Treadmill Exercise Buffers Behavioral Alterations Related to Ethanol Binge-Drinking in Adolescent Mice

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
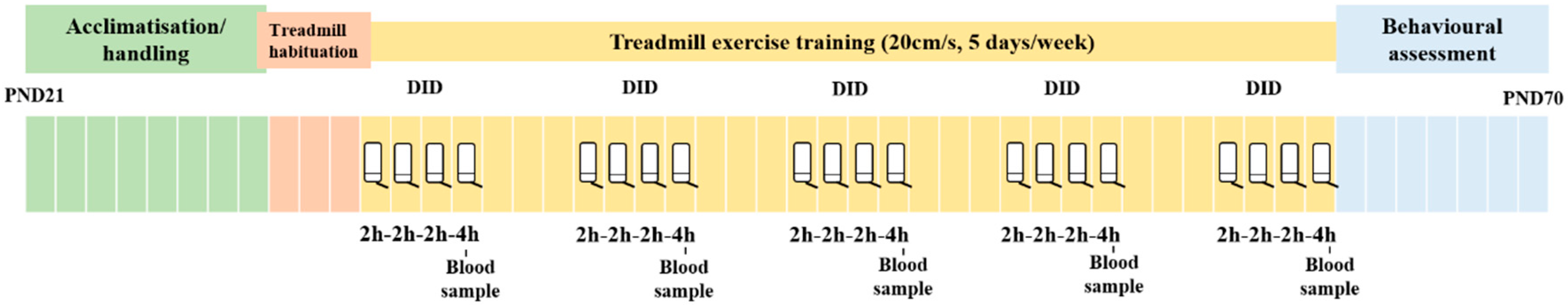
2.2. Procedure
2.2.1. Treadmill Exercise
2.2.2. Drinking in the Dark Procedure (DID)
2.2.3. Blood EtOH Concentration (BEC)
2.2.4. Behavioral Assessment
2.2.5. Elevated Plus Maze
2.2.6. Y Maze
2.2.7. Open Field Test
2.2.8. Nest Building Test
2.2.9. Sucrose Splash Test
2.2.10. Three-Chamber Test
2.3. Data Analysis
3. Results
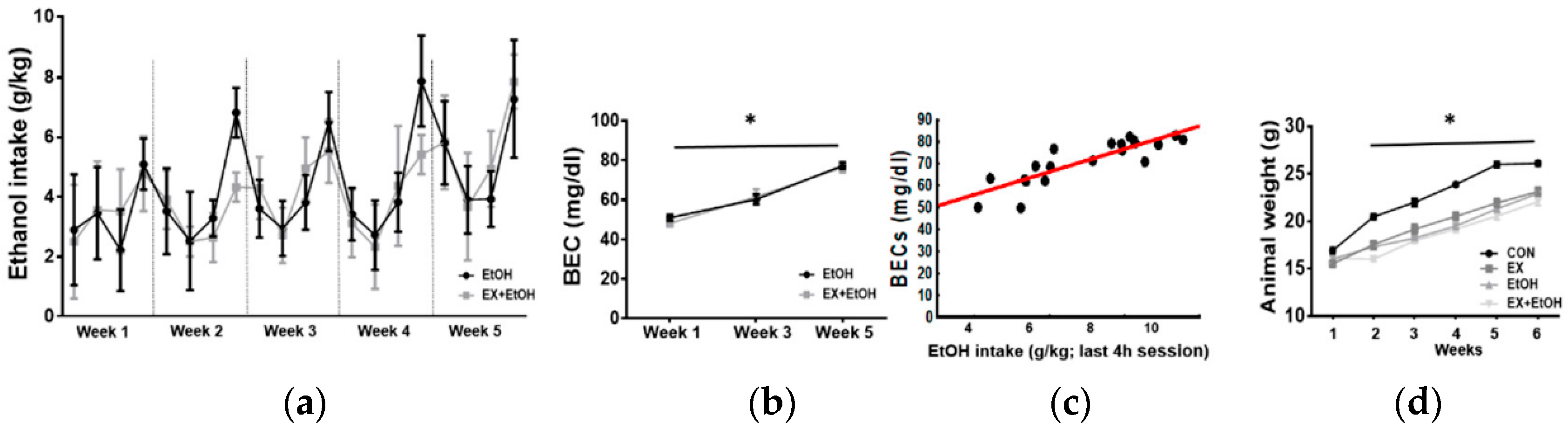
3.1. EtOH Intake and BEC
3.2. Body Weight
3.3. Behavioral Results
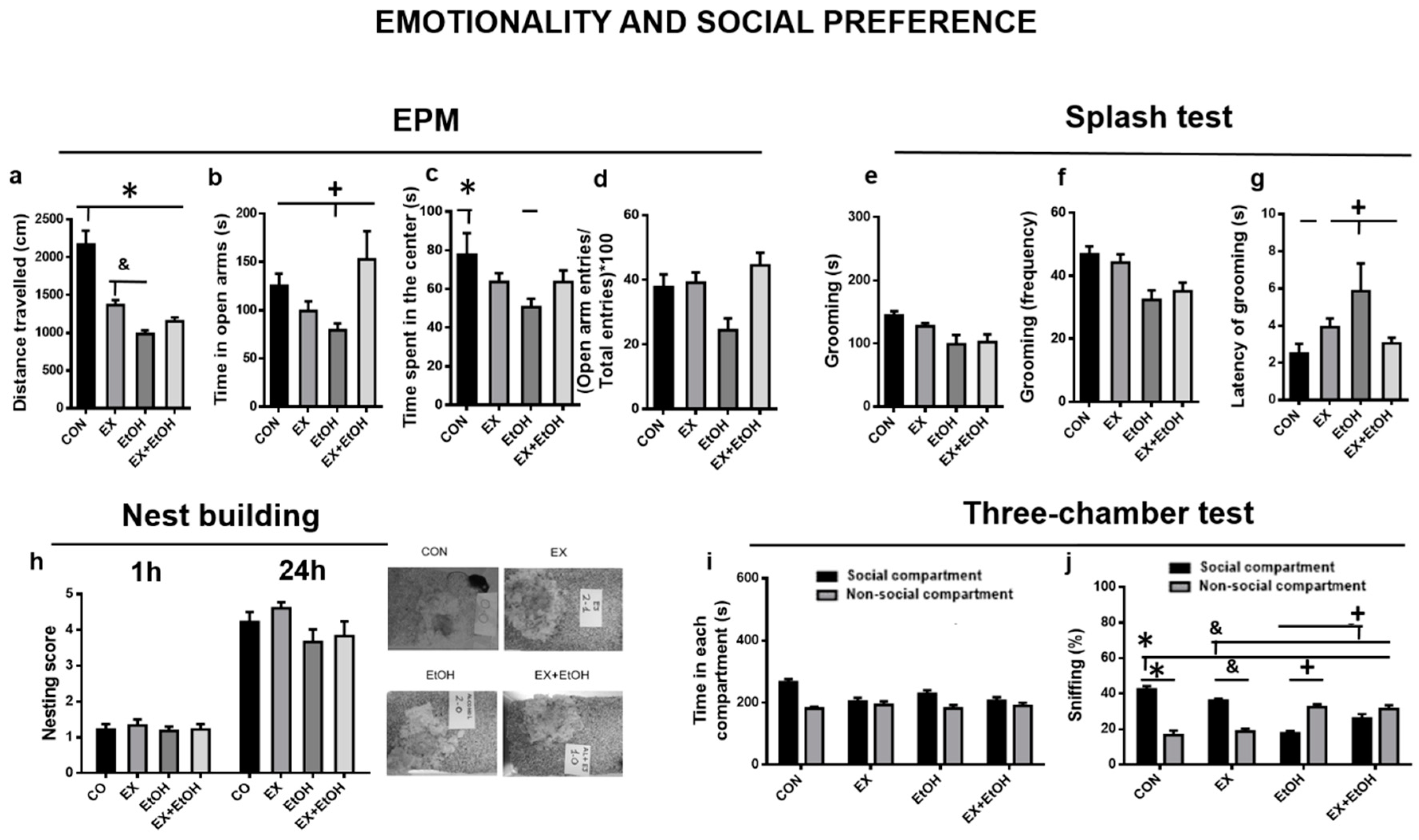
3.3.1. Emotionality and Social Preference
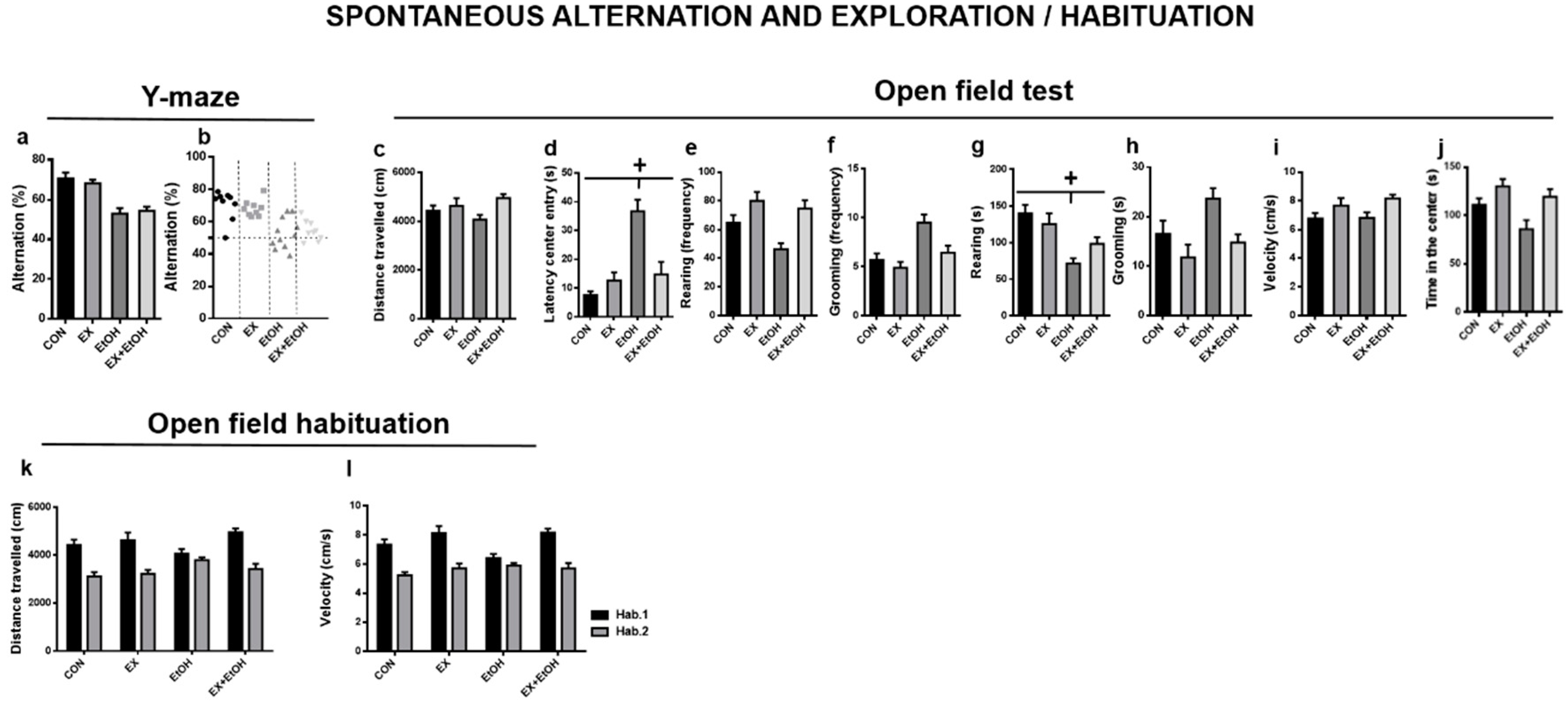
3.3.2. Spontaneous Alternation and Exploration
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kuntsche, E.; Kuntsche, S.; Thrul, J.; Gmel, G. Binge drinking: Health impact, prevalence, correlates and interventions. Psychol. Health 2017, 32, 976–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siqueira, L.; Smith, V.C. Committee on substance abuse binge drinking. Binge drinking. Pediatrics 2015, 136, e718–e726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual, M.; Pla, A.; Miñarro, J.; Guerri, C. Neuroimmune activation and myelin changes in adolescent rats exposed to high-dose alcohol and associated cognitive dysfunction: A review with reference to human adolescent drinking. Alcohol Alcohol. 2014, 49, 187–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squeglia, L.M.; Jacobus, J.; Tapert, S.F. The influence of substance use on adolescent brain development. Clin. EEG Neurosci. 2009, 40, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaudet, G.; Valable, S.; Bourgine, J.; Lelong-Boulouard, V.; Lanfumey, L.; Freret, T.; Boulouard, M.; Paizanis, E. Long-lasting effects of chronic intermittent alcohol exposure in adolescent mice on object recognition and hippocampal neuronal activity. Alcohol Clin. Exp. Res. 2016, 40, 2591–2603. [Google Scholar] [CrossRef]
