
Genetic and Pharmacological Manipulations of Glyoxalase 1 Mediate Ethanol Withdrawal Seizure Susceptibility in Mice

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Husbandry
2.2. Handling-Induced Convulsions
2.3. Drugs
2.4. Experiment 1: Acute Ethanol Withdrawal in Glo1 Transgenic Mice on a B6 Background
2.5. Experiment 2: Acute Ethanol Withdrawal in Glo1 Transgenic Mice on an FVB Background
2.6. Experiment 3: GLO1 Inhibitor Effects on Chronic Ethanol Withdrawal
2.7. Statistical Analyses
3. Results
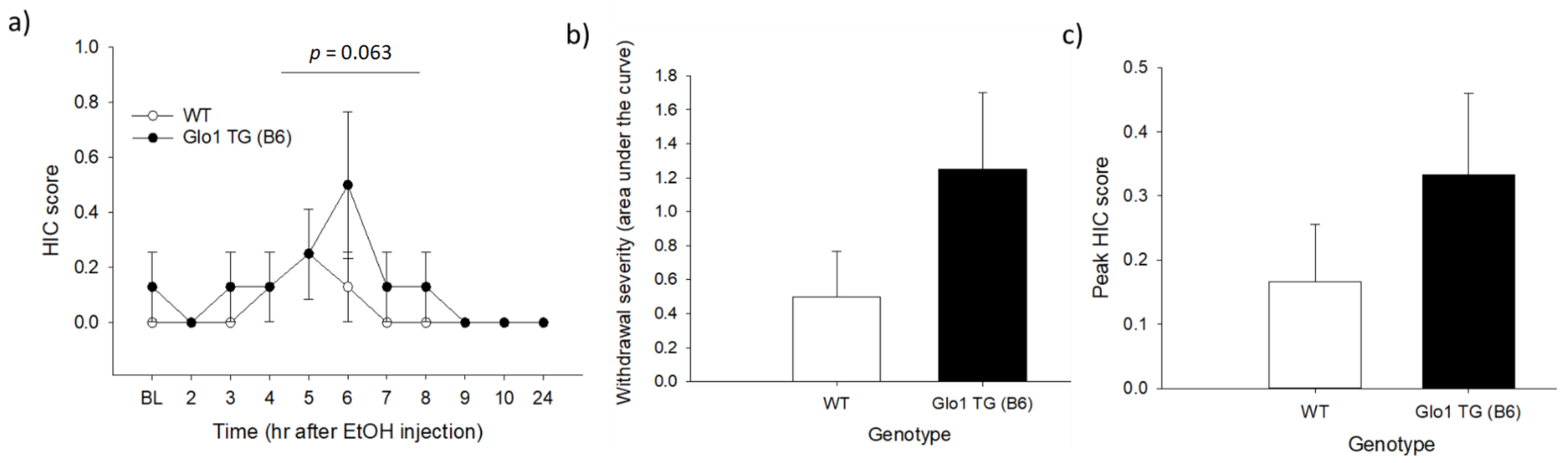
3.1. Experiment 1: Acute Ethanol Withdrawal in Glo1 Transgenic Mice on a B6 Background
3.2. Experiment 2: Acute Ethanol Withdrawal in Glo1 Transgenic Mice on an FVB Background
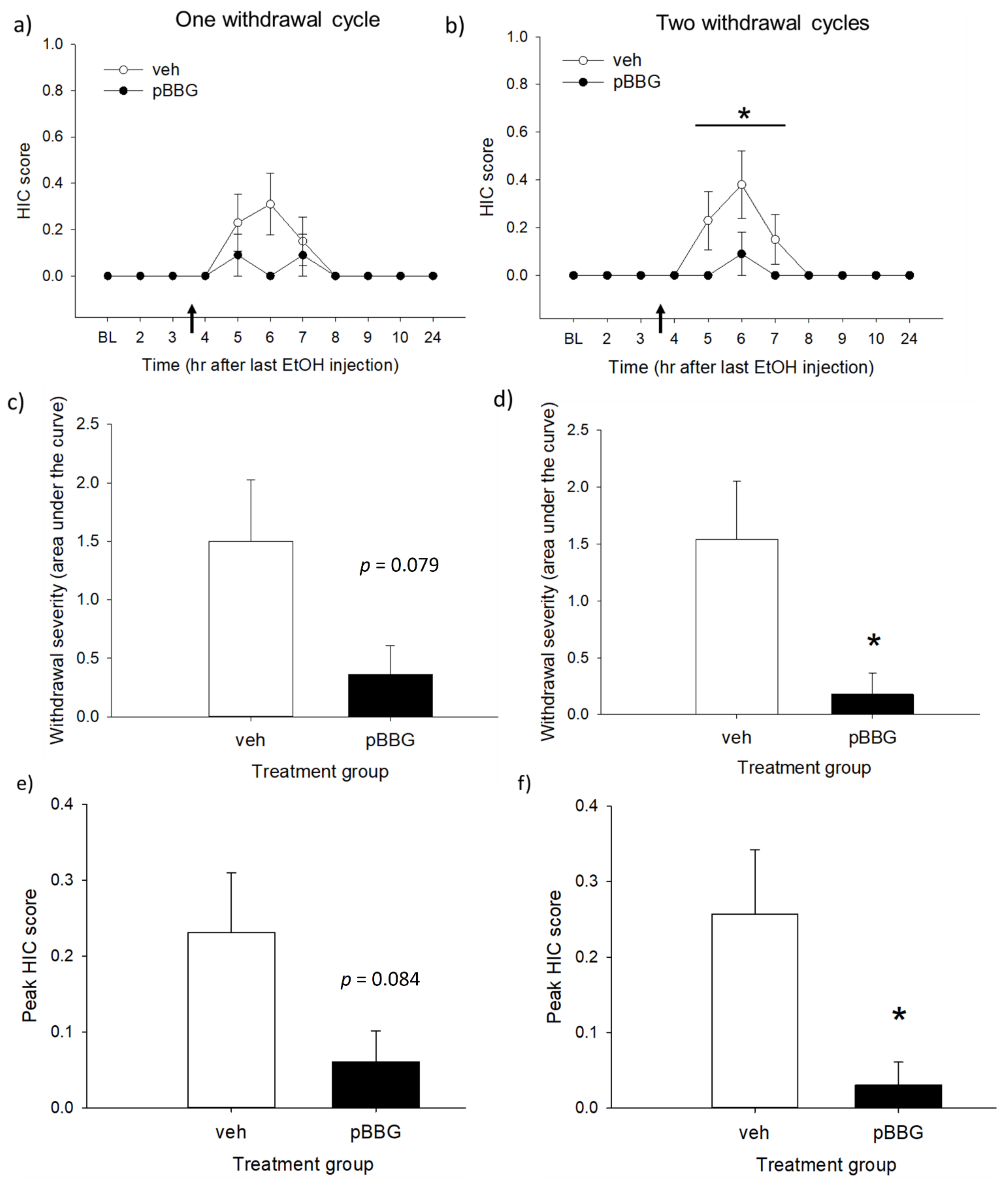
3.3. Experiment 3: GLO1 Inhibitor Effects on Chronic Ethanol Withdrawal
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; (DSM-5); American Psychiatric Association: Washington, DC, USA, 2013; p. 5. [Google Scholar]
- Becker, H.C. Alcohol Dependence, Withdrawal, and Relapse. Neurobiol. Alcohol Depend. 2014, 31, 377–410. [Google Scholar] [CrossRef] [Green Version]
- Breese, G.R.; Sinha, R.; Heilig, M. Chronic alcohol neuroadaptation and stress contribute to susceptibility for alcohol craving and relapse. Pharm. Ther. 2011, 129, 149–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilig, M.; Egli, M.; Crabbe, J.C.; Becker, H.C. Acute withdrawal, protracted abstinence and negative affect in alcoholism: Are they linked? Addict. Biol. 2010, 15, 169–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesse, S.; Bråthen, G.; Ferrara, M.; Keindl, M.; Ben-Menachem, E.; Tanasescu, R.; Brodtkorb, E.; Hillbom, M.; Leone, M.; Ludolph, A. Alcohol withdrawal syndrome: Mechanisms, manifestations, and management. Acta Neurol. Scand. 2017, 135, 4–16. [Google Scholar] [CrossRef]
- McKeon, A.; Frye, M.A.; Delanty, N. The alcohol withdrawal syndrome. J. Neurol. Neurosurg. Psychiatry 2008, 79, 854–862. [Google Scholar] [CrossRef] [Green Version]
- Saitz, R. Introduction to alcohol withdrawal. Alcohol Health Res. World 1998, 22, 5–12. [Google Scholar]
- Schmidt, L.; Sander, T. Genetics of alcohol withdrawal. Eur. Psychiatry 2000, 15, 135–139. [Google Scholar] [CrossRef]
- Kendler, K.S.; Aggen, S.H.; Prescott, C.A.; Crabbe, J.; Neale, M.C. Evidence for multiple genetic factors underlying the DSM-IV criteria for alcohol dependence. Mol. Psychiatry 2011, 17, 1306–1315. [Google Scholar] [CrossRef] [Green Version]
