
Impact of Early Rhythmic Training on Language Acquisition and Electrophysiological Functioning Underlying Auditory Processing: Feasibility and Preliminary Findings in Typically Developing Infants

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
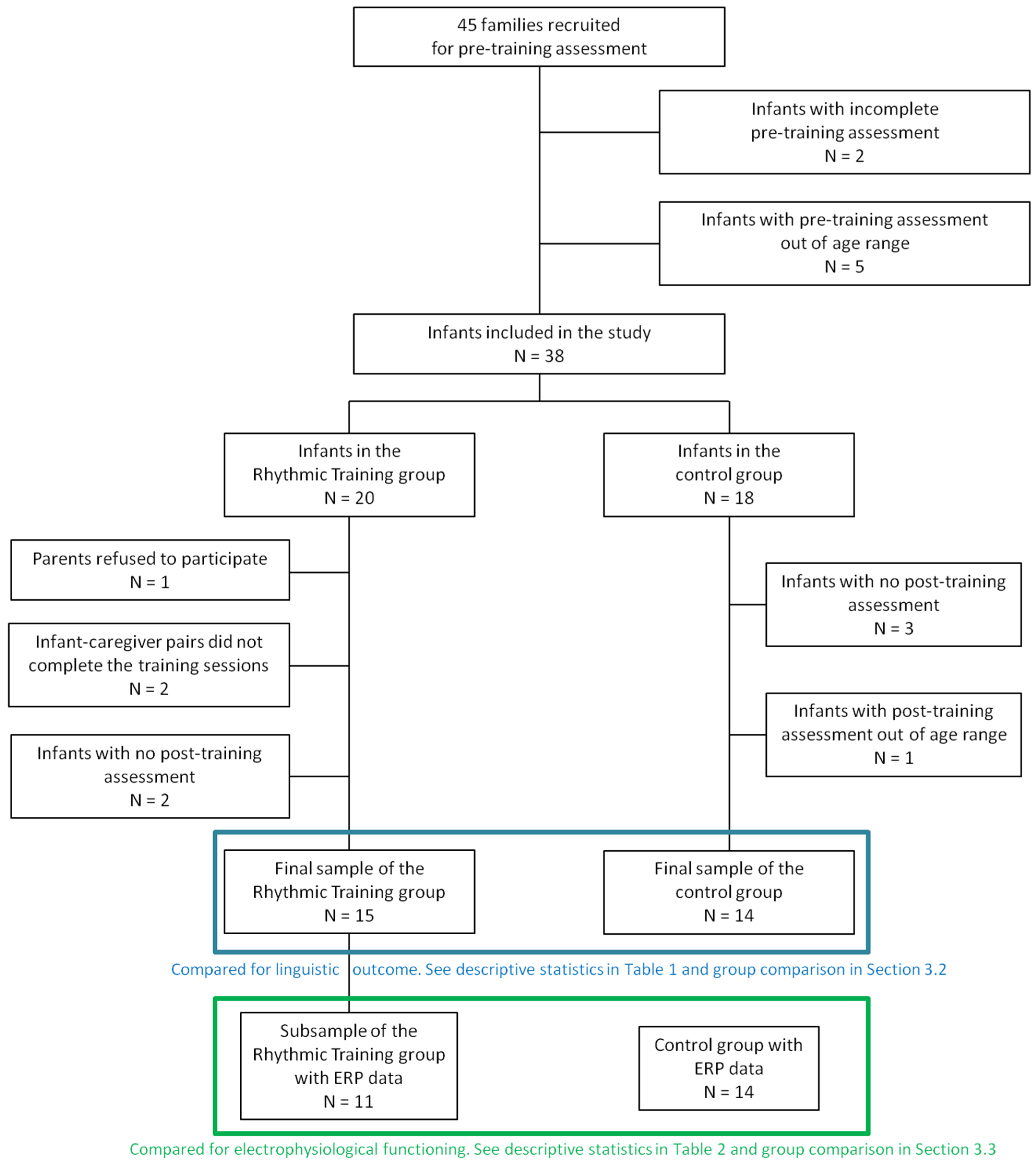
2.1. Participants
2.2. Procedure and Study Design
2.3. Tools
2.3.1. Bayley Scales of Infant and Toddler Development
2.3.2. The Brief Music Experience Questionnaire
