
Fear Extinction-Based Inter-Individual and Sex Differences in Pain-Related Vocalizations and Anxiety-like Behaviors but Not Nocifensive Reflexes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
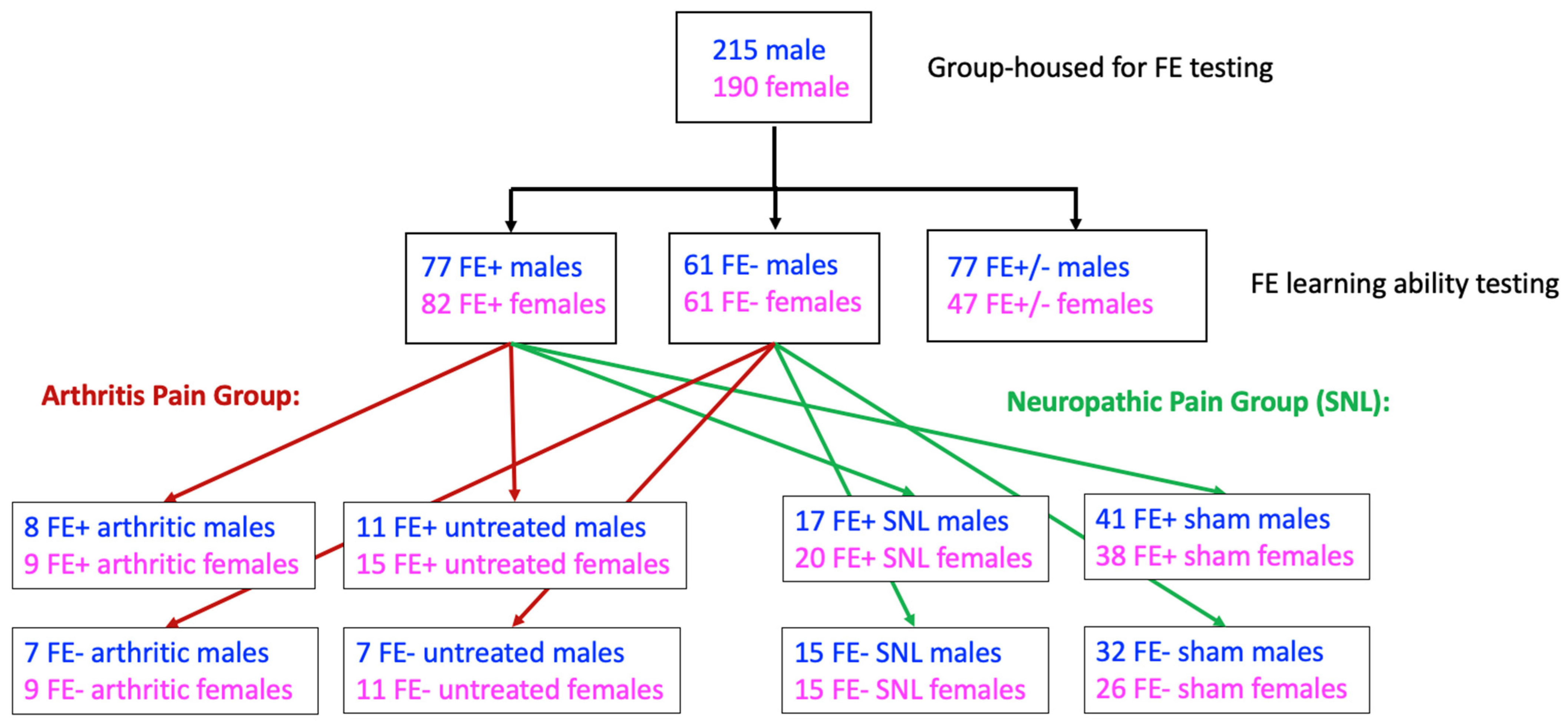
2.2. Experimental Protocol
2.3. Arthritis Pain Model
2.4. Neuropathic Pain Model
2.5. Behaviors
2.5.1. Fear Conditioning and Extinction
2.5.2. Mechanosensitivity
2.5.3. Emotional Responses
2.5.4. Anxiety-Like Behavior
2.6. Statistical Analysis
3. Results
3.1. Inter-Individual and Sex Differences in FE Learning Ability of Naïve Male and Female Rats
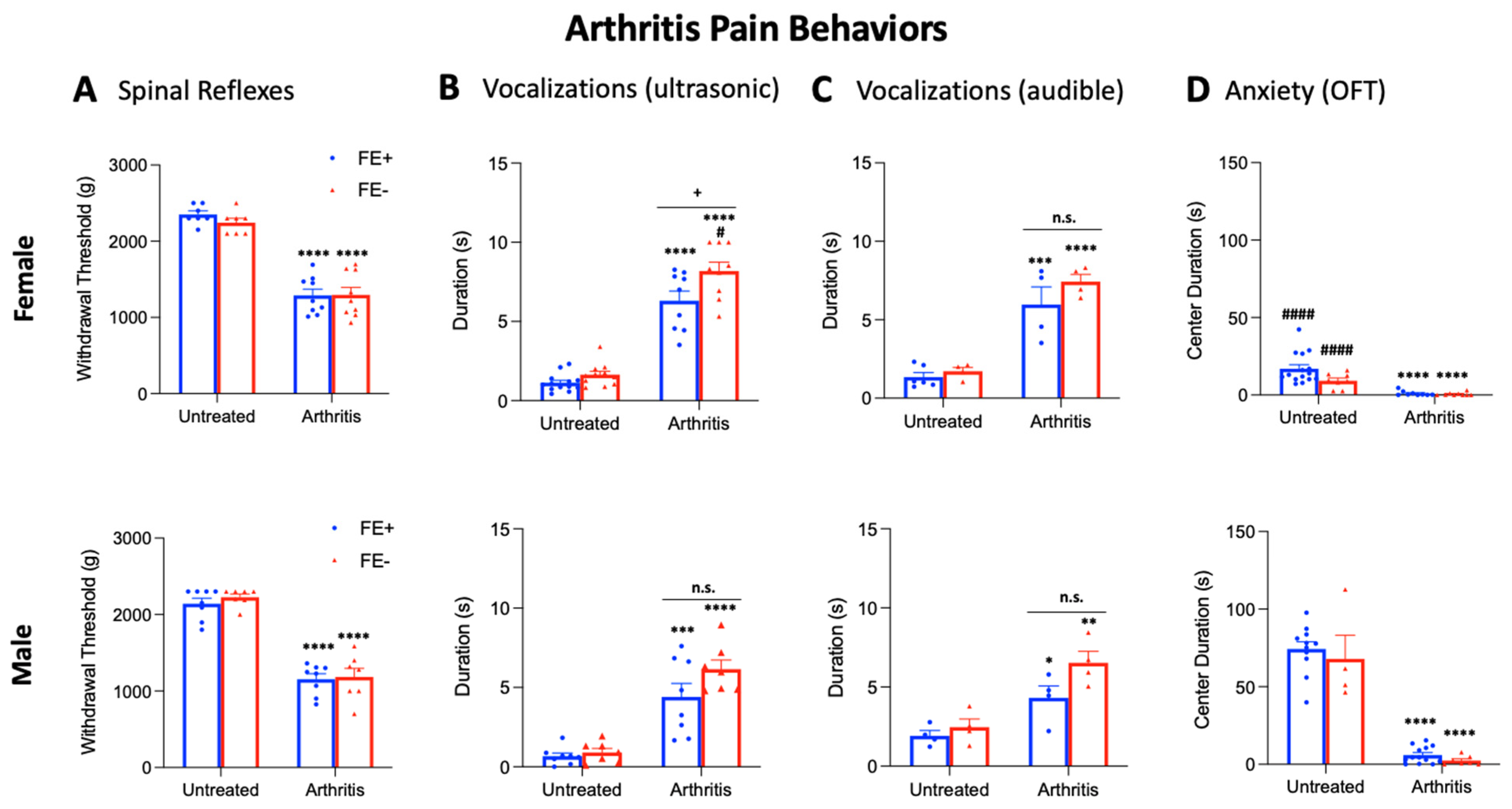
3.2. Inter-Individual and Sex Differences in Arthritis Pain-Related Behaviors of FE+ and FE− Rats
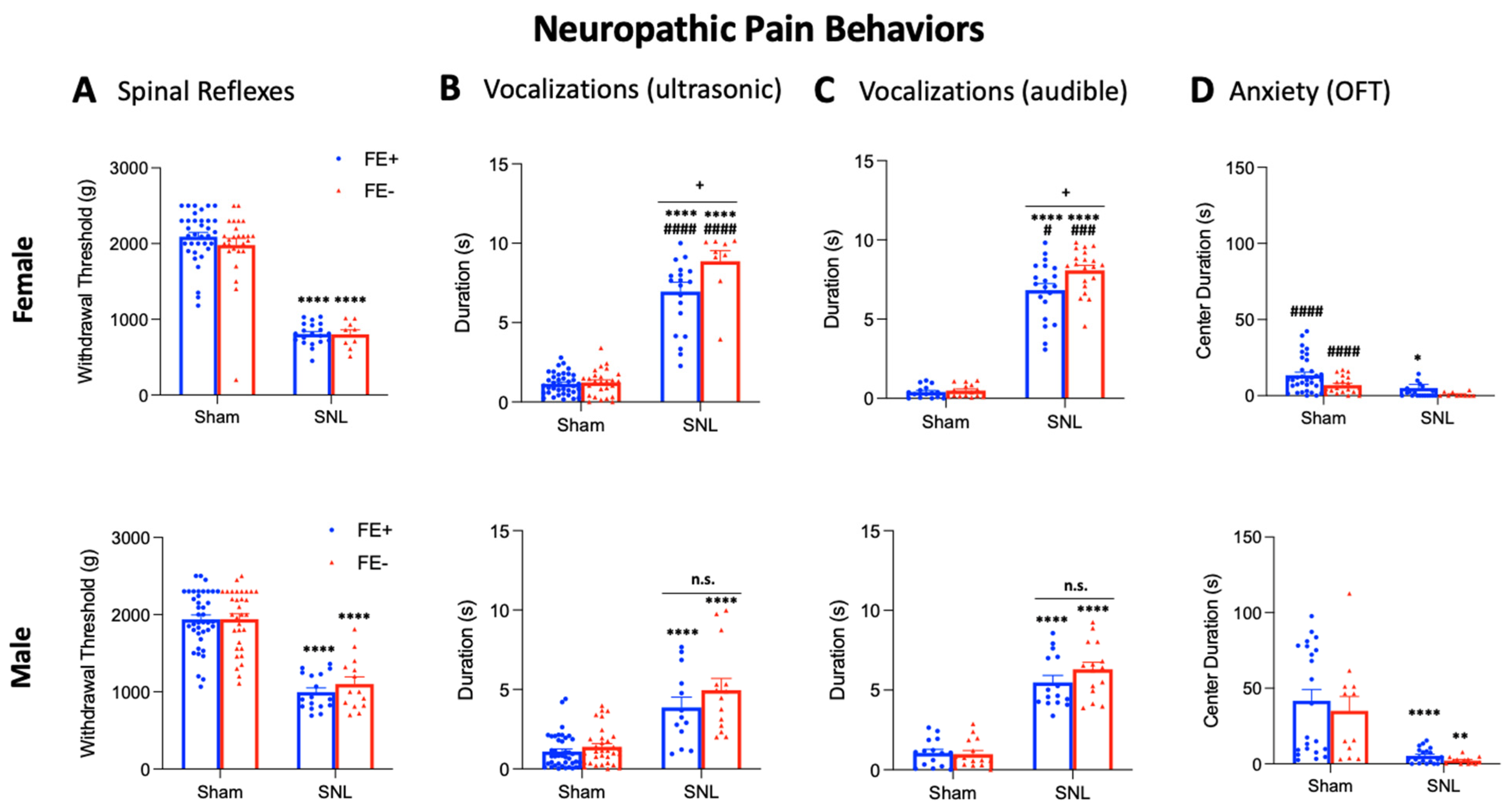
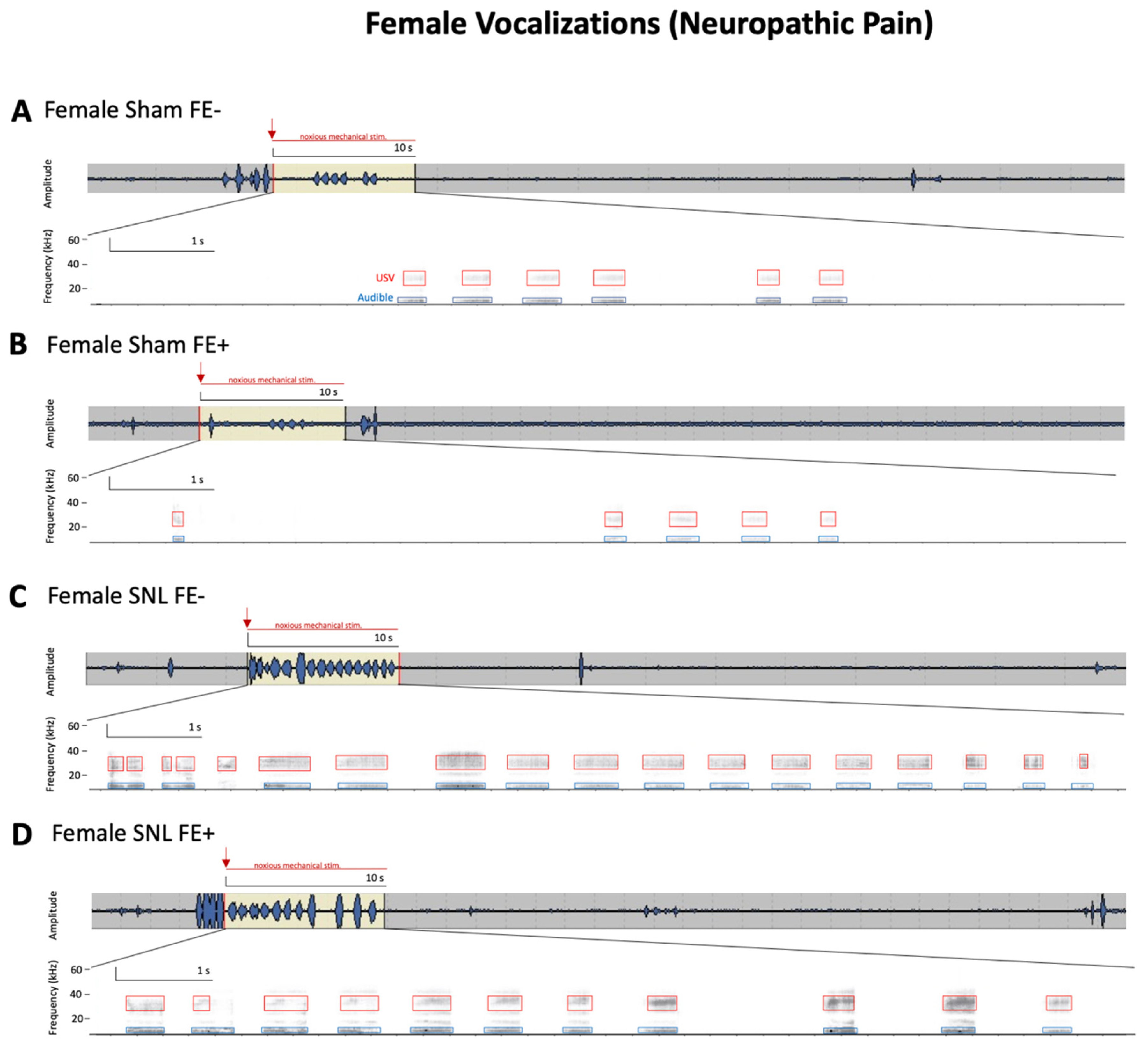
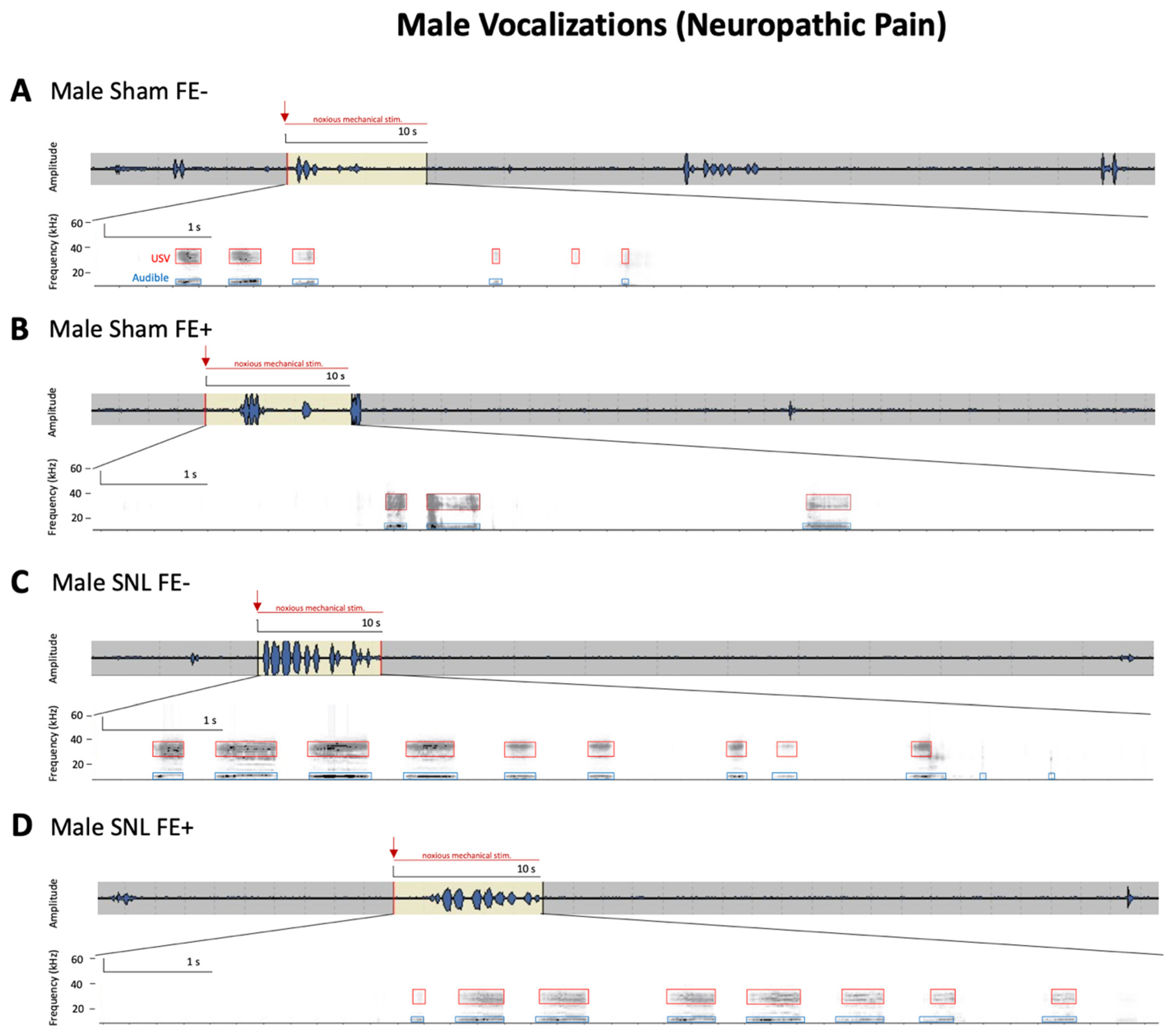
3.3. Inter-Individual and Sex Differences in Neuropathic Pain-Related Behaviors of FE+ and FE− Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Burke, N.N.; Coppinger, J.; Deaver, D.R.; Roche, M.; Finn, D.P.; Kelly, J. Sex differences and similarities in depressive- and anxiety-like behaviour in the Wistar-Kyoto rat. Physiol. Behav. 2016, 167, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Scira, J.; Donaldson, S.; Kajiji, N.; Dash, G.H.; Donaldson, S.T. Sex and Trait Anxiety Differences in Psychological Stress are Modified by Environment. Neuroscience 2018, 383, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Scholl, J.L.; Afzal, A.; Fox, L.C.; Watt, M.J.; Forster, G.L. Sex differences in anxiety-like behaviors in rats. Physiol. Behav. 2019, 211, 112670–112678. [Google Scholar] [CrossRef] [PubMed]
- Coghill, R.C.; McHaffie, J.G.; Yen, Y.-F. Neural correlates of interindividual differences in the subjective experience of Pain. Proc. Natl. Acad. Sci. USA 2003, 100, 8538–8542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fillingim, R.B. Individual Differences in Pain: Understanding the Mosaic that Makes Pain Personal. Pain 2017, 158 (Suppl. 1), S11–S18. [Google Scholar] [CrossRef] [Green Version]
- Sorge, R.E.; Totsch, S.K. Sex Differences in Pain. J. Neurosci. Res. 2017, 95, 1271–1281. [Google Scholar] [CrossRef]
- Moriarty, O.; McGuire, B.E.; Finn, D.P. The effect of pain on cognitive function: A review of clinical and preclinical research. Prog. Neurobiol. 2011, 93, 385–404. [Google Scholar] [CrossRef] [Green Version]
- Veinante, P.; Yalcin, I.; Barrot, M. The amygdala between sensation and affect: A role in Pain. J. Mol. Psychiatry 2013, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Vachon-Presseau, E.; Centeno, M.V.; Ren, W.; Berger, S.E.; Tétreault, P.; Ghantous, M.; Baria, A.; Farmer, M.; Baliki, M.N.; Schnitzer, T.J.; et al. The Emotional Brain as a Predictor and Amplifier of Chronic Pain. J. Dent. Res. 2016, 95, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Neugebauer, V. Amygdala physiology in Pain. Handb. Behav. Neurosci. 2020, 26, 101–113. [Google Scholar]
- Elman, I.; Borsook, D. Threat Response System: Parallel Brain Processes in Pain vis-à-vis Fear and Anxiety. Front. Psychiatry 2018, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- Palmiter, R.D. The parabrachial nucleus: CGRP neurons function as a general alarm. Trends Neurosci. 2018, 41, 280–293. [Google Scholar] [CrossRef] [PubMed]
