
Abnormal Functional Network Topology and Its Dynamics during Sustained Attention Processing Significantly Implicate Post-TBI Attention Deficits in Children


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Neuroimaging Data Acquisition Protocol
2.3. Visual Sustained Attention Task for fMRI
2.4. Individual-Level Neuroimaging Data Pre-Processing
2.5. Network Node Selection and BOLD Signal Extraction
2.6. Static Functional Network Construction
2.7. Analysis of Functional Network Dynamics
2.8. Group-Level Analyses
3. Results
3.1. Demographic and Clinical/Behavioral Measures
3.2. Topological Measures in Overall Functional Network
3.3. Dynamics of the Topological Measures
3.4. Brain Behavior Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dewan, M.C.; Mummareddy, N.; Wellons, J.C., 3rd; Bonfield, C.M. Epidemiology of Global Pediatric Traumatic Brain Injury: Qualitative Review. World Neurosurg. 2016, 91, 497–509.e491. [Google Scholar] [CrossRef]
- Emery, C.A.; Barlow, K.M.; Brooks, B.L.; Max, J.E.; Villavicencio-Requis, A.; Gnanakumar, V.; Robertson, H.L.; Schneider, K.; Yeates, K.O. A Systematic Review of Psychiatric, Psychological, and Behavioural Outcomes following Mild Traumatic Brain Injury in Children and Adolescents. Can. J. Psychiatry 2016, 61, 259–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, S.R.; Alexander, J.; Moore, D.; Sasser, H.C.; Laurent, S.; King, J.; Bartel, S.; Callahan, B.J.N. Caregiver reports of common symptoms in children following a traumatic brain injury. NeuroRehabilitation 2004, 19, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Konigs, M.; Heij, H.A.; van der Sluijs, J.A.; Vermeulen, R.J.; Goslings, J.C.; Luitse, J.S.; Poll-The, B.T.; Beelen, A.; van der Wees, M.; Kemps, R.J.; et al. Pediatric Traumatic Brain Injury and Attention Deficit. Pediatrics 2015, 136, 534–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polinder, S.; Haagsma, J.A.; van Klaveren, D.; Steyerberg, E.W.; van Beeck, E.F. Health-related quality of life after TBI: A systematic review of study design, instruments, measurement properties, and outcome. Popul. Health Metr. 2015, 13, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Max, J.E.; Schachar, R.J.; Levin, H.S.; Ewing-Cobbs, L.; Chapman, S.B.; Dennis, M.; Saunders, A.; Landis, J. Predictors of secondary attention-deficit/hyperactivity disorder in children and adolescents 6 to 24 months after traumatic brain injury. J. Am. Acad. Child Adolesc. Psychiatry 2005, 44, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Narad, M.E.; Kennelly, M.; Zhang, N.; Wade, S.L.; Yeates, K.O.; Taylor, H.G.; Epstein, J.N.; Kurowski, B.G. Secondary Attention-Deficit/Hyperactivity Disorder in Children and Adolescents 5 to 10 Years After Traumatic Brain Injury. JAMA Pediatr. 2018, 172, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Le Fur, C.; Camara-Costa, H.; Francillette, L.; Opatowski, M.; Toure, H.; Brugel, D.; Laurent-Vannier, A.; Meyer, P.; Watier, L.; Dellatolas, G.; et al. Executive functions and attention 7years after severe childhood traumatic brain injury: Results of the Traumatisme Grave de l’Enfant (TGE) cohort. Ann. Phys. Rehabil. Med. 2019, 63, 270–279. [Google Scholar] [CrossRef]