- Kosobud, A.; Crabbe, J.C. Ethanol withdrawal in mice bred to be genetically prone or resistant to ethanol with-drawal seizures. J. Pharmacol. Exp. Ther. 1986, 238, 170–177. [Google Scholar]
- Metten, P.; Crabbe, J.C. Alcohol withdrawal severity in inbred mouse (Mus musculus) strains. Behav. Neurosci. 2005, 119, 911–925. [Google Scholar] [CrossRef]
- Fehr, C.; Shirley, R.L.; Metten, P.; Kosobud, A.E.K.; Belknap, J.K.; Crabbe, J.C.; Buck, K.J. Potential pleiotropic effects of Mpdz on vulnerability to seizures. GenesBrain Behav. 2004, 3, 8–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, L.L.; Davis, S.J.; Yen, R.C.; Ordemann, G.J.; Nordquist, S.K.; Bannai, D.; Pierce, J.T. Behavioral deficits following withdrawal from chronic ethanol are influenced by SLO channel function in caenorhabditis elegans. Genetics 2017, 206, 1445–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, N.; Helton, S.G.; Schwandt, M.; Zhu, X.; Momenan, R.; Lohoff, F.W. Genetic Variation in the vesicular monoamine transporter 1 (VMAT1/SLC18A1) gene and alcohol withdrawal severity. Alcohol. Clin. Exp. Res. 2016, 40, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Grzywacz, A.; Jasiewicz, A.; Małecka, I.; Suchanecka, A.; Grochans, E.; Karakiewicz, B.; Samochowiec, A.; Bieńkowski, P.; Samochowiec, J. Influence of DRD2 and ANKK1 polymorphisms on the manifestation of withdrawal syndrome symptoms in alcohol addiction. Pharm. Rep. PR 2012, 64, 1126–1134. [Google Scholar] [CrossRef]
- Huang, M.-C.; Schwandt, M.L.; Chester, J.A.; Kirchhoff, A.M.; Kao, C.-F.; Liang, T.; Tapocik, J.D.; Ramchandani, V.A.; George, D.T.; Hodgkinson, C.A.; et al. FKBP5 moderates alcohol withdrawal severity: Human genetic association and functional validation in knockout mice. Neuropsychopharmacology 2014, 39, 2029–2038. [Google Scholar] [CrossRef] [Green Version]
- Sander, T.; Harms, H.; Podschus, J.; Finckh, U.; Nickel, B.; Rolfs, A.; Rommelspacher, H.; Schmidt, L.G. Allelic association of a dopamine transporter gene polymorphism in alcohol dependence with withdrawal seizures or delirium. Biol. Psychiatry 1997, 41, 299–304. [Google Scholar] [CrossRef]
- Smith, A.H.; Ovesen, P.L.; Skeldal, S.; Yeo, S.; Jensen, K.P.; Olsen, D.; DiazGranados, N.; Zhao, H.; Farrer, L.A.; Goldman, D.; et al. Risk locus identification ties alcohol withdrawal symptoms to SORCS2. Alcohol. Clin. Exp. Res. 2018, 42, 2337–2348. [Google Scholar] [CrossRef]
- Crabbe, J.C. Provisional mapping of quantitative trait loci for chronic ethanol withdrawal severity in BXD recombi-nant inbred mice. J. Pharmacol. Exp. Ther. 1998, 286, 263–271. [Google Scholar]