- Marin, M.F.; Milad, M.R. Extending the Examination of the Fear Extinction Network Beyond Anxiety and Fear-Based Disorders: Insight into Autism Spectrum Disorder. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2016, 1, 302–304. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, N.C.R.; Strong, D.; Abrantes, A.; Garnaat, S.; Cerny, A.; O’Connell, C.; Fadok, R.; Spofford, C.; Rasmussen, S.A.; Milad, M.R.; et al. Extinction retention and fear renewal in a lifetime obsessive–compulsive disorder sample. Behav. Brain Res. 2015, 280, 72–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milad, M.R.; Quirk, G.J. Fear Extinction as a Model for Translational Neuroscience: Ten Years of Progress. Annu. Rev. Psychol. 2012, 63, 129–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, G.; Scott, E.; Graham, B.M.; Richardson, R. Individual differences in fear extinction and anxiety-like behavior. Learn. Mem. 2017, 24, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Lonsdorf, T.B.; Merz, C.J. More than just noise: Inter-individual differences in fear acquisition, extinction and return of fear in humans—Biological, experiential, temperamental factors, and methodological pitfalls. Neurosci. Biobehav. Rev. 2017, 80, 703–728. [Google Scholar] [CrossRef]
- Hånell, A.; Marklund, N. Structured evaluation of rodent behavioral tests used in drug discovery research. Front. Behav. Neurosci. 2014, 8, 252. [Google Scholar] [CrossRef] [Green Version]
- Portfors, C.V. Types and functions of ultrasonic vocalizations in laboratory rats and mice. J. Am. Assoc. Lab. Anim. Sci. 2007, 46, 28–34. [Google Scholar] [PubMed]
- Han, J.S.; Bird, G.C.; Li, W.; Jones, J.; Neugebauer, V. Computerized analysis of audible and ultrasonic vocalizations of rats as a standardized measure of pain-related behavior. J. Neurosci. Methods 2005, 141, 261–269. [Google Scholar] [CrossRef]
- Neugebauer, V.; Han, J.S.; Adwanikar, H.; Fu, Y.; Ji, G. Techniques for assessing knee joint pain in arthritis. Mol. Pain 2007, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Piel, M.J.; Kroin, J.S.; Wijnen AJ van Kc, R.; Im, H.-J. Pain assessment in animal models of osteoarthritis. Gene 2014, 537, 184–188. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Thompson, J.; Ji, G.; Ganapathy, V.; Neugebauer, V. Monomethyl fumarate (MMF) inhibits pain behaviors and amygdala activity in a rat arthritis model. Pain 2017, 158, 2376–2385. [Google Scholar] [CrossRef]
- Han, J.S.; Neugebauer, V. mGluR1 and mGluR5 antagonists in the amygdala inhibit different components of audible and ultrasonic vocalizations in a model of arthritic Pain. Pain 2005, 113, 211–222. [Google Scholar] [CrossRef]
- Kurejova, M.; Nattenmüller, U.; Hildebrandt, U.; Selvaraj, D.; Stösser, S.; Kuner, R. An improved behavioural assay demonstrates that ultrasound vocalizations constitute a reliable indicator of chronic cancer pain and neuropathic Pain. Mol. Pain 2010, 6, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevcik, M.A.; Jonas, B.M.; Lindsay, T.H.; Halvorson, K.G.; Ghilardi, J.R.; Kuskowski, M.A.; Mukherjee, P.; Maggio, J.E.; Mantyh, P.W. Endogenous Opioids Inhibit Early Stage Pancreatic Pain in a Mouse Model