- Narad, M.E.; Riemersma, J.; Wade, S.L.; Smith-Paine, J.; Morrison, P.; Taylor, H.G.; Yeates, K.O.; Kurowski, B.G. Impact of Secondary ADHD on Long-Term Outcomes After Early Childhood Traumatic Brain Injury. J. Head Trauma Rehabil. 2019, 34, 36–44. [Google Scholar] [CrossRef]
- Mayer, A.R.; Bellgowan, P.S.; Hanlon, F.M. Functional magnetic resonance imaging of mild traumatic brain injury. Neurosci. Biobehav. Rev. 2015, 49, 8–18. [Google Scholar] [CrossRef]
- Kramer, M.E.; Chiu, C.Y.; Shear, P.K.; Wade, S.L. Neural correlates of verbal associative memory and mnemonic strategy use following childhood traumatic brain injury. J. Pediatr. Rehabil. Med. 2009, 2, 255–271. [Google Scholar] [CrossRef] [PubMed]
- Westfall, D.R.; West, J.D.; Bailey, J.N.; Arnold, T.W.; Kersey, P.A.; Saykin, A.J.; McDonald, B.C. Increased brain activation during working memory processing after pediatric mild traumatic brain injury (mTBI). J. Pediatr. Rehabil. Med. 2015, 8, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Sinopoli, K.J.; Chen, J.K.; Wells, G.; Fait, P.; Ptito, A.; Taha, T.; Keightley, M. Imaging “brain strain” in youth athletes with mild traumatic brain injury during dual-task performance. J. Neurotrauma 2014, 31, 1843–1859. [Google Scholar] [CrossRef]
- Newsome, M.R.; Steinberg, J.L.; Scheibel, R.S.; Troyanskaya, M.; Chu, Z.; Hanten, G.; Lu, H.; Lane, S.; Lin, X.; Hunter, J.V.; et al. Effects of traumatic brain injury on working memory-related brain activation in adolescents. Neuropsychology 2008, 22, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.L.; Virani, S.; Khetani, A.; Carlson, H.; Jadavji, Z.; Mauthner, M.; Low, T.A.; Plourde, V.; MacMaster, F.P.; Bray, S.; et al. Functional magnetic resonance imaging study of working memory several years after pediatric concussion. Brain Inj. 2020, 34, 895–904. [Google Scholar] [CrossRef]
- Caeyenberghs, K.; Wenderoth, N.; Smits-Engelsman, B.C.; Sunaert, S.; Swinnen, S.P. Neural correlates of motor dysfunction in children with traumatic brain injury: Exploration of compensatory recruitment patterns. Brain 2009, 132, 684–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunanayaka, P.R.; Holland, S.K.; Yuan, W.; Altaye, M.; Jones, B.V.; Michaud, L.J.; Walz, N.C.; Wade, S.L. Neural substrate differences in language networks and associated language-related behavioral impairments in children with TBI: A preliminary fMRI investigation. NeuroRehabilitation 2007, 22, 355–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newsome, M.R.; Scheibel, R.S.; Hanten, G.; Chu, Z.; Steinberg, J.L.; Hunter, J.V.; Lu, H.; Vasquez, A.C.; Li, X.; Lin, X.; et al. Brain activation while thinking about the self from another person’s perspective after traumatic brain injury in adolescents. Neuropsychology 2010, 24, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Kramer, M.E.; Chiu, C.Y.; Walz, N.C.; Holland, S.K.; Yuan, W.; Karunanayaka, P.; Wade, S.L. Long-term neural processing of attention following early childhood traumatic brain injury: fMRI and neurobehavioral outcomes. J. Int. Neuropsychol. Soc. 2008, 14, 424–435. [Google Scholar] [CrossRef] [Green Version]