- Rodd-Henricks, Z.A.; Bell, R.L.; Kuc, K.A.; Murphy, J.M.; McBride, W.J.; Lumeng, L.; Li, T.K. Effects of ethanol exposure on subsequent acquisition and extinction of ethanol self-administration and expression of alcohol-seeking behavior in adult alcohol-preferring (P) rats: I. Periadolescent exposure. Alcohol Clin. Exp. Res. 2002, 26, 1632–1641. [Google Scholar] [CrossRef]
- Rico-Barrio, I.; Peñasco, S.; Puente, N.; Ramos, A.; Fontaine, C.J.; Reguero, L.; Giordano, M.E.; Buceta, I.; Terradillos, I.; Lekunberri, L.; et al. Cognitive and neurobehavioral benefits of an enriched environment on young adult mice after chronic ethanol consumption during adolescence. Addict. Biol. 2018, 24, 969–980. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, E.; de la Fuente, L.; de Amo, E.; Cubero, I. Environmental enrichment during adolescence acts as a protective and therapeutic tool for ethanol binge-drinking, anxiety-like, novelty seeking and compulsive-like behaviors in C57BL/6J mice during adulthood. Front. Behav. Neurosci. 2018, 12, 177. [Google Scholar] [CrossRef]
- Frick, K.M.; Benoit, J.D. Use it or lose it: Environmental enrichment as a means to promote successful cognitive aging. Sci. World J. 2010, 10, 1129–1141. [Google Scholar] [CrossRef] [Green Version]
- Cotman, C.W.; Berchtold, N.C.; Christie, L.A. Exercise builds brain health: Key roles of growth factor cascades and inflammation. Trends Neurosci. 2007, 30, 464–472. [Google Scholar] [CrossRef]
- Kobilo, T.; Liu, Q.R.; Gandhi, K.; Mughai, M.; Shaham, Y.; van Praag, H. Running is the neurogenic and neurotrophic stimulus in environmental enrichment. Learn. Mem. 2011, 18, 605–609. [Google Scholar] [CrossRef] [Green Version]
- Carek, P.J.; Laibstain, S.E.; Carek, S.M. Exercise for the treatment of depression and anxiety. Int. J. Psychiatry Med. 2011, 41, 15–28. [Google Scholar] [CrossRef]
- Hötting, K.; Roder, B. Beneficial effects of physical exercise on neuroplasticity and cognition. Neurosci. Biobehav. Rev. 2013, 37, 2243–2257. [Google Scholar] [CrossRef]