of Pancreatic Cancer. Gastroenterology 2006, 131, 900–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgdorf, J.S.; Ghoreishi-Haack, N.; Cearley, C.N.; Kroes, R.A.; Moskal, J.R. Rat ultrasonic vocalizations as a measure of the emotional component of chronic Pain. Neuroreport 2019, 30, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.M.; Yakhnitsa, V.; Ji, G.; Neugebauer, V. Small conductance calcium activated potassium (SK) channel dependent and independent effects of riluzole on neuropathic pain-related amygdala activity and behaviors in rats. Neuropharmacology 2018, 138, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Zhang, W.; Mahimainathan, L.; Narasimhan, M.; Kiritoshi, T.; Fan, X.; Wang, J.; Green, T.A.; Neugebauer, V. 5-HT2C Receptor Knockdown in the Amygdala Inhibits Neuropathic-Pain-Related Plasticity and Behaviors. J. Neurosci. 2017, 37, 1378–1393. [Google Scholar] [CrossRef] [Green Version]
- Abraham, A.D.; Leung, E.J.Y.; Wong, B.A.; Rivera, Z.M.G.; Kruse, L.C.; Clark, J.J.; Land, B.B. Orally consumed cannabinoids provide long-lasting relief of allodynia in a mouse model of chronic neuropathic Pain. Neuropsychopharmacology 2020, 45, 1105–1114. [Google Scholar] [CrossRef]
- Jourdan, D.; Ardid, D.; Eschalier, A. Analysis of ultrasonic vocalisation does not allow chronic pain to be evaluated in rats. Pain 2002, 95, 165–173. [Google Scholar] [CrossRef]
- Knutson, B.; Burgdorf, J.; Panksepp, J. Ultrasonic vocalizations as indices of affective states in rats. Psychol. Bull. 2002, 128, 961–977. [Google Scholar] [CrossRef] [PubMed]
- Helmer, L.M.L.; Weijenberg, R.A.F.; de Vries Achterberg, R.W.P.; Lautenbacher, S.; Sampson, E.L.; Lobbezoo, F. Crying out in pain—A systematic review into the validity of vocalization as an indicator for Pain. Eur. J. Pain 2020, 24, 1703–1715. [Google Scholar] [CrossRef] [PubMed]
- Neugebauer, V.; Li, W.; Bird, G.C.; Bhave, G.; Gereau, R.W., IV. Synaptic Plasticity in the Amygdala in a Model of Arthritic Pain: Differential Roles of Metabotropic Glutamate Receptors 1 and 5. J. Neurosci. 2003, 23, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Grégoire, S.; Neugebauer, V. 5-HT2CR blockade in the amygdala conveys analgesic efficacy to SSRIs in a rat model of arthritis Pain. Mol. Pain 2013, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Kiritoshi, T.; Neugebauer, V. Group II mGluRs modulate baseline and arthritis pain-related synaptic transmission in the rat medial prefrontal cortex. Neuropharmacology 2015, 95, 388–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzitelli, M.; Neugebauer, V. Amygdala group II mGluRs mediate the inhibitory effects of systemic group II mGluR activation on behavior and spinal neurons in a rat model of arthritis Pain. Neuropharmacology 2019, 158, 107706. [Google Scholar] [CrossRef] [PubMed]
- Mazzitelli, M.; Marshall, K.; Pham, A.; Ji, G.; Neugebauer, V. Optogenetic Manipulations of Amygdala Neurons Modulate Spinal Nociceptive Processing and Behavior Under Normal Conditions and in an Arthritis Pain Model. Front. Pharmacol. 2021, 12, 25. [Google Scholar] [CrossRef]
- Bennett, G.J.; Chung, J.M.; Honore, M.; Seltzer, Z. Models of Neuropathic Pain in the Rat. Curr. Protoc. Neurosci. 2003, 22, 9.14.1–9.14.16. [Google Scholar] [CrossRef]
