
Online Movement Correction in Response to the Unexpectedly Perturbed Initial or Final Action Goals: An ERP and sLORETA Study


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Method
2.1. Participants
2.2. Apparatus and Stimuli
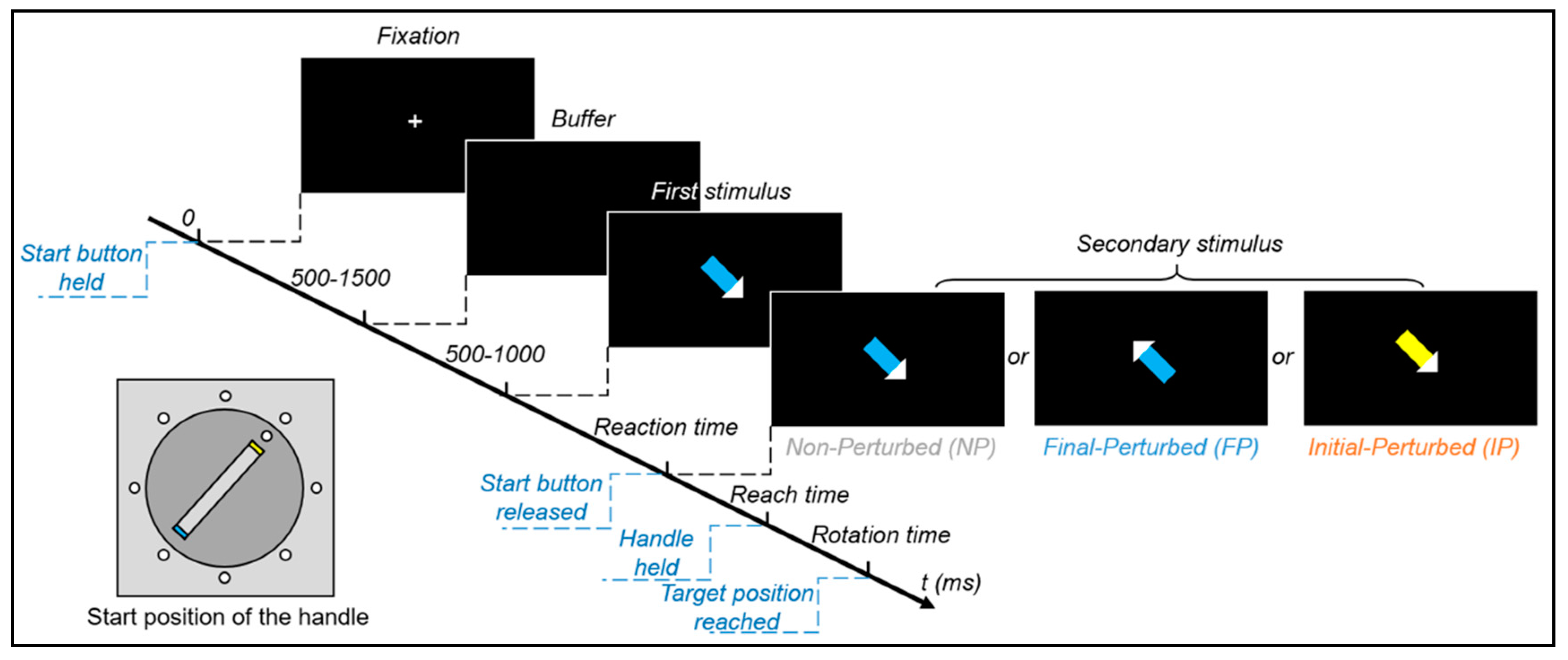
2.3. Experimental Paradigm
2.4. Procedure
2.5. Behavioral and Electrophysiological Recordings
2.6. Data Analysis
2.6.1. Behavioral Data
2.6.2. ERPs
2.6.3. Source Analysis
3. Results
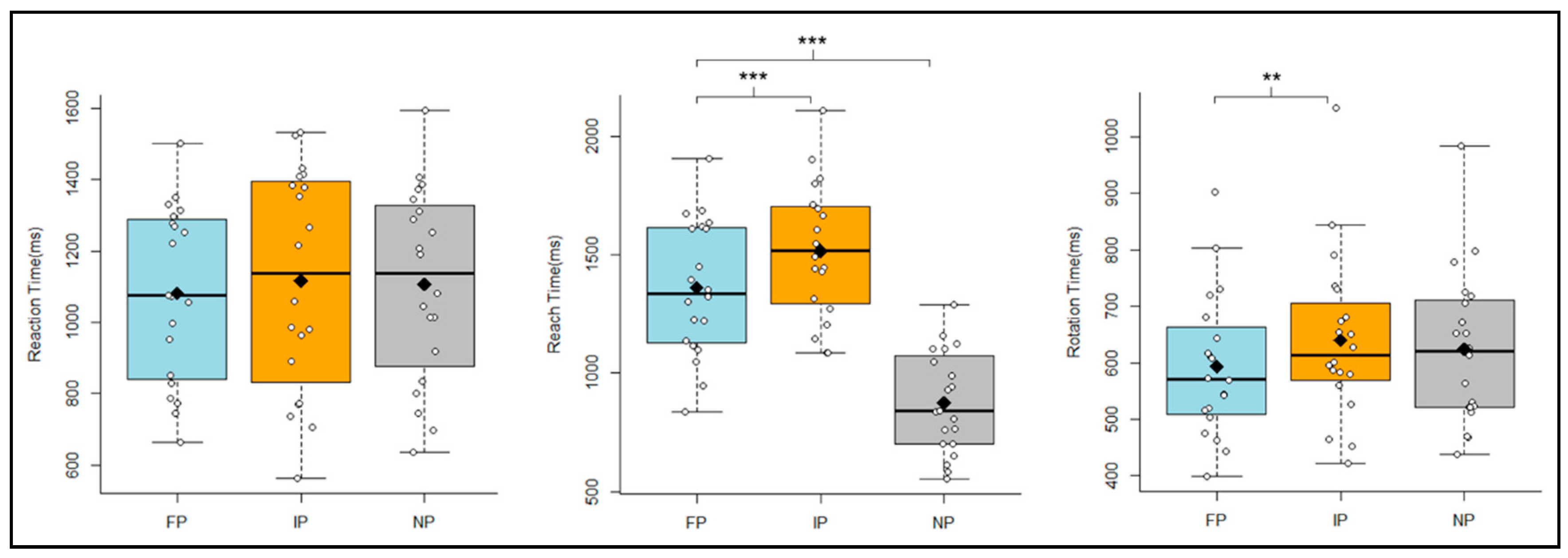
3.1. Subjective Difficulty Ratings
3.2. Timing
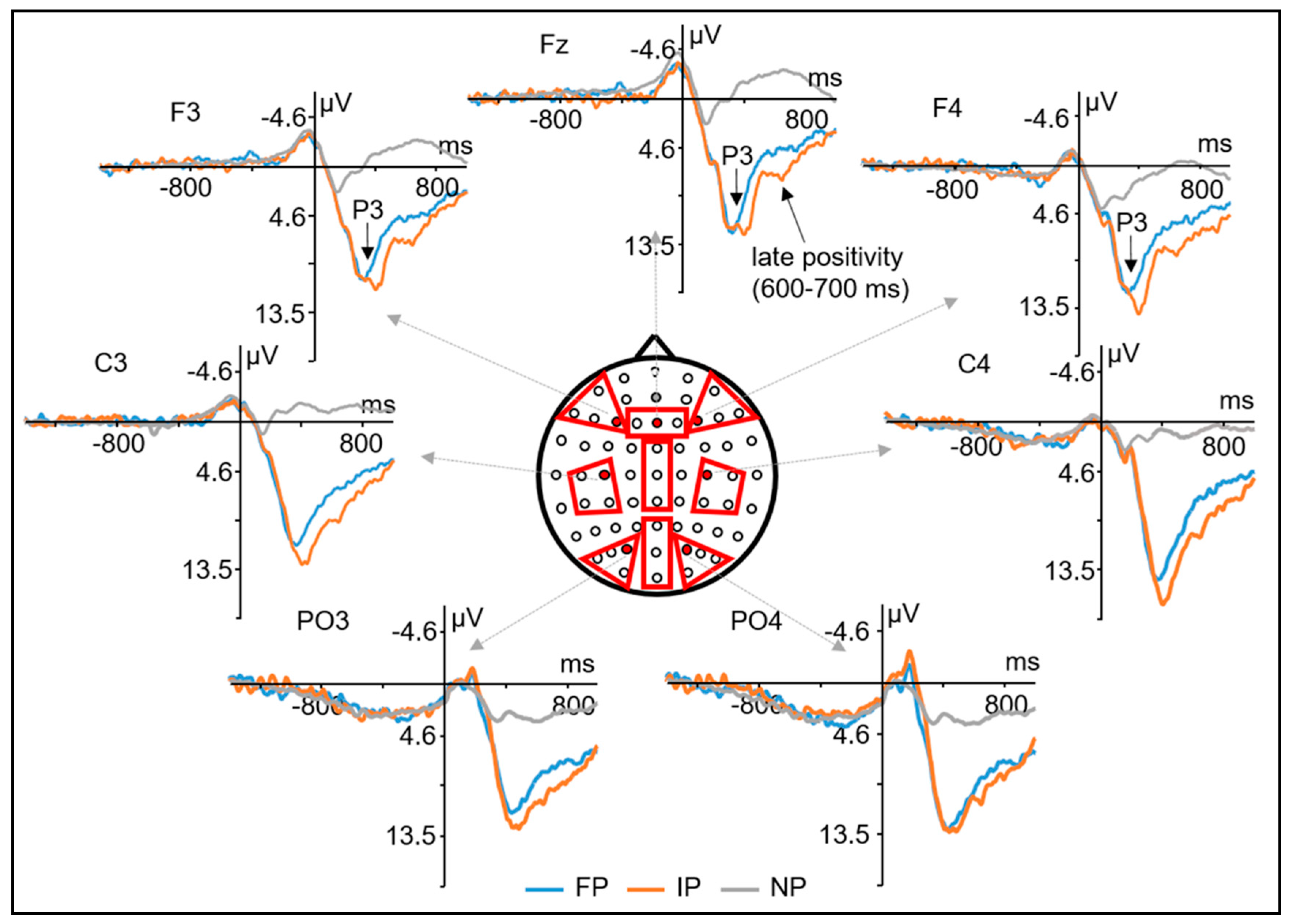
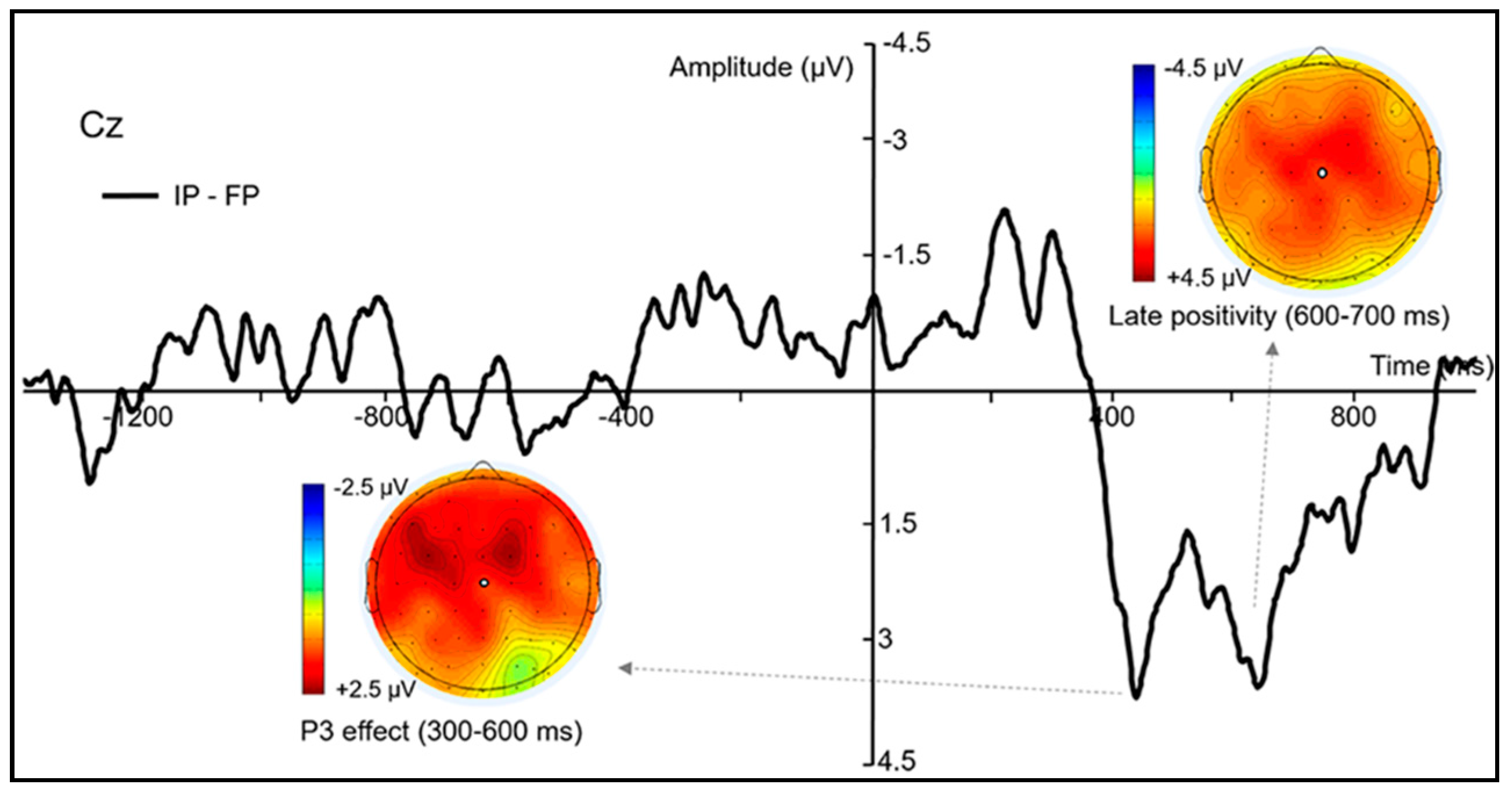
3.3. ERP Results
3.3.1. Epoch Time-Locked to the Secondary Stimulus
3.3.2. Epoch Time-Locked to Grasping
3.4. Source Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tunik, E.; Frey, S.H.; Grafton, S.T. Virtual lesions of the anterior intraparietal area disrupt goal-dependent on-line adjustments of grasp. Nat. Neurosci. 2005, 8, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.T.; Sherwood, D.E. Time requirements of changes in program and parameter variables in rapid ongoing movements. J. Mot. Behav. 1983, 15, 163–178. [Google Scholar] [CrossRef]
- Spiegel, M.A.; Koester, D.; Schack, T. The functional role of working memory in the (re-)planning and execution of grasping movements. J. Exp. Psychol. Hum. Percept. Perform. 2013, 39, 1326–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkataramani, P.V.; Gopal, A.; Murthy, A. An independent race model involving an abort and re-plan strategy explains reach redirecting movements during planning and execution. Eur. J. Neurosci. 2018, 47, 460–478. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.A.; Dos Santos Lima, E.; Marília Franzoni, M. The continuous nature of timing reprogramming in an interceptive task. J. Sports Sci. 2005, 23, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.A.; Chua, R.; Nagelkerke, P.; Franks, I.M. Reprogramming of interceptive actions: Time course of temporal corrections for unexpected target velocity change. J. Mot. Behav. 2006, 38, 467–477. [Google Scholar] [CrossRef]
- Ikudome, S.; Nakamoto, H.; Yotani, K.; Unenaka, S.; Mori, S. Potential of rapid adjustment of brief interceptive action using predicted information. Brain Cogn. 2015, 97, 51–58. [Google Scholar] [CrossRef]
- Tunik, E.; Ortigue, S.; Adamovich, S.V.; Grafton, S.T. Differential recruitment of anterior intraparietal sulcus and superior parietal lobule during visually guided grasping revealed by electrical neuroimaging. J. Neurosci. 2008, 28, 13615–13620. [Google Scholar] [CrossRef]
- Iannaccone, R.; Hauser, T.U.; Staempfli, P.; Walitza, S.; Brandeis, D.; Brem, S. Conflict monitoring and error processing: New insights from simultaneous EEG–fMRI. NeuroImage 2015, 105, 395–407. [Google Scholar] [CrossRef]
- Neubert, F.X.; Mars, R.B.; Buch, E.R.; Olivier, E.; Rushworth, M.F.S. Cortical and subcortical interactions during action reprogramming and their related white matter pathways. Proc. Natl. Acad. Sci. USA 2010, 107, 13240–13245. [Google Scholar] [CrossRef] [Green Version]
- Hartwigsen, G.; Bestmann, S.; Ward, N.S.; Woerbel, S.; Mastroeni, C.; Granert, O.; Siebner, H.R. Left dorsal premotor cortex and supramarginal gyrus complement each other during rapid action reprogramming. J. Neurosci. 2012, 32, 16162–16171. [Google Scholar] [CrossRef] [Green Version]
- Ullsperger, M.; Danielmeier, C.; Jocham, G. Neurophysiology of Performance Monitoring and Adaptive Behavior. Physiol. Rev. 2014, 94, 35–79. [Google Scholar] [CrossRef] [PubMed]