2.3.3. EEG paradigm, Data Acquisition and Pre-Processing
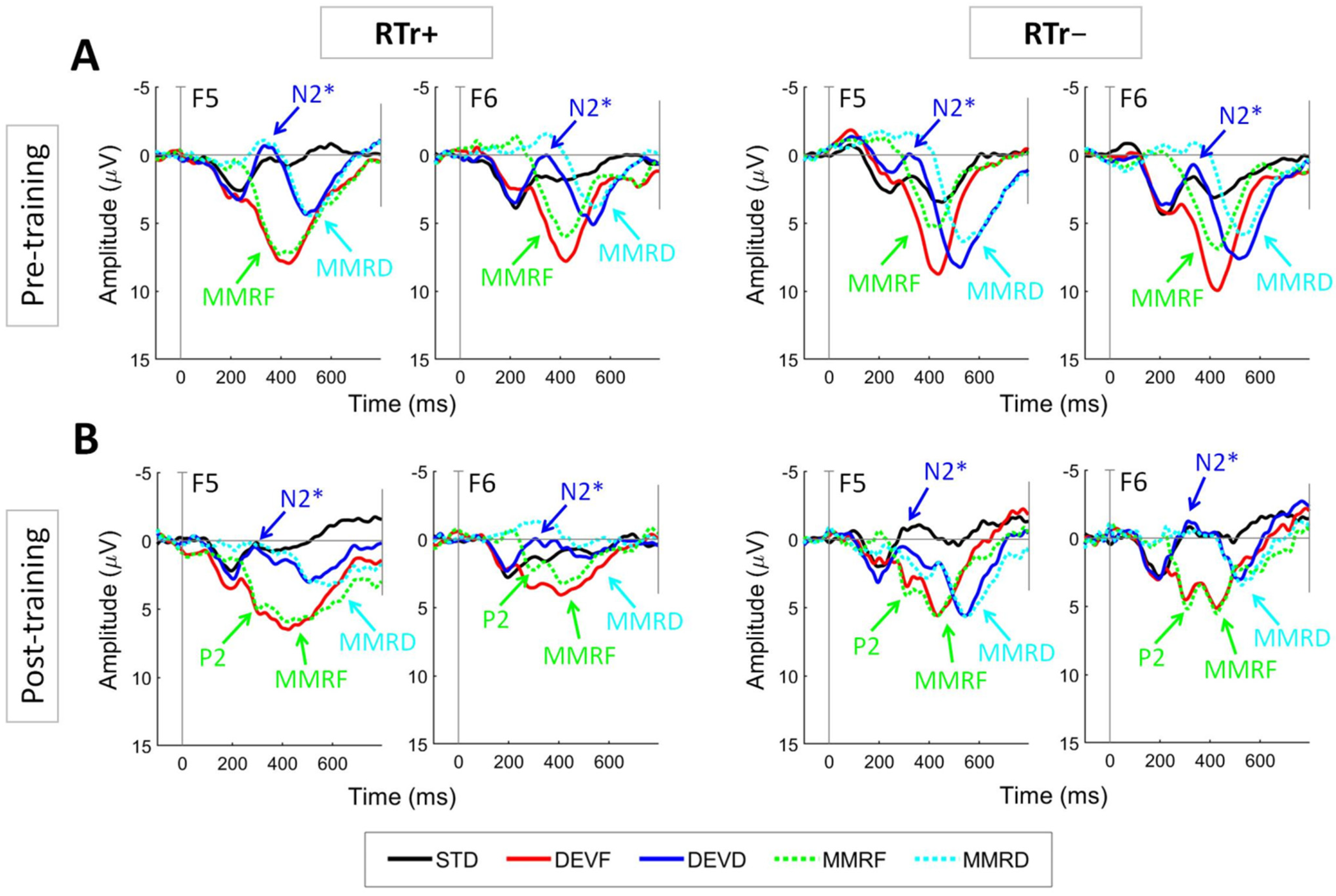
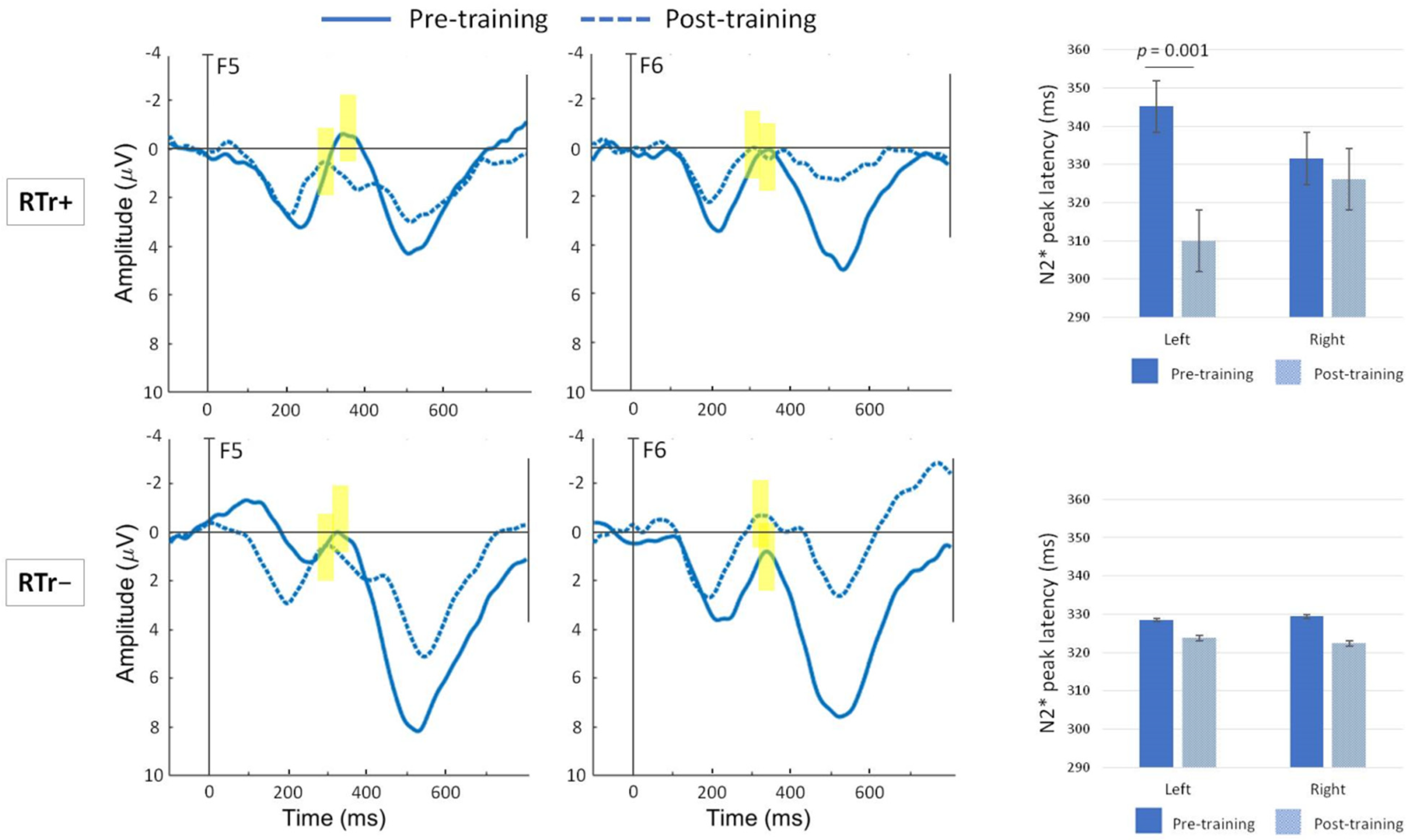
- N2*: 250–400 ms for DEVD (peak amplitude and latency).