- Dineley, K.T.; Kayed, R.; Neugebauer, V.; Fu, Y.; Zhang, W.; Reese, L.C.; Taglialatela, T. Amyloid-beta oligomers impair fear conditioned memory in a calcineurin-dependent fashion in mice. J. Neurosci. Res. 2010, 88, 2923–2932. [Google Scholar]
- Martin, Z.S.; Neugebauer, V.; Dineley, K.T.; Kayed, R.; Zhang, W.; Reese, L.C.; Taglialatela, T. Alpha synuclein oligomers oppose long-term potentiation and impair memory through a calcineurin-dependent mechanism: Relevance to human synucleopathic diseases. J. Neurochem. 2012, 120, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Yakhnitsa, V.; Kiritoshi, T.; Presto, P.; Neugebauer, V. Fear extinction learning ability predicts neuropathic pain behaviors and amygdala activity in male rats. Mol. Pain 2018, 14, 1744806918804441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.S.; Li, W.; Neugebauer, V. Critical Role of Calcitonin Gene-Related Peptide 1 Receptors in the Amygdala in Synaptic Plasticity and Pain Behavior. J. Neurosci. 2005, 25, 10717–10728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adwanikar, H.; Ji, G.; Li, W.; Doods, H.; Willis, W.D.; Neugebauer, V. Spinal CGRP1 receptors contribute to supraspinally organized pain behavior and pain-related sensitization of amygdala neurons. Pain 2007, 132, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Palazzo, E.; Fu, Y.; Ji, G.; Maione, S.; Neugebauer, V. Group III mGluR7 and mGluR8 in the amygdala differentially modulate nocifensive and affective pain behaviors. Neuropharmacology 2008, 55, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Hein, M.; Ji, G.; Tidwell, D.; D’Souza, P.; Kiritoshi, T.; Yakhnitsa, V.; Navratilova, E.; Porreca, F.; Neugebauer, V. Kappa opioid receptor activation in the amygdala disinhibits CRF neurons to generate pain-like behaviors. Neuropharmacology 2021, 185, 108456. [Google Scholar] [CrossRef]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to Measure Locomotor and Anxiety-like Behavior in Mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar] [CrossRef] [Green Version]
- Araya, E.I.; Baggio, D.F.; de Koren, L.O.; Andreatini, R.; Schwarting, R.K.W.; Zamponi, G.W.; Chichorro, J.G. Acute orofacial pain leads to prolonged changes in behavioral and affective pain components. Pain 2020, 161, 2830–2840. [Google Scholar] [CrossRef]
- Bush, D.E.A.; Sotres-Bayon, F.; LeDoux, J.E. Individual differences in fear: Isolating fear reactivity and fear recovery phenotypes. J. Trauma. Stress 2007, 20, 413–422. [Google Scholar] [CrossRef]
- Reznikov, R.; Diwan, M.; Nobrega, J.N.; Hamani, C. Towards a better preclinical model of PTSD: Characterizing animals with weak extinction, maladaptive stress responses and low plasma corticosterone. J. Psychiatr. Res. 2015, 61, 158–165. [Google Scholar] [CrossRef]
- Ueno, M.; Yamada, K.; Ichitani, Y. The relationship between fear extinction and resilience to drug-dependence in rats. Neurosci. Res. 2017, 121, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Duits, P.; Richter, J.; Baas, J.M.P.; Engelhard, I.M.; Limberg-Thiesen, A.; Heitland, I.; Hamm, A.O.; Cath, D.C. Enhancing effects of contingency instructions on fear acquisition and extinction in anxiety disorders. J. Abnorm. Psychol. 2017, 126, 378–391. [Google Scholar] [CrossRef] [PubMed]