- Tunik, E.; Rice, N.J.; Hamilton, A.; Grafton, S.T. Beyond grasping: Representation of action in human anterior intraparietal sulcus. NeuroImage 2007, 36, T77–T86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buch, E.R.; Mars, R.B.; Boorman, E.D.; Rushworth, M.F.S. A network centered on ventral premotor cortex exerts both facilitatory and inhibitory control over primary motor cortex during action reprogramming. J. Neurosci. 2010, 30, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, H.; Ikudome, S.; Yotani, K.; Maruyama, A.; Mori, S. Fast-ball sports experts depend on an inhibitory strategy to reprogram their movement timing. Exp. Brain Res. 2013, 228, 193–203. [Google Scholar] [CrossRef]
- Hartwigsen, G.; Siebner, H.R. Joint contribution of left dorsal premotor cortex and supramarginal gyrus to rapid action reprogramming. Brain Stimul. 2015, 8, 945–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mars, R.B.; Piekema, C.; Coles, M.G.H.; Hulstijn, W.; Toni, I. On the programming and reprogramming of actions. Cereb. Cortex 2007, 17, 2972–2979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Ulbert, I.; Schomer, D.L.; Marinkovic, K.; Halgren, E. Responses of human anterior cingulate cortex microdomains to error detection, conflict monitoring, stimulus-response mapping, familiarity, and orienting. J. Neurosci. 2005, 25, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Kropotov, J.D.; Ponomarev, V.A.; Hollup, S.; Mueller, A. Dissociating action inhibition, conflict monitoring and sensory mismatch into independent components of event related potentials in GO/NOGO task. NeuroImage 2011, 57, 565–575. [Google Scholar] [CrossRef]
- Leuthold, H.; Jentzsch, I. Spatiotemporal source localisation reveals involvement of medial premotor areas in movement reprogramming. Exp. Brain Res. 2002, 144, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Vidal, F.; Bonnet, M.; Macar, F. Programming the duration of a motor sequence: Role of the primary and supplementary motor areas in man. Exp. Brain Res. 1995, 106, 339–350. [Google Scholar] [CrossRef]