- Knapen, J.; Vancampfort, D.; Moriën, Y.; Marchal, Y. Exercise therapy improves both mental and physical health in patients with major depression. Disabil. Rehab. 2015, 37, 1490–1495. [Google Scholar] [CrossRef]
- Morgan, J.A.; Singhal, G.; Corrigan, F.; Jaehne, E.J.; Jawahar, M.C.; Baune, B.T. The effects of aerobic exercise on depression-like, anxiety-like, and cognition-like behaviours over the healthy adult lifespan of C57BL/6 mice. Behav. Brain Res. 2018, 337, 193–203. [Google Scholar] [CrossRef]
- Manthou, E.; Georgakouli, K.; Fatouros, I.G.; Gianoulakis, C.; Theodorakis, Y.; Jamurtas, A.Z. Role of exercise in the treatment of alcohol use disorders. Biomed. Rep. 2016, 4, 535–545. [Google Scholar] [CrossRef] [Green Version]
- Roessler, K.K.; Bilberg, R.; Søgaard-Nielsen, A.; Jensen, K.; Ekstrøm, C.T.; Sari, S. Exercise as adjunctive treatment for alcohol use disorder: A randomized controlled trial. PLoS ONE 2017, 12, e0186076. [Google Scholar] [CrossRef]
- Leasure, J.L.; Nixon, K. Exercise neuroprotection in a rat model of binge alcohol consumption. Alcohol Clin. Exp. Res. 2010, 34, 404–414. [Google Scholar] [CrossRef] [Green Version]
- Maynard, M.E.; Leasure, J.L. Exercise enhances hippocampal recovery following binge ethanol exposure. PLoS ONE 2013, 8, e76644. [Google Scholar] [CrossRef] [Green Version]
- Herting, M.M.; Chu, X. Exercise, cognition, and the adolescent brain. Birth Defects Res. 2017, 109, 1672–1979. [Google Scholar] [CrossRef]
- Lamarão-Vieira, K.; Pamplona-Santos, D.; Nascimento, P.C.; Corrêa, M.G.; Bittencourt, L.O.; Dos Santos, S.M.; Cartágenes, S.C.; Fernandes, L.M.; Monteiro, M.C.; Maia, C.S.; et al. Physical exercise attenuates oxidative stress and morphofunctional cerebellar damages induced by the ethanol binge drinking paradigm from adolescence to adulthood in rats. Oxid. Med. Cell Longev. 2019, 2019, 6802424. [Google Scholar] [CrossRef]