- Metten, P.; Crabbe, J.C. Common genetic determinants of severity of acute withdrawal from ethanol, pentobarbital and diazepam in inbred mice. Behav. Pharm. 1994, 5, 533–547. [Google Scholar] [CrossRef] [Green Version]
- Metten, P.; Sorensen, M.L.; Cameron, A.J.; Yu, C.-H.; Crabbe, J.C. Withdrawal severity after chronic intermittent ethanol in inbred mouse strains. Alcohol. Clin. Exp. Res. 2010, 34, 1552–1564. [Google Scholar] [CrossRef] [Green Version]
- Crabbe, J.C.; Kosobud, A.; Young, E.R. Genetic selection for ethanol withdrawal severity: Differences in replicate mouse lines. Life Sci. 1983, 33, 955–962. [Google Scholar] [CrossRef]
- Distler, M.G.; Plant, L.D.; Sokoloff, G.; Hawk, A.J.; Aneas, I.; Wuenschell, G.E.; Termini, J.; Meredith, S.C.; Nobrega, M.A.; Palmer, A.A. Glyoxalase 1 increases anxiety by reducing GABAA receptor agonist methylglyoxal. J. Clin. Investig. 2012, 122, 2306–2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurray, K.M.J.; Du, X.; Brownlee, M.; Palmer, A.A. Neuronal overexpression of Glo1 or amygdalar microinjection of methylglyoxal is sufficient to regulate anxiety-like behavior in mice. Behav. Brain Res. 2016, 301, 119–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurray, K.M.J.; Ramaker, M.J.; Barkley-Levenson, A.M.; Sidhu, P.S.; Elkin, P.; Reddy, M.K.; Guthrie, M.L.; Cook, J.M.; Rawal, V.H.; Arnold, L.A.; et al. Identification of a novel, fast-acting GABAergic antidepressant. Mol. Psychiatry 2018, 23, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Perez, C.; Barkley-Levenson, A.M.; Dick, B.L.; Glatt, P.F.; Martinez, Y.; Siegel, D.; Momper, J.D.; Palmer, A.A.; Cohen, S.M. Metal-binding pharmacophore library yields the discovery of a Glyoxalase 1 inhibitor. J. Med. Chem. 2019, 62, 1609–1625. [Google Scholar] [CrossRef]
- McMurray, K.M.J.; Sidhu, P.S.; Cook, J.M.; Arnold, L.A.; Palmer, A.A. Genetic and pharmacological manipulation of glyoxalase 1 regulates voluntary ethanol consumption in mice. Addict. Biol. 2015, 22, 381–389. [Google Scholar] [CrossRef] [Green Version]
- De Guglielmo, G.; Conlisk, D.E.; Barkley-Levenson, A.M.; Palmer, A.A.; George, O. Inhibition of Glyoxalase 1 reduces alcohol self-administration in dependent and nondependent rats. Pharm. Biochem. Behav. 2018, 167, 36–41. [Google Scholar] [CrossRef]