- Lee, H.W.; Lu, M.S.; Chen, C.Y.; Muggleton, N.G.; Hsu, T.Y.; Juan, C.H. Roles of the pre-SMA and rIFG in conditional stopping revealed by transcranial magnetic stimulation. Behav. Brain Res. 2016, 296, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Tunik, E.; Lo, O.-Y.; Adamovich, S.V. Transcranial magnetic stimulation to the frontal operculum and supramarginal gyrus disrupts planning of outcome-based hand-object interactions. J. Neurosci. 2008, 28, 14422–14427. [Google Scholar] [CrossRef] [PubMed]
- Ward, N.S.; Bestmann, S.; Hartwigsen, G.; Weiss, M.M.; Christensen, L.O.D.; Frackowiak, R.S.J.; Rothwell, J.C.; Siebner, H.R. Low-Frequency Transcranial Magnetic Stimulation over Left dorsal premotor cortex improves the dynamic control of visuospatially cued actions. J. Neurosci. 2010, 30, 9216–9223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culham, J.C.; Cavina-Pratesi, C.; Singhal, A. The role of parietal cortex in visuomotor control: What have we learned from neuroimaging? Neuropsychologia 2006, 44, 2668–2684. [Google Scholar] [CrossRef] [PubMed]
- Rice, N.J.; Tunik, E.; Grafton, S.T. The anterior intraparietal sulcus mediates grasp execution, independent of requirement to update: New insights from transcranial magnetic stimulation. J. Neurosci. 2006, 26, 8176–8182. [Google Scholar] [CrossRef] [Green Version]
- Striemer, C.L.; Chouinard, P.A.; Goodale, M.A. Programs for action in superior parietal cortex: A triple-pulse TMS investigation. Neuropsychologia 2011, 49, 2391–2399. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, F.; Aron, A.R.; Stevens, M.A.; Chambers, C.D. Theta burst stimulation dissociates attention and action updating in human inferior frontal cortex. Proc. Natl. Acad. Sci. USA 2010, 107, 13966–13971. [Google Scholar] [CrossRef] [Green Version]