- Pamplona-Santos, D.; Lamarão-Vieira, K.; Nascimento, P.C.; Bittencourt, L.O.; Corrêa, M.G.; Dos Santos, S.M.; Cartágenes, S.C.; Fernandes, L.M.; Monteiro, M.C.; Maia, C.S.; et al. Aerobic physical exercise as a neuroprotector strategy for ethanol binge-drinking effects in the hippocampus and systemic redox status in rats. Oxid. Med. Cell Longev. 2019, 2019, 2415243. [Google Scholar] [CrossRef]
- Brust, V.; Schindler, P.M.; Lewejohann, L. Lifetime development of behavioural phenotype in the house mouse (Mus musculus). Front. Zool. 2015, 12, S17. [Google Scholar] [CrossRef] [Green Version]
- Taft, R.A.; Davisson, M.; Wiles, M.V. Know thy mouse. Trends Genet. 2006, 22, 649–653. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef]
- Kim, D.M.; Leem, Y.H. Chronic stress-induced memory deficits are reversed by regular exercise via AMPK-mediated BDNF induction. Neuroscience 2016, 324, 271–285. [Google Scholar] [CrossRef]
- Rhodes, J.S.; Best, K.; Belknap, J.K.; Finn, D.A.; Crabbe, J.C. Evaluation of a simple model of ethanol drinking to intoxication in C57BL/6J mice. Physiol. Behav. 2005, 84, 53–63. [Google Scholar] [CrossRef]
- Ladrón de Guevara-Miranda, D.; Millón, C.; Rosell-Valle, C.; Pérez-Fernández, M.; Missiroli, M.; Serrano, A.; Pavón, F.J.; Rodríguez de Fonseca, F.; Martínez-Losa, M.; Álvarez-Dolado, M.; et al. Long-lasting memory deficits in mice withdrawn from cocaine are concomitant with neuroadaptations in hippocampal basal activity, GABAergic interneurons and adult neurogenesis. Dis. Model Mech. 2017, 10, 323–336. [Google Scholar] [CrossRef] [Green Version]
- Deacon, R.M. Assessing nest building in mice. Nat. Protoc. 2006, 1, 1117–1119. [Google Scholar] [CrossRef]
- Moreno-Fernández, R.D.; Pérez-Martin, M.; Castilla-Ortega, E.; Rosell Del Valle, C.; García-Fernández, M.I.; Chun, J.; Estivill-Torrús, G.; Rodríguez de Fonseca, F.; Santín, L.J.; Pedraza, C. maLPA1-null mice as an endophenotype of anxious depression. Transl. Psychiatry 2017, 7, e1077. [Google Scholar] [CrossRef] [Green Version]