- Tlustos, S.J.; Peter Chiu, C.Y.; Walz, N.C.; Wade, S.L. Neural substrates of inhibitory and emotional processing in adolescents with traumatic brain injury. J. Pediatr. Rehabil. Med. 2015, 8, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Tlustos, S.J.; Chiu, C.Y.; Walz, N.C.; Holland, S.K.; Bernard, L.; Wade, S.L. Neural correlates of interference control in adolescents with traumatic brain injury: Functional magnetic resonance imaging study of the counting stroop task. J. Int. Neuropsychol. Soc. 2011, 17, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strazzer, S.; Rocca, M.A.; Molteni, E.; De Meo, E.; Recla, M.; Valsasina, P.; Arrigoni, F.; Galbiati, S.; Bardoni, A.; Filippi, M. Altered Recruitment of the Attention Network Is Associated with Disability and Cognitive Impairment in Pediatric Patients with Acquired Brain Injury. Neural Plast. 2015, 2015, 104282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babikian, T.; Merkley, T.; Savage, R.C.; Giza, C.C.; Levin, H. Chronic Aspects of Pediatric Traumatic Brain Injury: Review of the Literature. J. Neurotrauma 2015, 32, 1849–1860. [Google Scholar] [CrossRef] [PubMed]
- Stephens, J.A.; Salorio, C.F.; Gomes, J.P.; Nebel, M.B.; Mostofsky, S.H.; Suskauer, S.J. Response Inhibition Deficits and Altered Motor Network Connectivity in the Chronic Phase of Pediatric Traumatic Brain Injury. J. Neurotrauma 2017, 34, 3117–3123. [Google Scholar] [CrossRef] [PubMed]
- Tuerk, C.; Degeilh, F.; Catroppa, C.; Dooley, J.J.; Kean, M.; Anderson, V.; Beauchamp, M.H. Altered resting-state functional connectivity within the developing social brain after pediatric traumatic brain injury. Hum. Brain Mapp. 2020, 41, 561–576. [Google Scholar] [CrossRef]
- Newsome, M.R.; Scheibel, R.S.; Mayer, A.R.; Chu, Z.D.; Wilde, E.A.; Hanten, G.; Steinberg, J.L.; Lin, X.; Li, X.; Merkley, T.L.; et al. How functional connectivity between emotion regulation structures can be disrupted: Preliminary evidence from adolescents with moderate to severe traumatic brain injury. J. Int. Neuropsychol. Soc. 2013, 19, 911–924. [Google Scholar] [CrossRef]
- Caeyenberghs, K.; Verhelst, H.; Clemente, A.; Wilson, P.H. Mapping the functional connectome in traumatic brain injury: What can graph metrics tell us? Neuroimage 2017, 160, 113–123. [Google Scholar] [CrossRef]
- Nomura, E.M.; Gratton, C.; Visser, R.M.; Kayser, A.; Perez, F.; D’Esposito, M. Double dissociation of two cognitive control networks in patients with focal brain lesions. Proc. Natl. Acad. Sci. USA 2010, 107, 12017–12022. [Google Scholar] [CrossRef] [Green Version]