- Pellegrino, G.; Tomasevic, L.; Herz, D.M.; Larsen, K.M.; Siebner, H.R. Theta Activity in the Left Dorsal Premotor Cortex During Action Re-Evaluation and Motor Reprogramming. Front. Hum. Neurosci. 2018, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Castiello, U.; Bennett, K.M.B.; Paulignan, Y. Does the type of prehension influence the kinematics of reaching. Behav. Brain Res. 1992, 50, 7–15. [Google Scholar] [CrossRef]
- Paulignan, Y.; MacKenzie, C.; Marteniuk, R.; Jeannerod, M. Selective perturbation of visual input during prehension movements—1. The effects of changing object position. Exp. Brain Res. 1991, 83, 502–512. [Google Scholar] [CrossRef]
- Paulignan, Y.; Jeannerod, M.; MacKenzie, C.; Marteniuk, R. Selective perturbation of visual input during prehension movements—2. The effects of changing object size. Exp. Brain Res. 1991, 87, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Castiello, U.; Bennett, K.; Stelmach, G.E. Reach to grasp: The natural response to perturbation of object size. Exp. Brain Res. 1993, 94, 163–178. [Google Scholar] [CrossRef]
- Castiello, U.; Bennett, K.M.B.; Chambers, H. Reach to grasp: The response to a simultaneous perturbation of object position and size. Exp. Brain Res. 1998, 120, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.M.L.; Seegelke, C.; Spiegel, M.A.; Oehmichen, C.; Hammes, J.; Schack, T. Corrections in grasp posture in response to modifications of action goals. PLoS ONE 2012, 7, e43015. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.M.L.; Seegelke, C. Perturbations in Action Goal Influence Bimanual Grasp Posture Planning. J. Mot. Behav. 2013, 45, 473–478. [Google Scholar] [CrossRef]