- Machado, D.G.; Cunha, M.P.; Neis, V.B.; Balen, G.O.; Colla, A.; Grando, J.; Brocardo, P.S.; Bettio, L.E.; Capra, J.C.; Rodrigues, A.L. Fluoxetine reverses depressive-like behaviors and increases hippocampal acetylcholinesterase activity induced by olfactory bulbectomy. Pharmacol. Biochem. Behav. 2012, 103, 220–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zain, M.A.; Pandy, V.; Majeed, A.B.A.; Wong, W.F.; Mohamed, Z. Chronic restraint stress impairs sociability but not social recognition and spatial memory in C57BL/6J mice. Exp. Anim. 2019, 68, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilpin, N.W.; Karanikas, C.A.; Richardson, H.N. Adolescent binge drinking leads to changes in alcohol drinking, anxiety, and amygdalar corticotropin releasing factor cells in adulthood in male rats. PLoS ONE 2012, 7, e31466. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Coelho, M.A.; Class, M.A.; Szumlinski, K.K. mGlu5-dependent modulation of anxiety during early withdrawal from binge-drinking in adult and adolescent male mice. Drug Alcohol Depend. 2018, 184, 1–11. [Google Scholar] [CrossRef]
- Wiese, J.G.; Shlipak, M.G.; Browner, W.S. The alcohol hangover. Ann. Intern. Med. 2000, 132, 897–902. [Google Scholar] [CrossRef] [Green Version]
- Karadayian, A.G.; Busso, M.J.; Feleder, C.; Cutrera, R.A. Alterations in affective behavior during the time course of alcohol hangover. Behav. Brain Res. 2013, 253, 128–138. [Google Scholar] [CrossRef]
- Doremus, T.L.; Brunell, S.C.; Varlinskaya, E.I.; Spear, L.P. Anxiogenic effects during withdrawal from acute ethanol in adolescent and adult rats. Pharmacol. Biochem. Behav. 2003, 75, 411–418. [Google Scholar] [CrossRef]
- Lal, H.; Prather, P.L.; Rezazadeh, S.M. Anxiogenic behavior in rats during acute and protracted ethanol withdrawal: Reversal by buspirone. Alcohol 1991, 8, 467–471. [Google Scholar] [CrossRef]
- Becker, H.C. Effects of alcohol dependence and withdrawal on stress responsiveness and alcohol consumption. Alcohol Res. Curr. Rev. 2012, 34, 448–458. [Google Scholar]
- Gatch, M.B.; Lal, H. Animal models of the anxiogenic effects of ethanol withdrawal. Drug Dev. Res. 2001, 54, 95–115. [Google Scholar] [CrossRef]