- Han, K.; Chapman, S.B.; Krawczyk, D.C. Disrupted Intrinsic Connectivity among Default, Dorsal Attention, and Frontoparietal Control Networks in Individuals with Chronic Traumatic Brain Injury. J. Int. Neuropsychol. Soc. 2016, 22, 263–279. [Google Scholar] [CrossRef] [Green Version]
- Pandit, A.S.; Expert, P.; Lambiotte, R.; Bonnelle, V.; Leech, R.; Turkheimer, F.E.; Sharp, D.J. Traumatic brain injury impairs small-world topology. Neurology 2013, 80, 1826–1833. [Google Scholar] [CrossRef] [Green Version]
- Kuceyeski, A.F.; Jamison, K.W.; Owen, J.P.; Raj, A.; Mukherjee, P. Longitudinal increases in structural connectome segregation and functional connectome integration are associated with better recovery after mild TBI. Hum. Brain Mapp. 2019, 40, 4441–4456. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, R.M.; Morton, J.B. Tracking the Brain’s Functional Coupling Dynamics over Development. J. Neurosci. 2015, 35, 6849–6859. [Google Scholar] [CrossRef]
- Braun, U.; Schafer, A.; Walter, H.; Erk, S.; Romanczuk-Seiferth, N.; Haddad, L.; Schweiger, J.I.; Grimm, O.; Heinz, A.; Tost, H.; et al. Dynamic reconfiguration of frontal brain networks during executive cognition in humans. Proc. Natl. Acad. Sci. USA 2015, 112, 11678–11683. [Google Scholar] [CrossRef] [Green Version]
- Kucyi, A.; Hove, M.J.; Esterman, M.; Hutchison, R.M.; Valera, E.M. Dynamic Brain Network Correlates of Spontaneous Fluctuations in Attention. Cereb Cortex 2017, 27, 1831–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, N.; Bernier, R.A.; Calhoun, V.D.; Brenner, E.; Grossner, E.; Rajtmajer, S.M.; Hillary, F.G. Diminished neural network dynamics after moderate and severe traumatic brain injury. PLoS ONE 2018, 13, e0197419. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Sours Rhodes, C.; Jiang, L.; Roys, S.; Zhuo, J.; JaJa, J.; Gullapalli, R.P. Dynamic Functional Network Analysis in Mild Traumatic Brain Injury. Brain Connect. 2019, 9, 475–487. [Google Scholar] [CrossRef]
- Cao, M.; Luo, Y.; Wu, Z.; Mazzola, C.A.; Catania, L.; Alvarez, T.L.; Halperin, J.M.; Biswal, B.; Li, X. Topological Aberrance of Structural Brain Network Provides Quantitative Substrates of Post-Traumatic Brain Injury Attention Deficits in Children. Brain Connect. 2021. Published electronically May 6. [Google Scholar] [CrossRef] [PubMed]
- Teasdale, G.; Jennett, B. Assessment of coma and impaired consciousness. A practical scale. Lancet 1974, 2, 81–84. [Google Scholar] [CrossRef]
- Conners, C.K. Conners 3; Western Psychological Services: Los Angeles, CA, USA, 2008. [Google Scholar]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Wechsler, D. Wechsler Abbreviated Scale of Intelligence–Second Edition (WASI-II); NCS Pearson: Bloomington, MN, USA, 2011. [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®); American Psychiatric Pub: Washington, DC, USA, 2013. [Google Scholar]