- Rosenbaum, D.A.; Cohen, R.G.; Jax, S.A.; Weiss, D.J.; van der Wel, R. The problem of serial order in behavior: Lashley’s legacy. Hum. Mov. Sci. 2007, 26, 525–554. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, D.A.; Vaughan, J.; Barnes, H.J.; Jorgensen, M.J. Time course of movement planning: Selection of handgrips for object manipulation. J. Exp. Psychol. Learn. Mem. Cogn. 1992, 18, 1058–1073. [Google Scholar] [CrossRef]
- Westerholz, J.; Schack, T.; Koester, D. Event-related brain potentials for goal-related power grips. PLoS ONE 2013, 8, e68501. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Schack, T.; Koester, D. Coordinating initial and final action goals in planning grasp-to-rotate movements: An ERP study. Neuroscience 2021, 459, 70–84. [Google Scholar] [CrossRef]
- Cohen, R.G.; Rosenbaum, D.A. Where grasps are made reveals how grasps are planned: Generation and recall of motor plans. Exp. Brain Res. 2004, 157, 486–495. [Google Scholar] [CrossRef]
- Golob, E.J.; Pratt, H.; Starr, A. Preparatory slow potentials and event-related potentials in an auditory cued attention task. Clin. Neurophysiol. 2002, 113, 1544–1557. [Google Scholar] [CrossRef] [Green Version]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Calderon, J.; Luck, S.J. ERPLAB: An open-source toolbox for the analysis of event-related potentials. Front. Hum. Neurosci. 2014, 8, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratton, G.; Coles, M.G.H.; Donchin, E. A new method for off-line removal of ocular artifact. Electroencephalogr. Clin. Neurophysiol. 1983, 55, 468–484. [Google Scholar] [CrossRef]
- Miller, G.A.; Gration, G.; Yee, C.M. Generalized implementation of an eye movement correction procedure. Psychophysiology 1988, 25, 241–243. [Google Scholar] [CrossRef]
