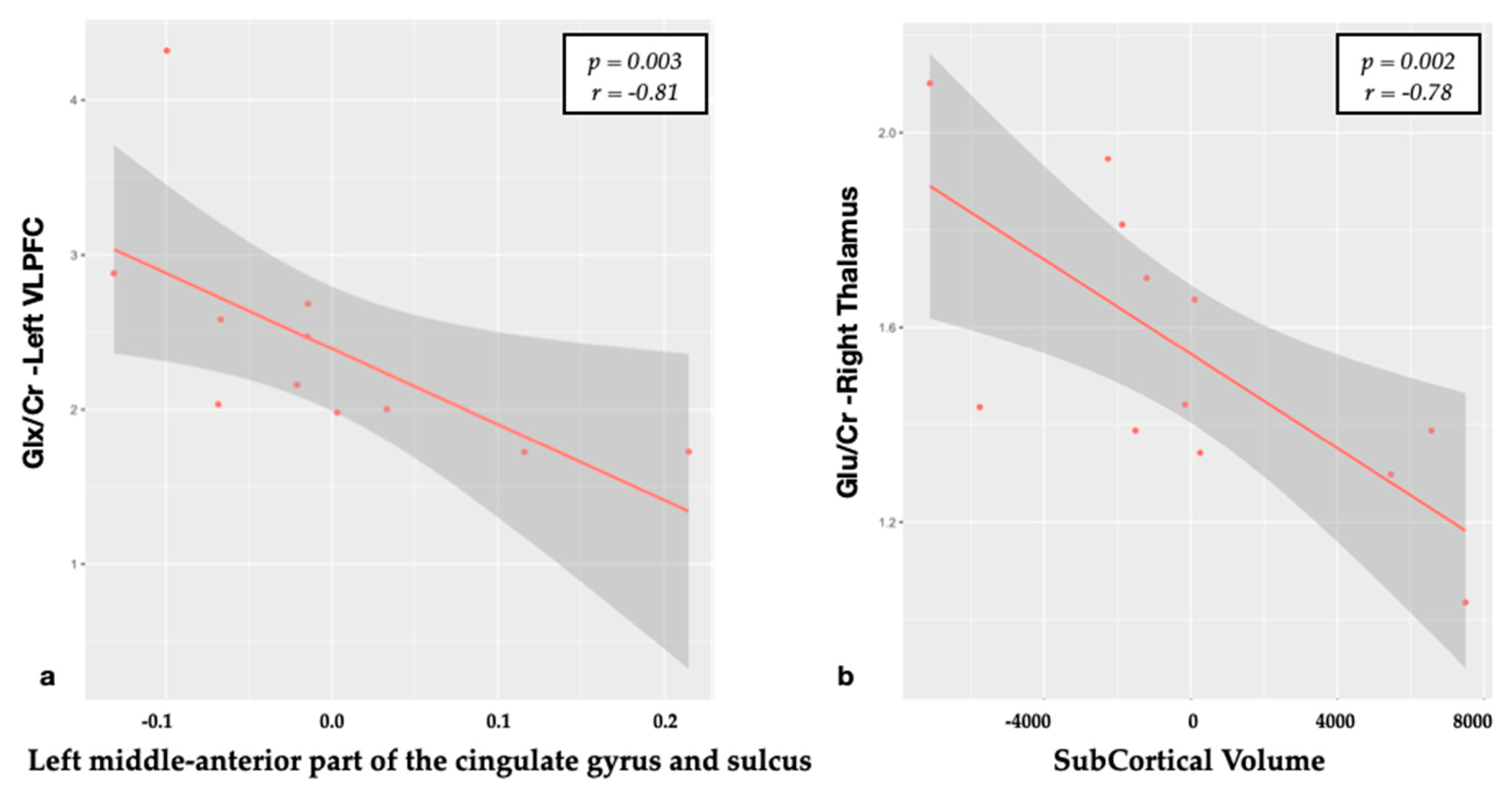
3.1. Relationship between Metabolites and Brain Structural Data
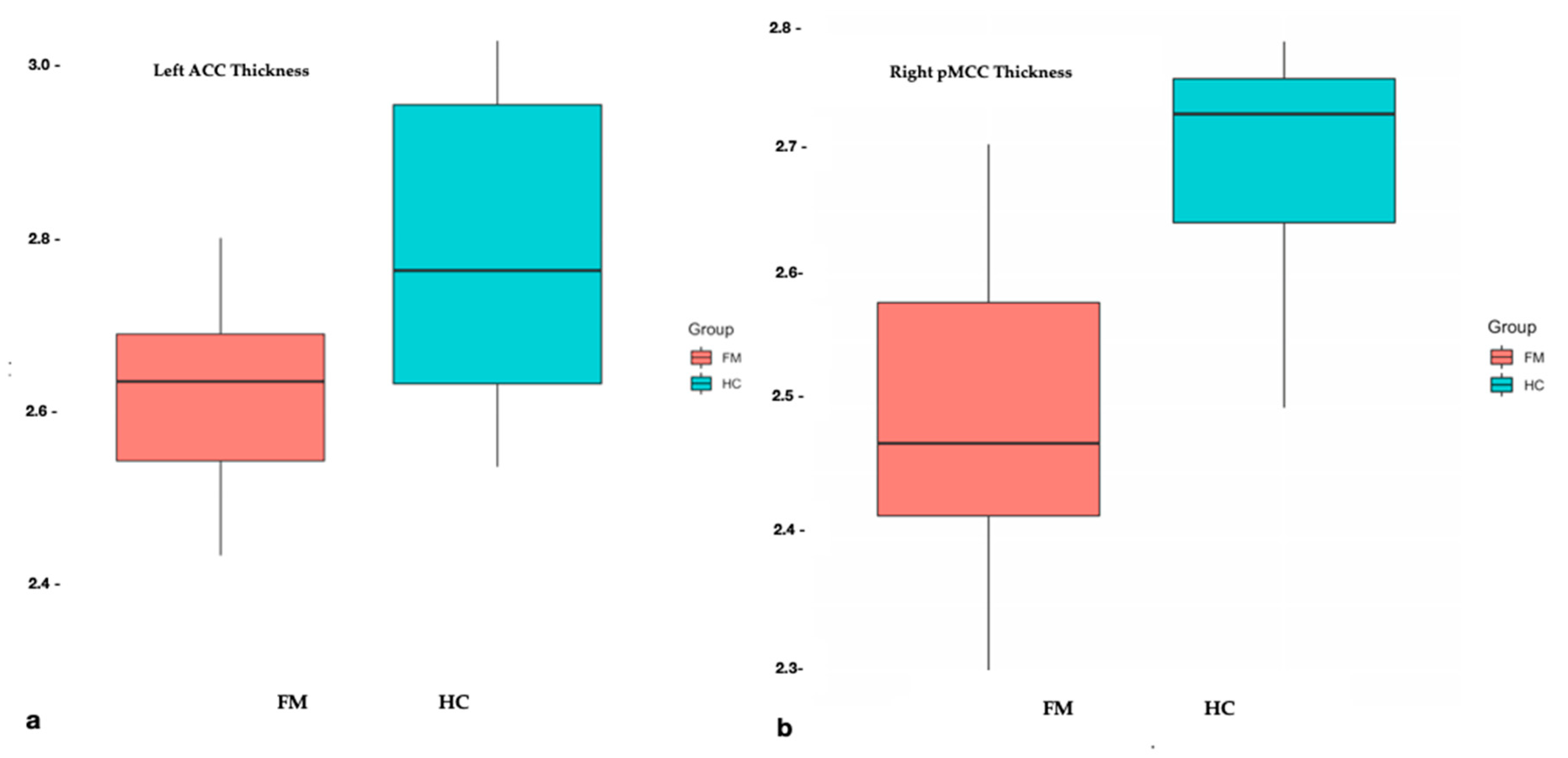
3.2. Structural Differences and Clinical Relationships
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wolfe, F.; A Smythe, H.; Yunus, M.B.; Bennett, R.M.; Bombardier, C.; Goldenberg, D.L.; Tugwell, P.; Campbell, S.M.; Abeles, M.; Clark, P. The American College of Rheumatology 1990 Criteria for the Classification of Fibromyalgia. Report of the Multicenter Criteria Committee. Arthritis Rheum. 1990, 33, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Yunus, M.B. Fibromyalgia and Overlapping Disorders: The Unifying Concept of Central Sensitivity Syndromes. Semin. Arthritis Rheum. 2007, 36, 339–356. [Google Scholar] [CrossRef] [PubMed]
- Staud, R. Evidence for Shared Pain Mechanisms in Osteoarthritis, Low Back Pain, and Fibromyalgia. Curr. Rheumatol. Rep. 2011, 13, 513–520. [Google Scholar] [CrossRef]
- Latremoliere, A.; Woolf, C.J. Central Sensitization: A Generator of Pain Hypersensitivity by Central Neural Plasticity. J. Pain 2009, 10, 895–926. [Google Scholar] [CrossRef] [Green Version]
- Pyke, T.L.; Osmotherly, P.; Baines, S. Measuring Glutamate Levels in the Brains of Fibromyalgia Patients and a Potential Role for Glutamate in the Pathophysiology of Fibromyalgia Symptoms. Clin. J. Pain 2017, 33, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Napadow, V.; Harris, R.E. What has functional connectivity and chemical neuroimaging in fibromyalgia taught us about the mechanisms and management of ‘centralized’ pain? Arthritis Res. Ther. 2014, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleich, S.; Wiltfang, J.; Kornhuber, J. Glutamate and the glutamate receptor system: A target for drug action. Int. J. Geriatr. Psychiatry 2003, 18, S33–S40. [Google Scholar] [CrossRef]