- Kaufman, J.; Birmaher, B.; Brent, D.A.; Ryan, N.D.; Rao, U. Schedule for Affective Disorders and Schizophrenia for School-Age Children-Present and Lifetime Version (K-SADS-PL): Initial Reliability and Validity Data. J. Am. Acad. Child. Adolesc. Psychiatry 2000, 36, 980–988. [Google Scholar] [CrossRef]
- Blakemore, S.J.; Choudhury, S. Development of the adolescent brain: Implications for executive function and social cognition. J. Child. Psychol. Psychiatry 2006, 47, 296–312. [Google Scholar] [CrossRef] [PubMed]
- Carskadon, M.A.; Acebo, C. A self-administered rating scale for pubertal development. J. Adolesc. Health 1993, 14, 190–195. [Google Scholar] [CrossRef]
- Wassenberg, R.; Max, J.E.; Lindgren, S.D.; Schatz, A. Sustained attention in children and adolescents after traumatic brain injury: Relation to severity of injury, adaptive functioning, ADHD and social background. Brain Inj. 2004, 18, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Ornstein, T.J.; Sagar, S.; Schachar, R.J.; Ewing-Cobbs, L.; Chapman, S.B.; Dennis, M.; Saunders, A.E.; Yang, T.T.; Levin, H.S.; Max, J.E. Neuropsychological performance of youth with secondary attention-deficit/hyperactivity disorder 6- and 12-months after traumatic brain injury. J. Int. Neuropsychol. Soc. 2014, 20, 971–981. [Google Scholar] [CrossRef] [Green Version]
- Ginstfeldt, T.; Emanuelson, I. An overview of attention deficits after paediatric traumatic brain injury. Brain Inj. 2010, 24, 1123–1134. [Google Scholar] [CrossRef]
- Li, X.; Sroubek, A.; Kelly, M.S.; Lesser, I.; Sussman, E.; He, Y.; Branch, C.; Foxe, J.J. Atypical pulvinar-cortical pathways during sustained attention performance in children with attention-deficit/hyperactivity disorder. J. Am. Acad. Child. Adolesc. Psychiatry 2012, 51, 1197–1207.e1194. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Foxe, J.J.; Sroubek, A.E.; Branch, C.; Li, X. Topological organization of the “small-world” visual attention network in children with attention deficit/hyperactivity disorder (ADHD). Front. Hum. Neurosci. 2014, 8, 162. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Mazzola, C.A.; Catania, L.; Owoeye, O.; Yaramothu, C.; Alvarez, T.; Gao, Y.; Li, X. Altered cortical activation and connectivity patterns for visual attention processing in young adults post-traumatic brain injury: A functional near infrared spectroscopy study. CNS Neurosci. Ther. 2018, 24, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Woolrich, M.W.; Ripley, B.D.; Brady, M.; Smith, S.M. Temporal autocorrelation in univariate linear modeling of FMRI data. Neuroimage 2001, 14, 1370–1386. [Google Scholar] [CrossRef] [Green Version]
- Satterthwaite, T.D.; Wolf, D.H.; Loughead, J.; Ruparel, K.; Elliott, M.A.; Hakonarson, H.; Gur, R.C.; Gur, R.E. Impact of in-scanner head motion on multiple measures of functional connectivity: Relevance for studies of neurodevelopment in youth. Neuroimage 2012, 60, 623–632. [Google Scholar] [CrossRef] [Green Version]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage 2012, 59, 2142–2154. [Google Scholar] [CrossRef] [Green Version]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Percival, D.B.; Walden, A.T. Wavelet Methods for Time Series Analysis; Cambridge University Press: Cambridge, UK, 2000; Volume 4. [Google Scholar]