- Lissek, S.; Powers, A.S.; McClure, E.B.; Phelps, E.A.; Woldehawariat, G.; Grillon, C.; Pine, D.S. Classical fear conditioning in the anxiety disorders: A meta-analysis. Behav. Res. Ther. 2005, 43, 1391–1424. [Google Scholar] [CrossRef] [PubMed]
- Stegmann, Y.; Schiele, M.A.; Schümann, D.; Lonsdorf, T.B.; Zwanzger, P.; Romanos, M.; Reif, A.; Domschke, K.; Deckert, J.; Gamer, M.; et al. Individual differences in human fear generalization—Pattern identification and implications for anxiety disorders. Transl. Psychiatry 2019, 9, 307. [Google Scholar] [CrossRef] [Green Version]
- Milad, M.R.; Rauch, S.L.; Pitman, R.K.; Quirk, G.J. Fear extinction in rats: Implications for human brain imaging and anxiety disorders. Biol. Psychol. 2006, 73, 61–71. [Google Scholar] [CrossRef]
- Milad, M.R.; Orr, S.P.; Lasko, N.B.; Chang, Y.; Rauch, S.L.; Pitman, R.K. Presence and Acquired Origin of Reduced Recall for Fear Extinction in PTSD: Results of a Twin Study. J. Psychiatr. Res. 2008, 42, 515–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milad, M.R.; Furtak, S.C.; Greenberg, J.L.; Keshaviah, A.; Im, J.J.; Falkenstein, M.J.; Jenike, M.; Rauch, S.L.; Wilhelm, S. Deficits in conditioned fear extinction in obsessive-compulsive disorder and neurobiological changes in the fear circuit. JAMA Psychiatry 2013, 70, 608–618. [Google Scholar] [CrossRef]
- Duits, P.; Cath, D.C.; Lissek, S.; Hox, J.J.; Hamm, A.O.; Engelhard, I.M.; Van Den Hout, M.A.; Baas, J.M. Updated meta-analysis of classical fear conditioning in the anxiety disorders. Depress. Anxiety 2015, 32, 239–253. [Google Scholar] [CrossRef]
- Galea, S.; Nandi, A.; Vlahov, D. The epidemiology of post-traumatic stress disorder after disasters. Epidemiol. Rev. 2005, 27, 78–91. [Google Scholar] [CrossRef] [Green Version]
- Crofford, L.J. Chronic Pain: Where the Body Meets the Brain. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 167. [Google Scholar]
- Altemus, M.; Sarvaiya, N.; Epperson, C.N. Sex differences in anxiety and depression clinical perspectives. Front. Neuroendocrinol. 2014, 35, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Donner, N.C.; Lowry, C.A. Sex differences in anxiety and emotional behavior. Pflugers Arch. Eur. J. Psihol. 2013, 465, 601–626. [Google Scholar] [CrossRef]
- Gruene, T.M.; Roberts, E.; Thomas, V.; Ronzio, A.; Shansky, R.M. Sex-specific neuroanatomical correlates of fear expression inprefrontal-amygdala circuits. Biol. Psychiatry 2015, 78, 186–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baran, S.E.; Armstrong, C.E.; Niren, D.C.; Hanna, J.J.; Conrad, C.D. Chronic Stress and Sex Differences on the Recall of Fear Conditioning and Extinction. Neurobiol. Learn. Mem. 2009, 91, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Baran, S.E.; Armstrong, C.E.; Niren, D.C.; Conrad, C.D. Prefrontal cortex lesions and sex differences in fear extinction and perseveration. Learn. Mem. 2010, 17, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Maren, S.; De Oca, B.; Fanselow, M.S. Sex differences in hippocampal long-term potentiation (LTP) and Pavlovian fear conditioning in rats: Positive correlation between LTP and contextual learning. Brain Res. 1994, 661, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Barker, J.M.; Galea, L.A.M. Males show stronger contextual fear conditioning than females after context pre-exposure. Physiol. Behav. 