- Distler, M.G.; Gorfinkle, N.; Papale, L.A.; Wuenschell, G.E.; Termini, J.; Escayg, A.; Winawer, M.R.; Palmer, A.A. Glyoxalase 1 and its substrate methylglyoxal are novel regulators of seizure susceptibility. Epilepsia 2013, 54, 649–657. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Porcu, P.; Werner, D.F.; Matthews, D.B.; Diaz-Granados, J.L.; Helfand, R.S.; Morrow, A.L. The role of GABAA receptors in the acute and chronic effects of ethanol: A decade of progress. Psychopharmacology 2009, 205, 529–564. [Google Scholar] [CrossRef] [Green Version]
- Rogawski, M.A. Update on the neurobiology of alcohol withdrawal seizures. Epilepsy Curr. 2005, 5, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Sachdeva, A.; Choudhary, M.; Chandra, M. Alcohol withdrawal syndrome: Benzodiazepines and beyond. J. Clin. Diagn. Res. 2015, 9, VE01–VE07. [Google Scholar] [CrossRef] [PubMed]
- Crabbe, J.C.; Merrill, C.D.; Belknap, J.K. Effects of convulsants on handling-induced convulsions in mice selected for ethanol withdrawal severity. Brain Res. 1991, 550, 1–6. [Google Scholar] [CrossRef]
- Goldstein, D.B.; Pal, N. Alcohol dependence produced in mice by inhalation of ethanol: Grading the withdrawal reaction. Science 1971, 172, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.M.; Groot, J.A.; Curtis, D.C.; Allison, C.L.; Marquardt, P.C.; Holmes, A.N.; Edwards, D.S.; Trotter, D.R.M.; Syapin, P.J.; Finn, D.A.; et al. Effective Reduction of acute ethanol withdrawal by the tetracycline derivative, tigecycline, in female and male DBA/2J mice. Alcohol. Clin. Exp. Res. 2016, 40, 2499–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nipper, M.A.; Jensen, J.P.; Helms, M.L.; Ford, M.M.; Crabbe, J.C.; Rossi, D.J.; Finn, D.A. Genotype differences in sensitivity to the anticonvulsant effect of the synthetic neurosteroid ganaxolone during chronic ethanol withdrawal. Neuroscience 2019, 397, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Perez, E.; De Biasi, M. Assessment of affective and somatic signs of ethanol withdrawal in C57BL/6J mice using a short-term ethanol treatment. Alcohol 2015, 49, 237–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homanics, G.E.; Le, N.Q.; Kist, F.; Mihalek, R.; Hart, A.R.; Quinlan, J.J. Ethanol tolerance and withdrawal responses in GABAA receptor alpha 6 subunit null allele mice and in inbred C57BL/6J and strain 129/SvJ mice. Alcohol. Clin. Exp. Res. 1998, 22, 259–265. [Google Scholar] [CrossRef]
- Gorin-Meyer, R.E.; Wiren, K.M.; Tanchuck, M.A.; Long, S.L.; Yoneyama, N.; Finn, D.A. Sex differences in the effect of finasteride on acute ethanol withdrawal severity in C57BL/6J and DBA/2J mice. Neuroscience 2007, 146, 1302–1315. [Google Scholar] [CrossRef] [Green Version]