- Bassett, D.S.; Wymbs, N.F.; Porter, M.A.; Mucha, P.J.; Carlson, J.M.; Grafton, S.T. Dynamic reconfiguration of human brain networks during learning. Proc. Natl. Acad. Sci. USA 2011, 108, 7641–7646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latora, V.; Marchiori, M. Efficient behavior of small-world networks. Phys. Rev. Lett 2001, 87, 198701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achard, S.; Bullmore, E. Efficiency and cost of economical brain functional networks. PLoS Comput. Biol. 2007, 3, e17. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, M.P.; de Lange, S.C.; Zalesky, A.; Seguin, C.; Yeo, B.T.T.; Schmidt, R. Proportional thresholding in resting-state fMRI functional connectivity networks and consequences for patient-control connectome studies: Issues and recommendations. Neuroimage 2017, 152, 437–449. [Google Scholar] [CrossRef]
- Onnela, J.P.; Saramaki, J.; Kertesz, J.; Kaski, K. Intensity and coherence of motifs in weighted complex networks. Phys. Rev. E 2005, 71, 065103. [Google Scholar] [CrossRef] [Green Version]
- Freeman, L.C. Centrality in social networks conceptual clarification. Soc. Netw. 1978, 1, 215–239. [Google Scholar] [CrossRef] [Green Version]
- Jolly, A.E.; Scott, G.T.; Sharp, D.J.; Hampshire, A.H. Distinct patterns of structural damage underlie working memory and reasoning deficits after traumatic brain injury. Brain 2020, 143, 1158–1176. [Google Scholar] [CrossRef]
- Fagerholm, E.D.; Hellyer, P.J.; Scott, G.; Leech, R.; Sharp, D.J. Disconnection of network hubs and cognitive impairment after traumatic brain injury. Brain 2015, 138, 1696–1709. [Google Scholar] [CrossRef] [Green Version]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Green, G.H.; Diggle, P.J. On the operational characteristics of the Benjamini and Hochberg false discovery rate procedure. Stat. Appl. Genet. Mol. Biol. 2007, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Li, C.S.; Huang, C.; Constable, R.T.; Sinha, R. Imaging response inhibition in a stop-signal task: Neural correlates independent of signal monitoring and post-response processing. J. Neurosci. 2006, 26, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Levin, H.S.; Hanten, G.; Zhang, L.; Swank, P.R.; Hunter, J. Selective impairment of inhibition after TBI in children. J. Clin. Exp. Neuropsychol. 2004, 26, 589–597. [Google Scholar] [CrossRef]