- Perez-Rodriguez, R.; Oliván, A.; Roncero, C.; Morón-Oset, J.; Gonzalez, M.; Oset-Gasque, M.J. Glutamate triggers neurosecretion and apoptosis in bovine chromaffin cells through a mechanism involving NO production by neuronal NO synthase activation. Free. Radic. Biol. Med. 2014, 69, 390–402. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, X.; Bhavnani, B.R. Equine estrogens differentially inhibit DNA fragmentation induced by glutamate in neuronal cells by modulation of regulatory proteins involved in programmed cell death. BMC Neurosci. 2003, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- A Kosten, T.; Galloway, M.; Duman, R.S.; Russell, D.S.; D’Sa, C. Repeated Unpredictable Stress and Antidepressants Differentially Regulate Expression of the Bcl-2 Family of Apoptotic Genes in Rat Cortical, Hippocampal, and Limbic Brain Structures. Neuropsychopharmacol. 2007, 33, 1545–1558. [Google Scholar] [CrossRef]
- Sawaddiruk, P.; Paiboonworachat, S.; Chattipakorn, N.; Chattipakorn, S.C. Alterations of brain activity in fibromyalgia patients. J. Clin. Neurosci. 2017, 38, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Feraco, P.; Bacci, A.; Pedrabissi, F.; Passamonti, L.; Zampogna, G.; Pedrabissi, F.; Malavolta, N.; Leonardi, M. Metabolic abnormalities in pain-processing regions of patiNents with fiburomyradiologia: A 3T MR spectroscopy study. Am. J. Neuroradiol. 2020, 32, 1585–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provencher, S.W. Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn. Reson. Med. 1993, 30, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Dale, A.M.; Fischl, B.; Sereno, M.I. Cortical Surface-Based Analysis: I. Segmentation and surface reconstruction. NeuroImage 1999, 9, 179–194. [Google Scholar] [CrossRef]
- Fischl, B. FreeSurfer. NeuroImage 2012, 62, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Fischl, B.; Sereno, M.I.; Dale, A.M. Cortical Surface-Based Analysis. II: Inflation, flattening, and a surface-basedcoordinate system. NeuroImage 1999, 9, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Destrieux, C.; Fischl, B.; Dale, A.; Halgren, E. Automatic parcellation of human cortical gyri and sulci using standard anatomical nomenclature. NeuroImage 2010, 53, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fischl, B.; Salat, D.H.; Busa, E.; Albert, M.; Dieterich, M.; Haselgrove, C.; Van Der Kouwe, A.; Killiany, R.; Kennedy, D.; Klaveness, S.; et al. Whole brain segmentation: Automated labeling of neuroanatomical structures in the human brain. Neuron 2002, 33, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Petrovic, P.; Kalso, E.; Petersson, K.M.; Andersson, J.; Fransson, P.; Ingvar, M. A prefrontal non-opioid mechanism in placebo analgesia. Pain 2010, 150, 59–65. [Google Scholar] [CrossRef]
- Badre, D.; Wagner, A.D. Left ventrolateral prefrontal cortex and the cognitive control of memory. Neuropsychologia 2007, 45, 2883–2901. [Google Scholar] [CrossRef]
- Jensen, K.B.; Loitoile, R.; Kosek, E.; Petzke, F.; Carville, S.; Fransson, P.; Marcus, H.; Williams, S.C.; Choy, E.; Mainguy, Y.; et al. Patients with fibromyalgia display less functional connectivity in the brain’s pain inhibitory network. Mol. Pain 2012, 8, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, K.B.; Srinivasan, P.; Spaeth, R.; Tan, Y.; Kosek, E.; Petzke, F.; Carville, S.; Fransson, P.; Marcus, H.; Williams, S.C.; et al. Overlapping structural and functional brain changes in patients with long-term exposure to fibromyalgia pain. Arthritis Rheum. 2013, 65, 3293–3303. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Piedra, C.; Guzmán, M.; Buela-Casal, G.; Catena, A. The impact of fibromyalgia symptoms on brain morphometry. Brain Imaging Behav. 2015, 10, 1184–1197. [Google Scholar] [CrossRef]
- Wood, P.B.; Glabus, M.F.; Simpson, R.; Patterson, J.C. Changes in Gray Matter Density in Fibromyalgia: Correlation with Dopamine Metabolism. J. Pain 2009, 10, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.B.; Kosek, E.; Petzke, F.; Carville, S.; Fransson, P.; Marcus, H.; Williams, S.C.; Choy, E.; Giesecke, T.; Mainguy, Y.; et al. Evidence of dysfunctional pain inhibition in Fibromyalgia reflected in rACC during provoked pain. Pain 2009, 144, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Napadow, V.; LaCount, L.; Park, K.; As-Sanie, S.; Clauw, D.J.; Harris, R.E.; As-Sanie, S. Intrinsic brain connectivity in fibromyalgia is associated with chronic pain intensity. Arthritis Rheum. 2010, 62, 2545–2555. [Google Scholar] [CrossRef] [Green Version]