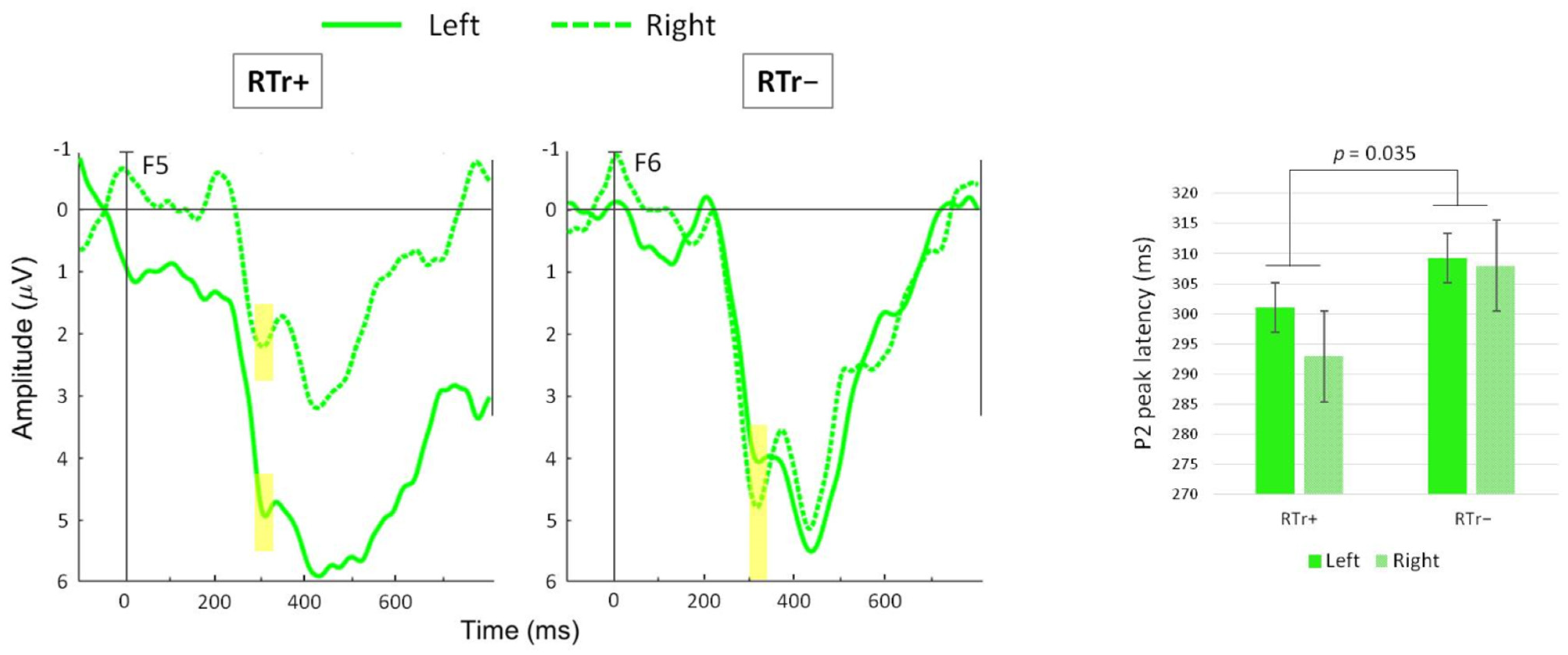
- P2: 250–350 ms for DEVF (different waveform, peak amplitude and latency).
- MMR: 350–550 ms for STD and DEVF and 420–620 ms for DEVD (different waveforms, mean amplitude).
2.4. Early Rhythmic Training
2.5. Statistical Analyses
3. Results
3.1. Behavioral Pre-Training Assessment
3.2. Effect of Training on Language Skills
3.3. Preliminary Findings on the ERP Oddball Paradigm
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Endress, A.D.; Nespor, M.; Mehler, J. Perceptual and memory constraints on language acquisition. Trends Cogn. Sci. 2009, 13, 348–353. [Google Scholar] [CrossRef]
- Saffran, J.R.; Estes, K.G. Mapping sound to meaning: Connections between learning about sounds and learning about words. Adv. Child Dev. Behav. 2006, 34, 1–38. [Google Scholar] [CrossRef]
- Hannon, E.E.; Trainor, L.J. Music acquisition: Effects of enculturation and formal training on development. Trends Cogn. Sci. 2007, 11, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Benasich, A.A.; Tallal, P. Infant discrimination of rapid auditory cues predicts later language impairment. Behav. Brain Res. 2002, 136, 31–49. [Google Scholar] [CrossRef]
- Choudhury, N.; Benasich, A.A. Maturation of auditory evoked potentials from 6 to 48 months: Prediction to 3 and 4 year language and cognitive abilities. Clin. Neurophysiol. 2011, 122, 320–338. [Google Scholar] [CrossRef] [PubMed]
- Cantiani, C.; Riva, V.; Piazza, C.; Bettoni, R.; Molteni, M.; Choudhury, N.; Marino, C.; Benasich, A.A. Auditory discrimination predicts linguistic outcome in Italian infants with and without familial risk for language learning impairment. Dev. Cogn. Neurosci. 2016, 20, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohvansuu, K.; Hämäläinen, J.A.; Ervast, L.; Lyytinen, H.; Leppänen, P.H.T. Longitudinal interactions between brain and cognitive measures on reading development from 6 months to 14 years. Neuropsychologia 2018, 108, 6–12. [Google Scholar] [CrossRef]
- Van Zuijen, T.L.; Plakas, A.; Maassen, B.A.M.; Been, P.; Maurits, N.M.; Krikhaar, E.; van Driel, J.; van der Leij, A. Temporal auditory processing at 17 months of age is associated with preliterate language comprehension and later word reading fluency: An ERP study. Neurosci. Lett. 2012, 528, 31–35. [Google Scholar] [CrossRef]
- Hämäläinen, J.A.; Salminen, H.K.; Leppänen, P.H.T. Basic Auditory Processing Deficits in Dyslexia: Systematic Review of the Behavioral and Event-Related Potential/ Field Evidence. J. Learn. Disabil. 2012, 46, 413–427. [Google Scholar] [CrossRef]
- Cantiani, C.; Lorusso, M.L.; Valnegri, C.; Molteni, M. Perception of non-verbal auditory stimuli in Italian dyslexic children. Dev. Neuropsychol. 2010, 35, 115–123. [Google Scholar] [CrossRef]
- Choudhury, N.; Leppanen, P.H.T.; Leevers, H.J.; Benasich, A.A. Infant information processing and family history of specific language impairment: Converging evidence for RAP deficits from two paradigms. Dev. Sci. 2007, 10, 236. [Google Scholar] [CrossRef] [PubMed]
- Leppänen, P.H.T.; Richardson, U.; Pihko, E.; Eklund, K.M.; Guttorm, T.K.; Aro, M.; Lyytinen, H. Brain Responses to Changes in Speech Sound Durations Differ Between Infants With and Without Familial Risk for Dyslexia. Dev. Neuropsychol. 2010, 22, 407–422. [Google Scholar] [CrossRef] [PubMed]