- Cryan, J.F.; Holmes, A. The ascent of mouse: Advances in modelling human depression and anxiety. Nat. Rev. Drug Discov. 2005, 4, 775–790. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, G.D.; Phillips, T.J.; Crabbe, J.C. Effects of acute alcohol withdrawal on nest building in mice selectively bred for alcohol withdrawal severity. Physiol. Behav. 2016, 165, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isingrini, E.; Camus, V.; Le Guisquet, A.M.; Pingaud, M.; Derves, S.; Belzung, C. Association between repeated unpredictable chronic mild stress (UCMS) procedures with a high fat diet: A model of fluoxetine resistance in mice. PLoS ONE 2010, 5, e10404. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Morán, S.; Estanislau, C.; Cañete, T.; Blázquez, G.; Ráez, A.; Tobeña, A.; Fernández-Teruel, A. Relationships of open-field behaviour with anxiety in the elevated zero-maze test: Focus on freezing and grooming. WJNS 2014, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Escorihuela, R.M.; Fernandez-Teruel, A.; Gil, L.; Aguilar, R.; Tobena, A.; Driscoll, P. Inbred Roman high- and low-avoidance rats: Differences in anxiety, novelty-seeking, and shuttle box behaviors. Physiol. Behav. 1999, 67, 19–26. [Google Scholar] [CrossRef]
- Planchez, B.; Surget, A.; Belzung, C. Animal models of major depression: Drawbacks and challenges. J. Neural Transm. 2019, 126, 1383–1408. [Google Scholar] [CrossRef] [Green Version]
- Levy, R.; Dubois, B. Apathy and the functional anatomy of the prefrontal cortex-basal ganglia circuits. Cereb. Cortex 2006, 16, 916–928. [Google Scholar] [CrossRef] [Green Version]
- Cathomas, F.; Hartmann, M.N.; Seifritz, E.; Pryce, C.R.; Kaiser, S. The translational study of apathy-an ecological approach. Front. Behav. Neurosci. 2015, 9, 241. [Google Scholar] [CrossRef]
- Sampedro-Piquero, P.; Ávila-Gámiz, F.; Moreno-Fernández, R.D.; Castilla-Ortega, E.; Santín, L.J. The presence of a social stimulus reduces cocaine-seeking in a place preference conditioning paradigm. J. Psychopharmacol. 2019, 33, 1501–1511. [Google Scholar] [CrossRef]