- Frøkjær, J.B.; Olesen, A.E.; Simonsen, C.W.; Drewes, A.M.; Hansen, T.M. Cingulate metabolites during pain and morphine treatment as assessed by magnetic resonance spectroscopy. J. Pain Res. 2014, 7, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Vogt, B.A. Pain and emotion interactions in subregions of the cingulate gyrus. Nat. Rev. Neurosci. 2005, 6, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Vogt, B.A.; Berger, G.R.; Derbyshire, S.W.G. Structural and functional dichotomy of human midcingulate cortex. Eur. J. Neurosci. 2003, 18, 3134–3144. [Google Scholar] [CrossRef] [Green Version]
- Miguel, N.F.; García-Campayo, J.; Magallón, R.; Andrés-Bergareche, H.; Luciano, J.V.; Andrés-Esteban, E.M.; Beltrán, J. Localized 1H-NMR spectroscopy in patients with fibromyalgia: A controlled study of changes in cerebral glutamate/glutamine, inositol, choline, and N-acetylaspartate. Arthritis Res. Ther. 2010, 12, R134. [Google Scholar] [CrossRef] [Green Version]
- Mullins, P.G.; Rowland, L.M.; Jung, R.; Sibbitt, W.L. A novel technique to study the brain’s response to pain: Proton magnetic resonance spectroscopy. NeuroImage 2005, 26, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Shackman, A.J.; Salomons, T.; Slagter, H.; Fox, A.S.; Winter, J.; Davidson, R.J. The integration of negative affect, pain and cognitive control in the cingulate cortex. Nat. Rev. Neurosci. 2011, 12, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Willnow, T.E. Excitatory Amino Acid Transporters in Physiology and Disorders of the Central Nervous System. Int. J. Mol. Sci. 2019, 20, 5671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Casillas, A.; Fernández-Guasti, A. The dose makes the poison: From glutamate-mediated neurogenesis to neuronal atrophy and depression. Rev. Neurosci. 2016, 27, 599–622. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Lee, S.-H.; Weng, H.-H. Gray Matter Atrophy within the Default Mode Network of Fibromyalgia: A Meta-Analysis of Voxel-Based Morphometry Studies. BioMed Res. Int. 2016, 2016, 1–9. [Google Scholar] [CrossRef]