- Cantiani, C.; Ortiz-Mantilla, S.; Riva, V.; Piazza, C.; Bettoni, R.; Musacchia, G.; Molteni, M.; Marino, C.; Benasich, A.A. Clinical Reduced left-lateralized pattern of event-related EEG oscillations in infants at familial risk for language and learning impairment. Neuroimage Clin. 2019, 22, 101778. [Google Scholar] [CrossRef]
- Riva, V.; Cantiani, C.; Mornati, G.; Gallo, M.; Villa, L.; Mani, E.; Saviozzi, I.; Marino, C.; Molteni, M. Distinct ERP profiles for auditory processing in infants at-risk for autism and language impairment. Sci. Rep. 2018, 8, 715. [Google Scholar] [CrossRef] [Green Version]
- Piazza, C.; Cantiani, C.; Akalin-Acar, Z.; Miyakoshi, M.; Benasich, A.A.; Reni, G.; Bianchi, A.M.; Makeig, S. ICA-derived cortical responses indexing rapid multi-feature auditory processing in six-month-old infants. Neuroimage 2016, 133, 75–87. [Google Scholar] [CrossRef]
- Riva, V.; Cantiani, C.; Benasich, A.A.; Molteni, M.; Piazza, C.; Giorda, R.; Dionne, G.; Marino, C. From CNTNAP2 to Early Expressive Language in Infancy: The Mediation Role of Rapid Auditory Processing. Cereb. Cortex 2018, 28, 2100–2108. [Google Scholar] [CrossRef]
- Benasich, A.A.; Choudhury, N.A.; Realpe-Bonilla, T.; Roesler, C.P. Plasticity in Developing Brain: Active Auditory Exposure Impacts Prelinguistic Acoustic Mapping. J. Neurosci. 2014, 34, 13349–13363. [Google Scholar] [CrossRef] [PubMed]
- Musacchia, G.; Ortiz-Mantilla, S.; Choudhury, N.; Realpe-Bonilla, T.; Roesler, C.; Benasich, A.A. Active auditory experience in infancy promotes brain plasticity in Theta and Gamma oscillations. Dev. Cogn. Neurosci. 2017, 26, 9–19. [Google Scholar] [CrossRef]
- Ortiz-Mantilla, S.; Realpe-Bonilla, T.; Benasich, A.A. Early Interactive Acoustic Experience with Non-speech Generalizes to Speech and Confers a Syllabic Processing Advantage at 9 Months. Cereb. Cortex 2019, 29, 1789–1801. [Google Scholar] [CrossRef]
- Kraus, N.; Skoe, E.; Parbery-Clark, A.; Ashley, R. Experience-induced Malleability in Neural Encoding of Pitch, Timbre, and Timing: Implications for Language and Music. Ann. N. Y. Acad. Sci. 2009, 1169, 557. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.D. Can nonlinguistic musical training change the way the brain processes speech? The expanded OPERA hypothesis. Hear. Res. 2014, 308, 98–108. [Google Scholar] [CrossRef]
- Kraus, N.; Chandrasekaran, B. Music training for the development of auditory skills. Nat. Rev. Neurosci. 2010, 11, 599–605. [Google Scholar] [CrossRef]
- Pantev, C.; Oostenveld, R.; Engelien, A.; Ross, B.; Roberts, L.E.; Hoke, M. Increased auditory cortical representation in musicians. Nature 1998, 392, 811–814. [Google Scholar] [CrossRef]
- Parbery-Clark, A.; Tierney, A.; Strait, D.L.; Kraus, N. Musicians have fine-tuned neural distinction of speech syllables. Neuroscience 2012, 219, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Zuk, J.; Ozernov-Palchik, O.; Kim, H.; Lakshminarayanan, K.; Gabrieli, J.D.E.; Tallal, P.; Gaab, N. Enhanced Syllable Discrimination Thresholds in Musicians. PLoS ONE 2013, 8, e80546. [Google Scholar] [CrossRef] [Green Version]