- Gililland-Kaufman, K.R.; Finn, D.A. The Impact of gonadectomy and adrenalectomy on acute withdrawal severity in male and female C57BL/6J and DBA/2J mice following a single high dose of ethanol. Alcohol. Clin. Exp. Res. 2007, 31, 1846–1857. [Google Scholar] [CrossRef] [Green Version]
- Becker, H.C.; Hale, R.L. Repeated episodes of ethanol withdrawal potentiate the severity of subsequent withdrawal seizures: An animal model of alcohol withdrawal “kindling”. Alcohol. Clin. Exp. Res. 1993, 17, 94–98. [Google Scholar] [CrossRef]
- Booth, B.M.; Blow, F.C. The kindling hypothesis: Further evidence from a U.S. national study of alcoholic men*. Alcohol Alcohol. 1993, 28, 593–598. [Google Scholar] [CrossRef] [PubMed]
- McCown, T.J.; Breese, G.R. Multiple withdrawals from chronic ethanol “kindles” inferior collicular seizure activity: Evidence for kindling of seizures associated with alcoholism. Alcohol. Clin. Exp. Res. 1990, 14, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Becker, H.C.; Mulholland, P.J. Neurochemical mechanisms of alcohol withdrawal. Interv. Neuroradiol. 2014, 125, 133–156. [Google Scholar] [CrossRef]
- Lovinger, D.M.; Roberto, M. Synaptic effects induced by alcohol. Curr. Top. Behav. Neurosci. 2010, 13, 31–86. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.W.; Spigelman, I. GABAA receptor plasticity in alcohol withdrawal. In Jasper’s Basic Mechanisms of the Epilepsies, 4th ed.; Noebels, J.L., Avoli, M., Rogawski, M.A., Olsen, R.W., Delgado-Escueta, A.V., Eds.; National Center for Biotechnology Information: Bethesda, MD, USA, 2012. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK98172/ (accessed on 14 November 2020).



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barkley-Levenson, A.M.; Lee, A.; Palmer, A.A. Genetic and Pharmacological Manipulations of Glyoxalase 1 Mediate Ethanol Withdrawal Seizure Susceptibility in Mice. Brain Sci. 2021, 11, 127. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11010127
Barkley-Levenson AM, Lee A, Palmer AA. Genetic and Pharmacological Manipulations of Glyoxalase 1 Mediate Ethanol Withdrawal Seizure Susceptibility in Mice. Brain Sciences. 2021; 11(1):127. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11010127
Chicago/Turabian StyleBarkley-Levenson, Amanda M., Amy Lee, and Abraham A. Palmer. 2021. "Genetic and Pharmacological Manipulations of Glyoxalase 1 Mediate Ethanol Withdrawal Seizure Susceptibility in Mice" Brain Sciences 11, no. 1: 127. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11010127