- Harris, R.E.; Sundgren, P.; Craig, A.; Kirshenbaum, E.; Sen, A.; Napadow, V.; Clauw, D.J. Elevated insular glutamate in fibromyalgia is associated with experimental pain. Arthritis Rheum. 2009, 60, 3146–3152. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.E.; Napadow, V.; Huggins, J.P.; Pauer, L.; Kim, J.; Hampson, J.; Sundgren, P.; Foerster, B.; Petrou, M.; Schmidt-Wilcke, T.; et al. Pregabalin Rectifies Aberrant Brain Chemistry, Connectivity, and Functional Response in Chronic Pain Patients. Anesthesiol. 2013, 119, 1453–1464. [Google Scholar] [CrossRef] [Green Version]
- Ceko, M.; Bushnell, M.C.; Fitzcharles, M.-A.; Schweinhardt, P. Fibromyalgia interacts with age to change the brain. NeuroImage: Clin. 2013, 3, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Choe, M.K.; Lim, M.; Kim, J.S.; Lee, N.S.; Chung, C.K. Disrupted Resting State Network of Fibromyalgia in Theta frequency. Sci. Rep. 2018, 8, 2064. [Google Scholar] [CrossRef]
- Bergink, V. Glutamate and anxiety. Eur. Neuropsychopharmacol. 2004, 14, 175–183. [Google Scholar] [CrossRef]
- Wolfe, F.; Clauw, D.J.; Fitzcharles, M.A.; Goldenberg, D.L.; Häuser, W.; Katz, R.L.; Mease, P.J.; Russell, A.S.; Russell, I.J.; Walitt, B. 2016 Revisions to the 2010/2011 fibromyalgia diagnostic criteria. Semin Arthritis Rheum. 2016, 46, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, F.; Butler, S.H.; Fitzcharles, M.; Häuser, W.; Katz, R.L.; Mease, P.J.; Rasker, J.J.; Russell, A.S.; Russell, I.J.; Walitt, B. Revised chronic widespread pain criteria: Development from and integration with fibromyalgia criteria. Scand. J. Pain 2019, 20, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka-Wójcik, E.; Kłodzinska, A.; Pilc, A. Glutamate receptor ligands as anxiolytics. Curr. Opin. Investig. Drugs 2001, 2, 1112–1119. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of HC | Age, Year | Sex | No. of Pt | AGE, Year | Sex | Onset, Year | Duration, Year | TP | VAS | FIQ |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 33 | F | 1 | 34 | F | 30 | 4 | 11 | 47 | 48.4 |
| 2 | 54 | F | 2 | 54 | F | 45 | 9 | 18 | 87 | 83.15 |
| 3 | 41 | F | 3 | 44 | F | 40 | 4 | 18 | 36 | 38 |
| 4 | 45 | F | 4 | 45 | F | 42 | 3 | 18 | 70 | 67.86 |
| 5 | 40 | F | 5 | 39 | F | 36 | 3 | 16 | 50 | 44 |
| 6 | 48 | F | 6 | 49 | F | 47 | 1.5 | 16 | 80 | 69.77 |