- Moreno, S.; Marques, C.; Santos, A.; Santos, M.; Castro, S.L.; Besson, M. Musical Training Influences Linguistic Abilities in 8-Year-Old Children: More Evidence for Brain Plasticity. Cereb. Cortex 2009, 19, 712–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, S.; Bialystok, E.; Barac, R.; Schellenberg, E.G.; Cepeda, N.J.; Chau, T. Short-Term Music Training Enhances Verbal Intelligence and Executive Function. Psychol. Sci. 2011, 22, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Bhide, A.; Power, A.; Goswami, U. A Rhythmic Musical Intervention for Poor Readers: A Comparison of Efficacy With a Letter-Based Intervention. Mind Brain Educ. 2013, 7, 113–123. [Google Scholar] [CrossRef]
- Kraus, N.; Slater, J.; Thompson, E.C.; Hornickel, J.; Strait, D.L.; Nicol, T.; White-Schwoch, T. Music Enrichment Programs Improve the Neural Encoding of Speech in At-Risk Children. J. Neurosci. 2014, 34, 11913–11918. [Google Scholar] [CrossRef] [PubMed]
- Flaugnacco, E.; Lopez, L.; Terribili, C.; Montico, M.; Zoia, S.; Schön, D. Music Training Increases Phonological Awareness and Reading Skills in Developmental Dyslexia: A Randomized Control Trial. PLoS ONE 2015, 10, e0138715. [Google Scholar] [CrossRef]
- Musacchia, G.; Strait, D.; Kraus, N. Relationships between behavior, brainstem and cortical encoding of seen and heard speech in musicians and non-musicians. Hear. Res. 2008, 241, 34–42. [Google Scholar] [CrossRef]
- Wong, P.C.M.; Skoe, E.; Russo, N.M.; Dees, T.; Kraus, N. Musical experience shapes human brainstem encoding of linguistic pitch patterns. Nat. Neurosci. 2007, 10, 420–422. [Google Scholar] [CrossRef] [Green Version]
- Doelling, K.B.; Poeppel, D. Cortical entrainment to music and its modulation by expertise. Proc. Natl. Acad. Sci. USA 2015, 112, E6233–E6242. [Google Scholar] [CrossRef] [Green Version]
- Herholz, S.C.; Zatorre, R.J. Musical Training as a Framework for Brain Plasticity: Behavior, Function, and Structure. Neuron 2012, 76, 486–502. [Google Scholar] [CrossRef] [Green Version]
- Gaser, C.; Schlaug, G. Gray Matter Differences between Musicians and Nonmusicians. Ann. N. Y. Acad. Sci. 2003, 999, 514–517. [Google Scholar] [CrossRef]
- Ohnishi, T.; Matsuda, H.; Asada, T.; Aruga, M.; Hirakata, M.; Nishikawa, M.; Katoh, A.; Imabayashi, E. Functional Anatomy of Musical Perception in Musicians. Cereb. Cortex 2001, 11, 754–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlaug, G.; Jäncke, L.; Huang, Y.; Staiger, J.F.; Steinmetz, H. Increased corpus callosum size in musicians. Neuropsychologia 1995, 33, 1047–1055. [Google Scholar] [CrossRef] [Green Version]
- Winkler, I.; Háden, G.P.; Ladinig, O.; Sziller, I.; Honing, H. Newborn infants detect the beat in music. Proc. Natl. Acad. Sci. USA 2009, 106, 2468–2471. [Google Scholar] [CrossRef] [Green Version]
- Gerry, D.W.; Faux, A.L.; Trainor, L.J. Effects of Kindermusik training on infants’ rhythmic enculturation. Dev. Sci. 2010, 13, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Gerry, D.; Unrau, A.; Trainor, L.J. Active music classes in infancy enhance musical, communicative and social development. Dev. Sci. 2012, 15, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.C.; Kuhl, P.K. Musical intervention enhances infants’ neural processing of temporal structure in music and speech. Proc. Natl. Acad. Sci. USA 2016, 113, 5212–5217. [Google Scholar] [CrossRef] [Green Version]