2010, 99, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daviu, N.; Andero, R.; Armario, A.; Nadal, R. Sex differences in the behavioural and hypothalamic–pituitary–adrenal response to contextual fear conditioning in rats. Horm. Behav. 2014, 66, 713–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willadsen, M.; Uengoer, M.; Sługocka, A.; Schwarting, R.K.W.; Homberg, J.R.; Wöhr, M. Fear Extinction and Predictive Trait-Like Inter-Individual Differences in Rats Lacking the Serotonin Transporter. Int. J. Mol. Sci. 2021, 22, 7088. [Google Scholar] [CrossRef]
- Glover, E.M.; Jovanovic, T.; Norrholm, S.D. Estrogen and Extinction of Fear Memories: Implications for Posttraumatic Stress Disorder Treatment. Biol. Psychiatry 2015, 78, 178–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebron-Milad, K.; Milad, M.R. Sex differences, gonadal hormones and the fear extinction network: Implications for anxiety disorders. Biol. Mood Anxiety Disord. 2012, 2, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, M.L.; Stämpfli, P.; Humphreys, B.K.; Vrana, A.; Seifritz, E.; Schweinhardt, P. The impact of pain-related fear on neural pathways of pain modulation in chronic low back Pain. Pain Rep. 2017, 2, e601. [Google Scholar] [CrossRef] [PubMed]
- Nees, F.; Becker, S. Psychological Processes in Chronic Pain: Influences of Reward and Fear Learning as Key Mechanisms—Behavioral Evidence, Neural Circuits, and Maladaptive Changes. Neuroscience 2018, 387, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Phelps, E.A.; LeDoux, J.E. Contributions of the amygdala to emotion processing: From animal models to human behavior. Neuron 2005, 48, 175–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsini, C.A.; Maren, S. Neural and Cellular Mechanisms of Fear and Extinction Memory Formation. Neurosci. Biobehav. Rev. 2012, 36, 1773–1802. [Google Scholar] [CrossRef] [Green Version]
- Herry, C.; Johansen, J.P. Encoding of fear learning and memory in distributed neuronal circuits. Nat. Neurosci. 2014, 17, 1644–1654. [Google Scholar] [CrossRef]
- Hartley, C.A.; Phelps, E.A. Changing Fear: The Neurocircuitry of Emotion Regulation. Neuropsychopharmacology 2010, 35, 136–146. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.M.; Neugebauer, V. Cortico-limbic pain mechanisms. Neurosci. Lett. 2019, 702, 15–23. [Google Scholar] [CrossRef]
- Vachon-Presseau, E.; Tétreault, P.; Petre, B.; Huang, L.; Berger, S.E.; Torbey, S.; Baria, A.T.; Mansour, A.R.; Hashmi, J.A.; Griffith, J.W.; et al. Corticolimbic anatomical characteristics predetermine risk for chronic Pain. Brain 2016, 139, 1958. [Google Scholar] [CrossRef] [Green Version]
- Meier, M.L.; Stämpfli, P.; Vrana, A.; Humphreys, B.K.; Seifritz, E.; Hotz-Boendermaker, S. Neural Correlates of Fear of Movement in Patients with Chronic Low Back Pain vs. Pain-Free Individuals. Front. Hum. Neurosci. 2016, 10, 386. [Google Scholar] [CrossRef] [Green Version]
- Butler, R.K.; Ehling, S.; Barbar, M.; Thomas, J.; Hughes, M.A.; Smith, C.E.; Pogorelov, V.M.; Aryal, D.K.; Wetsel, W.C.; Lascelles, B.D.X. Distinct neuronal populations in the basolateral and central amygdala are activated with acute pain, conditioned fear, and fear-conditioned analgesia. Neurosci. Lett. 2017, 661, 11–17. [Google Scholar] [CrossRef]