- Setlalentoa, M.; Ryke, E.; Strydom, H. The influence of binge drinking on social support networks. Soc. Work 2014, 50, 349–370. [Google Scholar] [CrossRef]
- Varlinskaya, E.I.; Kim, E.U.; Spear, L.P. Chronic intermittent ethanol exposure during adolescence: Effects on stress-induced social alterations and social drinking in adulthood. Brain Res. 2017, 1654, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Lynch, W.J.; Piehl, K.B.; Acosta, G.; Peterson, A.B.; Hemby, S.E. Aerobic exercise attenuates reinstatement of cocaine-seeking behavior and associated neuroadaptations in the prefrontal cortex. Biol. Psychiatry 2010, 68, 774–777. [Google Scholar] [CrossRef] [Green Version]
- Morrison, T.R.; Ricci, L.A.; Puckett, A.S.; Joyce, J.; Curran, R.; Davis, C.; Melloni, R.H. Serotonin type-3 receptors differentially modulate anxiety and aggression during withdrawal from adolescent anabolic steroid exposure. Horm. Behav. 2020, 119, 104650. [Google Scholar] [CrossRef]
- Skorzewska, A.; Lehner, A.; Wislowska-Stanek, A.; Krzascik, P.; Ziemba, A.; Plaznik, A. The effect of chronic administration of corticosterone on anxiety- and depression-like behavior and the expression of GABA-A receptor alpha-2 subunits in brain structures of low- and high-anxiety rats. Horm. Behav. 2014, 65, 6–13. [Google Scholar] [CrossRef]
- Zare, Z.; Tehrani, N.; Rezaei, N.; Dana Ghalebarzand, B.; Mohammadi, M. Anxiolytic activity of paraoxon is associated with alterations in rat brain glutamatergic system. Neurotoxicol. Teratol. 2019, 71, 32–40. [Google Scholar] [CrossRef]
- DeBoer, L.B.; Powers, M.B.; Utschig, A.C.; Otto, M.W.; Smits, J.A. Exploring exercise as an avenue for the treatment of anxiety disorders. Expert Rev. Neurother. 2012, 12, 1011–1022. [Google Scholar] [CrossRef] [Green Version]
- Ehringer, M.A.; Hoft, N.R.; Zunhammer, M. Reduced alcohol consumption in mice with access to a running wheel. Alcohol 2009, 43, 443–452. [Google Scholar] [CrossRef]