- Levin, H.S.; Wilde, E.A.; Chu, Z.; Yallampalli, R.; Hanten, G.R.; Li, X.; Chia, J.; Vasquez, A.C.; Hunter, J.V. Diffusion tensor imaging in relation to cognitive and functional outcome of traumatic brain injury in children. J. Head Trauma Rehabil. 2008, 23, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Konrad, K.; Gauggel, S.; Manz, A.; Scholl, M. Inhibitory control in children with traumatic brain injury (TBI) and children with attention deficit/hyperactivity disorder (ADHD). Brain Inj. 2000, 14, 859–875. [Google Scholar] [CrossRef]
- Buschman, T.J.; Miller, E.K. Top-down versus bottom-up control of attention in the prefrontal and posterior parietal cortices. Science 2007, 315, 1860–1862. [Google Scholar] [CrossRef] [Green Version]
- Katsuki, F.; Constantinidis, C. Bottom-up and top-down attention: Different processes and overlapping neural systems. Neuroscientist 2014, 20, 509–521. [Google Scholar] [CrossRef]
- Macaluso, E.; Frith, C.D.; Driver, J. Modulation of human visual cortex by crossmodal spatial attention. Science 2000, 289, 1206–1208. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.L.; Kahn, I.; Snyder, A.Z.; Raichle, M.E.; Buckner, R.L. Evidence for a frontoparietal control system revealed by intrinsic functional connectivity. J. Neurophysiol. 2008, 100, 3328–3342. [Google Scholar] [CrossRef] [Green Version]
- Venkatesan, U.M.; Hillary, F.G. Functional connectivity within lateral posterior parietal cortex in moderate to severe traumatic brain injury. Neuropsychology 2019, 33, 893–910. [Google Scholar] [CrossRef]
- Grill-Spector, K.; Weiner, K.S. The functional architecture of the ventral temporal cortex and its role in categorization. Nat. Rev. Neurosci. 2014, 15, 536–548. [Google Scholar] [CrossRef]
- Haxby, J.V.; Gobbini, M.I.; Furey, M.L.; Ishai, A.; Schouten, J.L.; Pietrini, P. Distributed and overlapping representations of faces and objects in ventral temporal cortex. Science 2001, 293, 2425–2430. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wei, Q.; Bai, T.; Zhou, X.; Sun, H.; Becker, B.; Tian, Y.; Wang, K.; Kendrick, K. Electroconvulsive therapy selectively enhanced feedforward connectivity from fusiform face area to amygdala in major depressive disorder. Soc. Cogn. Affect. Neurosci. 2017, 12, 1983–1992. [Google Scholar] [CrossRef]
- Ho, T.C.; Zhang, S.; Sacchet, M.D.; Weng, H.; Connolly, C.G.; Henje Blom, E.; Han, L.K.; Mobayed, N.O.; Yang, T.T. Fusiform Gyrus Dysfunction is Associated with Perceptual Processing Efficiency to Emotional Faces in Adolescent Depression: A Model-Based Approach. Front. Psychol. 2016, 7, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.U.; Shenton, M.E.; Salisbury, D.F.; Kasai, K.; Onitsuka, T.; Dickey, C.C.; Yurgelun-Todd, D.; Kikinis, R.; Jolesz, F.A.; McCarley, R.W. Fusiform gyrus volume reduction in first-episode schizophrenia: A magnetic resonance imaging study. Arch. Gen. Psychiatry 2002, 59, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, S.M.; All, S.D.; Kasi, R.; Berten, S.; Essex, B.; Lathrop, K.L.; Little, D.M. Increased fusiform area activation in schizophrenia during processing of spatial frequency-degraded faces, as revealed by fMRI. Psychol. Med. 2010, 40, 