- Virtala, P.; Partanen, E. Can very early music interventions promote at-risk infants’ development? Ann. N. Y. Acad. Sci. 2018, 1423, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Partanen, E.; Kujala, T.; Tervaniemi, M.; Huotilainen, M. Prenatal Music Exposure Induces Long-Term Neural Effects. PLoS ONE 2013, 8, e78946. [Google Scholar] [CrossRef] [Green Version]
- Hollingshead, A.B. Four Factor Index of Social Status; Yale University 1975. Available online: https://sociology.yale.edu/sites/default/files/files/yjs_fall_2011.pdf#page=21 (accessed on 15 November 2021).
- Werner, P.D.; Swope, A.J.; Heide, F.J. The Music Experience Questionnaire: Development and correlates. J. Psychol. 2006, 140, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Bayley, N. Bayley Scales of Infant and Toddler Development; PsychCorp, Pearson: London, UK, 2006. [Google Scholar]
- Piazza, C.; Cantiani, C.; Miyakoshi, M.; Riva, V.; Molteni, M.; Reni, G.; Makeig, S. EEG Effective Source Projections Are More Bilaterally Symmetric in Infants Than in Adults. Front. Hum. Neurosci. 2020, 14, 82. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Calderon, J.; Luck, S.J.; Heekeren, H.R. ERPLAB: An open-source toolbox for the analysis of event-related potentials. Front. Hum. Neurosci. 2014, 8, 213. [Google Scholar] [CrossRef] [Green Version]
- AbyRhythms—Music for Early Development. Available online: http://www.babyrhythms.com/ (accessed on 15 October 2021).
- Weitzman, E.; Girolametto, L.; Drake, L. Hanen Programs® for parents: Parent implemented early language intervention. In Treatment of Language Disorders in Children; McCauley, R.J., Fey, M.E., Gillam, R.B., Eds.; Paul H Brookes Publishing: Baltimore, MD, USA, 2017; Volume 2, pp. 27–56. [Google Scholar]
- Phillips-Silver, J.; Trainor, L.J. Feeling the Beat: Movement Influences Infant Rhythm Perception. Science 2005, 308, 1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirelli, L.K.; Einarson, K.M.; Trainor, L.J. Interpersonal synchrony increases prosocial behavior in infants. Dev. Sci. 2014, 17, 1003–1011. [Google Scholar] [CrossRef]
- Corrigall, K.A.; Trainor, L.J. Associations Between Length of Music Training and Reading Skills in Children. Music Percept. 2011, 29, 147–155. [Google Scholar] [CrossRef]
- Roden, I.; Früchtenicht, K.; Kreutz, G.; Linderkamp, F.; Grube, D. Auditory Stimulation Training With Technically Manipulated Musical Material in Preschool Children With Specific Language Impairments: An Explorative Study. Front. Psychol. 2019, 10, 2026. [Google Scholar] [CrossRef]
- White, E.J.; Hutka, S.A.; Williams, L.J.; Moreno, S. Learning, neural plasticity and sensitive periods: Implications for language acquisition, music training and transfer across the lifespan. Front. Syst. Neurosci. 2013, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Brandt, A.K.; Slevc, R.; Gebrian, M. Music and Early Language Acquisition. Front. Psychol. 2012, 3, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushnerenko, E.; Ceponiene, R.; Balan, P.; Fellman, V.; Huotilaine, M.; Näätäne, R. Maturation of the auditory event-related potentials during the first year of life. Neuroreport 2002, 13, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Morr, M.L.; Shafer, V.L.; Kreuzer, J.A.; Kurtzberg, D. Maturation of mismatch negativity in typically developing infants and preschool children. Ear Hear. 2002, 23, 118–136. [Google Scholar] [CrossRef] [PubMed]