- Rea, K.; Roche, M.; Finn, D.P. Modulation of Conditioned Fear, Fear-Conditioned Analgesia, and Brain Regional C-Fos Expression Following Administration of Muscimol into the Rat Basolateral Amygdala. J. Pain 2011, 12, 712–721. [Google Scholar] [CrossRef] [Green Version]
- Rea, K.; Olango, W.M.; Harhen, B.; Kerr, D.M.; Galligan, R.; Fitzgerald, S.; Moore, M.; Roche, M.; Finn, D.P. Evidence for a role of GABAergic and glutamatergic signalling in the basolateral amygdala in endocannabinoid-mediated fear-conditioned analgesia in rats. Pain 2013, 154, 576–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neugebauer, V.; Li, W.; Bird, G.C.; Han, J.S. The amygdala and persistent Pain. Neuroscientist 2004, 10, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.M.; Neugebauer, V. Amygdala Plasticity and Pain. Pain Res. Manag. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.-H.; Zhao, W.-J.; Yin, J.-B.; Chen, Y.-B.; Li, J.-N.; Feng, B.; Lu, Y.-C.; Wang, J.; Dong, Y.-L.; Li, Y.-Q. A Neural Circuit from Thalamic Paraventricular Nucleus to Central Amygdala for the Facilitation of Neuropathic Pain. J. Neurosci. 2020, 40, 7837–7854. [Google Scholar] [CrossRef]
- Missig, G.A.; Roman, C.W.; Vizzard, M.A.; Braas, K.M.; Hammack, S.E.; May, V. Parabrachial nucleus (PBn) pituitary adenylate cyclase activating polypeptide (PACAP) signaling in the amygdala: Implication for the sensory and behavioral effects of Pain. Neuropharmacology 2014, 86, 38–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahrens, S.; Wu, M.V.; Furlan, A.; Hwang, G.-R.; Paik, R.; Li, H.; Penzo, M.A.; Tollkuhn, J.; Li, B. A Central Extended Amygdala Circuit That Modulates Anxiety. J. Neurosci. 2018, 38, 5567–5583. [Google Scholar] [CrossRef] [Green Version]
- Paretkar, T.; Dimitrov, E. The central amygdala corticotropin-releasing hormone (CRH) neurons modulation of anxiety-like behavior and hippocampus-dependent memory in mice. Neuroscience 2018, 390, 187–197. [Google Scholar] [CrossRef]
- Penzo, M.A.; Robert, V.; Tucciarone, J.; De Bundel, D.; Wang, M.; Van Aelst, L.; Darvas, M.; Parada, L.F.; Palmiter, R.D.; He, M.; et al. The paraventricular thalamus controls a central amygdala fear circuit. Nature 2015, 519, 455–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neugebauer, V.; Li, W. Differential sensitization of amygdala neurons to afferent inputs in a model of arthritic Pain. J. Neurophysiol. 2003, 89, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Kiritoshi, T.; Neugebauer, V. Pathway-specific alterations of cortico-amygdala transmission in an arthritis pain model. ACS Chem. Neurosci. 2018, 9, 2252–2261. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Fang, D.; Kong, L.-Y.; Jin, Z.-R.; Cai, J.; Kang, X.-J.; Wan, Y.; Xing, G.-G. Sensitization of neurons in the central nucleus of the amygdala via the decreased GABAergic inhibition contributes to the development of neuropathic pain-related anxiety-like behaviors in rats. Mol. Brain 2014, 7, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engman, J.; Linnman, C.; Van Dijk, K.R.A.; Milad, M.R. Amygdala subnuclei resting-state functional connectivity sex and estrogen differences. Psychoneuroendocrinology 2016, 63, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Labrenz, F.; Knuf-Rtveliashvili, S.; Elsenbruch, S. Sex Differences Linking Pain-Related Fear and Interoceptive Hypervigilance: Attentional Biases to Conditioned Threat and Safety Signals in a Visceral Pain Model. Front. Psychiatry 2020, 11, 197. [Google Scholar] [CrossRef]