1159–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlman, S.B.; Fournier, J.C.; Bebko, G.; Bertocci, M.A.; Hinze, A.K.; Bonar, L.; Almeida, J.R.; Versace, A.; Schirda, C.; Travis, M.; et al. Emotional face processing in pediatric bipolar disorder: Evidence for functional impairments in the fusiform gyrus. J. Am. Acad. Child. Adolesc. Psychiatry 2013, 52, 1314–1325.e1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strakowski, S.M.; Adler, C.M.; Holland, S.K.; Mills, N.; DelBello, M.P. A preliminary FMRI study of sustained attention in euthymic, unmedicated bipolar disorder. Neuropsychopharmacology 2004, 29, 1734–1740. [Google Scholar] [CrossRef] [PubMed]
- Fujii, D.E.; Ahmed, I. Psychotic disorder caused by traumatic brain injury. Psychiatr. Clin. N. Am. 2014, 37, 113–124. [Google Scholar] [CrossRef]
- Molloy, C.; Conroy, R.M.; Cotter, D.R.; Cannon, M. Is traumatic brain injury a risk factor for schizophrenia? A meta-analysis of case-controlled population-based studies. Schizophr. Bull. 2011, 37, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Rabner, J.; Gottlieb, S.; Lazdowsky, L.; LeBel, A. Psychosis following traumatic brain injury and cannabis use in late adolescence. Am. J. Addict. 2016, 25, 91–93. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
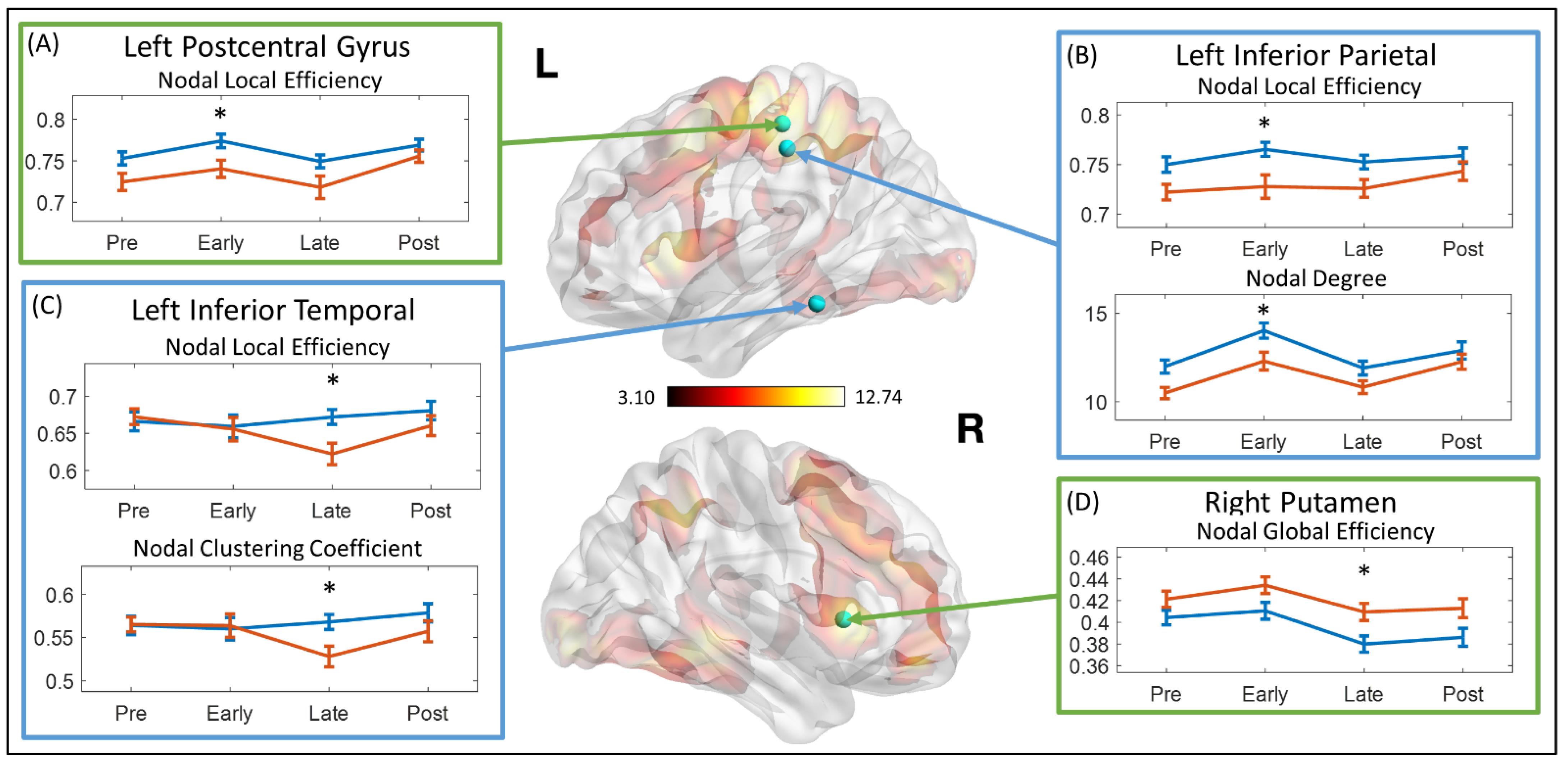{kind=link}
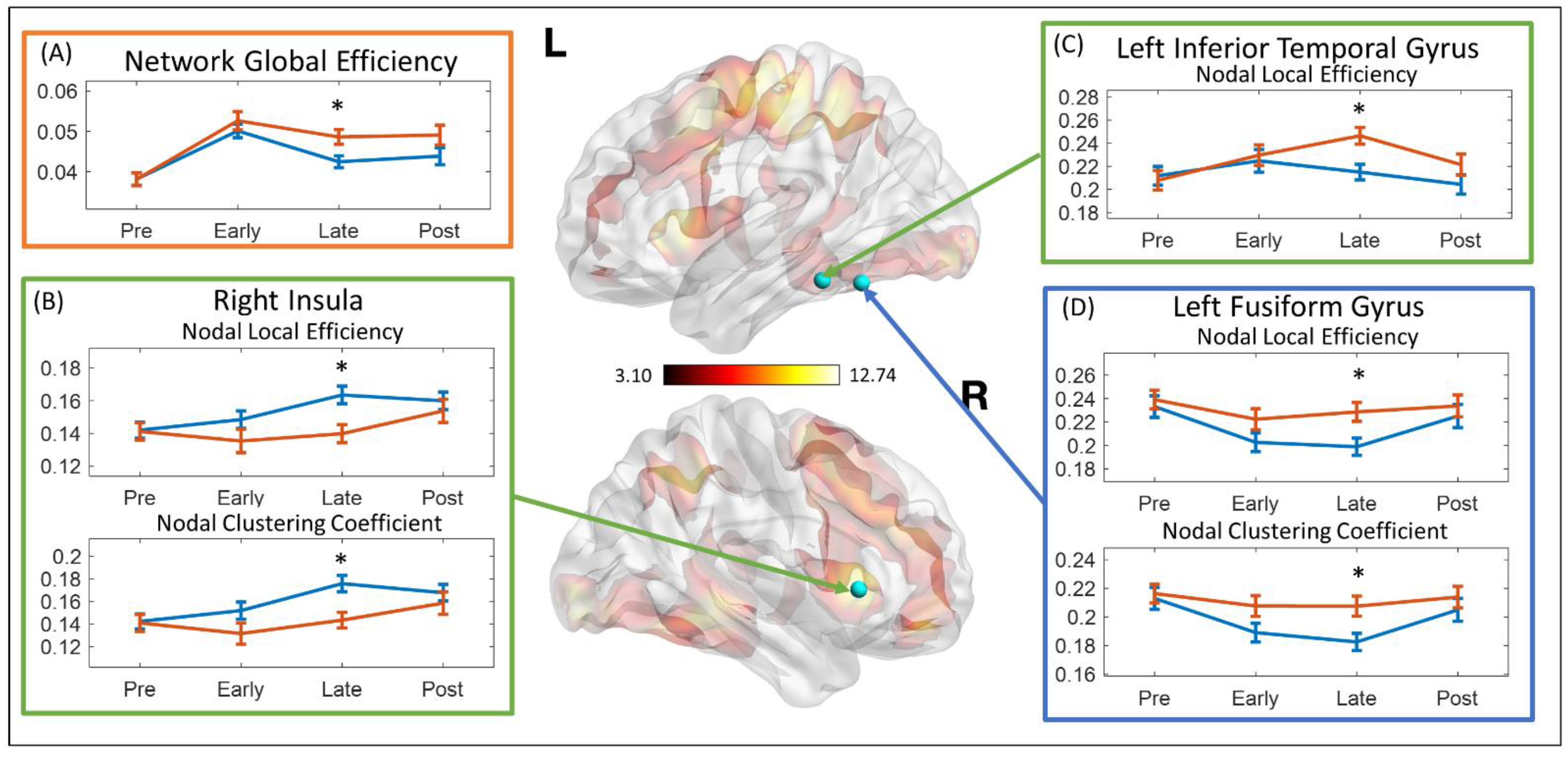{kind=link}
{kind=link}
| Controls Mean (SD) | TBI-A Mean (SD) | t or χ2 Value | p Value | |
|---|---|---|---|---|
| n | 46 | 40 | ||
| Male/Female | 26/20 | 24/16 | 0.106 (χ2) | 0.744 |
| Socioeconomic Status 1 | 16.19 (1.80) | 15.57 (4.19) | 1.450 | 0.151 |
| Full Scale IQ | 115.00 (10.60) | 110.48 (12.68) | 1.802 | 0.075 |
| Age | 13.24 (1.47) | 13.27 (1.72) | −0.075 | 0.940 |
| Ethnicity/Race | 2.411 (χ2) | 0.300 | ||
| Caucasian | 27 | 29 | ||
| Hispanic | 8 | 3 | ||
| Others | 11 | 8 | ||
| Conners 3–PS-based T Score | ||||
| Inattention | 46.67 (6.22) | 71.75 (8.09) | −16.366 | <0.001 |
| Hyperactivity/Impulsivity | 47.91 (5.68) | 63.50 (14.23) | −6.835 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, M.; Halperin, J.M.; Li, X. Abnormal Functional Network Topology and Its Dynamics during Sustained Attention Processing Significantly Implicate Post-TBI Attention Deficits in Children. Brain Sci. 2021, 11, 1348. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11101348
Cao M, Halperin JM, Li X. Abnormal Functional Network Topology and Its Dynamics during Sustained Attention Processing Significantly Implicate Post-TBI Attention Deficits in Children. Brain Sciences. 2021; 11(10):1348. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11101348
Chicago/Turabian StyleCao, Meng, Jeffery M. Halperin, and Xiaobo Li. 2021. "Abnormal Functional Network Topology and Its Dynamics during Sustained Attention Processing Significantly Implicate Post-TBI Attention Deficits in Children" Brain Sciences 11, no. 10: 1348. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11101348