| 7 | 49 | F | 7 | 49 | F | 42 | 7 | 12 | 85 | 61 |
| 8 | 45 | F | 8 | 45 | F | 44 | 1 | 18 | 90 | 74.21 |
| 9 | 41 | F | 9 | 39 | F | 30 | 9 | 11 | 54 | 58 |
| 10 | 50 | F | 10 | 53 | F | 50 | 3 | 12 | 54 | 37 |
| 11 | 49 | F | 11 | 48 | F | 47 | 1 | 14 | 60 | 47 |
| 12 | 52 | M | 12 | 41 | M | 35 | 6 | 18 | 90 | 80.44 |
| Metabolite | rho | p-Value | |
|---|---|---|---|
| Left VLPFC | - | - | - |
| Opercular part of the inferior frontal gyrus CT | Glu/Cr | −0.75 | 0.010 |
| Triangular part of the inferior frontal gyrus CT | Glu/Cr | −0.70 | 0.010 |
| Middle anterior part of the cingulate gyrus CT | Glx/Cr | −0.81 | 0.003 |
| Right thalamus | - | - | - |
| SubCortGrayVol | Glu/Cr | −0.78 | 0.002 |
| Cortical Regions | Controls (Mean mm2 ± SD) | FM Patients (Mean ± SD) | p-Value |
|---|---|---|---|
| Left hemisphere | |||
| Anterior part of the cingulate gyrus and sulcus | 2.78 ± 0.17 | 2.61 ± 0.14 | 0.007 |
| Right hemisphere | |||
| Inferior occipital gyrus and sulcus | 2.66 ± 0.17 | 2.46 ± 0.19 | 0.009 |
| Middle posterior part of the cingulate gyrus and sulcus | 2.68 ± 0.09 | 2.49 ± 0.12 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feraco, P.; Nigro, S.; Passamonti, L.; Grecucci, A.; Caligiuri, M.E.; Gagliardo, C.; Bacci, A. Neurochemical Correlates of Brain Atrophy in Fibromyalgia Syndrome: A Magnetic Resonance Spectroscopy and Cortical Thickness Study. Brain Sci. 2020, 10, 395. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060395
Feraco P, Nigro S, Passamonti L, Grecucci A, Caligiuri ME, Gagliardo C, Bacci A. Neurochemical Correlates of Brain Atrophy in Fibromyalgia Syndrome: A Magnetic Resonance Spectroscopy and Cortical Thickness Study. Brain Sciences. 2020; 10(6):395. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060395
Chicago/Turabian StyleFeraco, Paola, Salvatore Nigro, Luca Passamonti, Alessandro Grecucci, Maria Eugenia Caligiuri, Cesare Gagliardo, and Antonella Bacci. 2020. "Neurochemical Correlates of Brain Atrophy in Fibromyalgia Syndrome: A Magnetic Resonance Spectroscopy and Cortical Thickness Study" Brain Sciences 10, no. 6: 395. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10060395