- Dehaene-Lambertz, G. The human infant brain: A neural architecture able to learn language. Psychon. Bull. Rev. 2017, 24, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Minagawa-Kawai, Y.; Cristià, A.; Dupoux, E. Cerebral lateralization and early speech acquisition: A developmental scenario. Dev. Cogn. Neurosci. 2011, 1, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Musacchia, G.; Choudhury, N.A.; Ortiz-Mantilla, S.; Realpe-Bonilla, T.; Roesler, C.P.; Benasich, A.A. Oscillatory support for rapid frequency change processing in infants. Neuropsychologia 2013, 51, 2812–2824. [Google Scholar] [CrossRef]
- Musacchia, G.; Ortiz-Mantilla, S.; Realpe-Bonilla, T.; Roesler, C.P.; Benasich, A.A. Infant Auditory Processing and Event-related Brain Oscillations. J. Vis. Exp. 2015, 2015, 52420. [Google Scholar] [CrossRef] [Green Version]
- Kirschner, S.; Tomasello, M. Joint drumming: Social context facilitates synchronization in preschool children. J. Exp. Child Psychol. 2009, 102, 299–314. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | ||||
| RTr+ (N = 15) | RTr− (N = 14) | t (df) | p | |
| Socioeconomic status a | 68.00 (16.56) | 70.00 (17.97) | 0.312 (27) | 0.758 |
| Maternal education level b | 63.33 (15.89) | 64.29 (10.16) | 0.191 (27) | 0.850 |
| Paternal education level b | 55.00 (13.76) | 50.71 (19.79) | −0.681 (27) | 0.502 |
| B | ||||
| RTr+ Subsample with ERP Data (N = 11) | RTr− Sample with ERP Data (N = 14) | t (df) | p | |
| Socioeconomic status a | 58.18 (17.22) | 66.07 (12.43) | 0.164 (23) | 0.196 |
| Maternal education level b | 59.09 (18.14) | 59.29 (13.28) | 0.031 (23) | 0.976 |
| Paternal education level b | 51.36 (13.43) | 55.38 (8.77) | 0.881 (23) | 0.388 |
| (A) Bayley Language Scale—Receptive Communication Subtest | ||||
| RTr− | RTr+ | t (27) | p | |
| Pre-training | 11.46 (1.34) | 11.60 (0.74) | −0.335 | 0.735 |
| Post-training | 14.86 (1.23) | 16.40 (2.23) | −2.283 | 0.031 |
| (B) Bayley Language Scale—Expressive Communication Subtest | ||||
| RTr− | RTr+ | t (27) | p | |
| Pre-training | 10.32 (1.64) | 10.00 (1.54) | 0.546 | 0.590 |
| Post-training | 15.79 (1.63) | 17.00 (1.93) | −1.827 | 0.079 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dondena, C.; Riva, V.; Molteni, M.; Musacchia, G.; Cantiani, C. Impact of Early Rhythmic Training on Language Acquisition and Electrophysiological Functioning Underlying Auditory Processing: Feasibility and Preliminary Findings in Typically Developing Infants. Brain Sci. 2021, 11, 1546. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11111546
Dondena C, Riva V, Molteni M, Musacchia G, Cantiani C. Impact of Early Rhythmic Training on Language Acquisition and Electrophysiological Functioning Underlying Auditory Processing: Feasibility and Preliminary Findings in Typically Developing Infants. Brain Sciences. 2021; 11(11):1546. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11111546
Chicago/Turabian StyleDondena, Chiara, Valentina Riva, Massimo Molteni, Gabriella Musacchia, and Chiara Cantiani. 2021. "Impact of Early Rhythmic Training on Language Acquisition and Electrophysiological Functioning Underlying Auditory Processing: Feasibility and Preliminary Findings in Typically Developing Infants" Brain Sciences 11, no. 11: 1546. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11111546