- Wöhr, M.; Schwarting, R.K.W. Affective communication in rodents: Ultrasonic vocalizations as a tool for research on emotion and motivation. Cell Tissue Res. 2013, 354, 81–97. [Google Scholar] [CrossRef]
- Knight, P.; Chellian, R.; Wilson, R.; Behnood-Rod, A.; Panunzio, S.; Bruijnzeel, A.W. Sex differences in the elevated plus-maze test and large open field test in adult Wistar rats. Pharmacol. Biochem. Behav. 2021, 204, 173168. [Google Scholar] [CrossRef]
- Won, S.; Park, K.; Lim, H.; Lee, S.J. Sexual dimorphism in cognitive disorders in a murine model of neuropathic Pain. Behav. Brain Funct. 2020, 16, 1. [Google Scholar] [CrossRef]
- Graham, L.K.; Yoon, T.; Lee, H.J.; Kim, J.J. Strain and sex differences in fear conditioning: 22 kHz ultrasonic vocalizations and freezing in rats. Psychol. Neurosci. 2009, 2, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Williams, W.O.; Riskin, D.K.; Mott, K.M. Ultrasonic Sound as an Indicator of Acute Pain in Laboratory Mice. J. Am. Assoc. Lab. Anim. Sci. 2008, 47, 8. [Google Scholar] [PubMed]
- Jourdan, D.; Ardid, D.; Chapuy, E.; Eschalier, A.; Le Bars, D. Audible and ultrasonic vocalization elicited by single electrical nociceptive stimuli to the tail in the rat. Pain 1995, 63, 237–249. [Google Scholar] [CrossRef]
- Tsuzuki, H.; Maekawa, M.; Konno, R.; Hori, Y. Functional roles of endogenous D-serine in pain-induced ultrasonic vocalization. Neuroreport 2012, 23, 937–941. [Google Scholar] [CrossRef]
- Oliveira, A.R.; Barros, H.M.T. Ultrasonic rat vocalizations during the formalin test: A measure of the affective dimension of pain? Anesth. Analg. 2006, 102, 832–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brudzynski, S.M.; Ociepa, D. Ultrasonic vocalization of laboratory rats in response to handling and touch. Physiol. Behav. 1992, 52, 655–660. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Presto, P.; Ji, G.; Junell, R.; Griffin, Z.; Neugebauer, V. Fear Extinction-Based Inter-Individual and Sex Differences in Pain-Related Vocalizations and Anxiety-like Behaviors but Not Nocifensive Reflexes. Brain Sci. 2021, 11, 1339. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11101339
Presto P, Ji G, Junell R, Griffin Z, Neugebauer V. Fear Extinction-Based Inter-Individual and Sex Differences in Pain-Related Vocalizations and Anxiety-like Behaviors but Not Nocifensive Reflexes. Brain Sciences. 2021; 11(10):1339. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11101339
Chicago/Turabian StylePresto, Peyton, Guangchen Ji, Riley Junell, Zach Griffin, and Volker Neugebauer. 2021. "Fear Extinction-Based Inter-Individual and Sex Differences in Pain-Related Vocalizations and Anxiety-like Behaviors but Not Nocifensive Reflexes" Brain Sciences 11, no. 10: 1339. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11101339