- Kim, J.W.; Nam, S.M.; Yoo, D.Y.; Jung, H.Y.; Kim, I.Y.; Seong, J.K.; Yoon, Y.S. Comparison of adult hippocampal neurogenesis and susceptibility to treadmill exercise in nine mouse strains. Neural Plast. 2017, 2017, 5863258. [Google Scholar] [CrossRef]
- Llorens-Martín, M.; Torres-Alemán, I.; Trejo, J.L. Exercise modulates insulin-like growth factor 1-dependent and- independent effects on adult hippocampal neurogenesis and behaviour. Mol. Cell Neurosci. 2010, 44, 109–117. [Google Scholar] [CrossRef]
- Hwang, D.S.; Kwak, H.B.; Ko, I.G.; Kim, S.E.; Jin, J.J.; Ji, E.S.; Choi, H.H.; Kwon, O.Y. Treadmill exercise improves memory function depending on circadian rhythm changes in mice. Int. Neurourol. J. 2016, 20, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Roedel, A.; Storch, C.; Holsboer, F.; Ohl, F. Effects of light or dark phase testing on behavioural and cognitive performance in DBA mice. Lab. Anim. 2006, 40, 371–381. [Google Scholar] [CrossRef]
- Eckel-Mahan, K.; Sassone-Corsi, P. Phenotyping circadian rhythms in mice. Curr. Protoc. Mouse Biol. 2015, 5, 271–281. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sampedro-Piquero, P.; Millón, C.; Moreno-Fernández, R.D.; García-Fernández, M.; Diaz-Cabiale, Z.; Santin, L.J. Treadmill Exercise Buffers Behavioral Alterations Related to Ethanol Binge-Drinking in Adolescent Mice. Brain Sci. 2020, 10, 576. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10090576
Sampedro-Piquero P, Millón C, Moreno-Fernández RD, García-Fernández M, Diaz-Cabiale Z, Santin LJ. Treadmill Exercise Buffers Behavioral Alterations Related to Ethanol Binge-Drinking in Adolescent Mice. Brain Sciences. 2020; 10(9):576. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10090576
Chicago/Turabian StyleSampedro-Piquero, Patricia, Carmelo Millón, Román D. Moreno-Fernández, María García-Fernández, Zaida Diaz-Cabiale, and Luis Javier Santin. 2020. "Treadmill Exercise Buffers Behavioral Alterations Related to Ethanol Binge-Drinking in Adolescent Mice" Brain Sciences 10, no. 9: 576. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10090576