- Van Schie, H.T.; Bekkering, H. Neural mechanisms underlying immediate and final action goals in object use reflected by slow wave brain potentials. Brain Res. 2007, 1148, 183–197. [Google Scholar] [CrossRef]
- Westerholz, J.; Schack, T.; Schutz, C.; Koester, D. Habitual vs non-habitual manual actions: An ERP study on overt movement execution. PLoS ONE 2014, 9, e93116. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Pascual-Marqui, R.D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods Find. Exp. Clin. Pharmacol. 2002, 24, 5–12. [Google Scholar]
- Fuchs, M.; Kastner, J.; Wagner, M.; Hawes, S.; Ebersole, J.S. A standardized boundary element method volume conductor model. Clin. Neurophysiol. 2002, 113, 702–712. [Google Scholar] [CrossRef] [Green Version]
- Mazziotta, J.C.; Toga, A.; Evans, A.; Fox, P.; Lancaster, J.; Zilles, K.; Woods, R.; Paus, T.; Simpson, G.; Pike, B.; et al. A probabilistic atlas and reference system for the human brain: International Consortium for Brain Mapping (ICBM). Philos. Trans. R. Soc. B Biol. Sci. 2001, 356, 1293–1322. [Google Scholar] [CrossRef] [PubMed]
- Nichols, T.; Holmes, A. Nonparametric permutation tests for functional neuroimaging. Hum. Brain Mapp. 2002, 15, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botwinick, J.; Thompson, L.W. Premotor and motor components of reaction time. J. Exp. Psychol. 1966, 71, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, D.; Hansen, S.; Grierson, L.E.M.; Lyons, J.; Bennett, S.J.; Hayes, S.J. Goal-directed aiming: Two components but multiple processes. Psychol. Bull. 2010, 136, 1023–1044. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, U.M.; Knight, R.T.; Muente, T.F. Electrophysiological evidence for different inhibitory mechanisms when stopping or changing a planned response. J. Cogn. Neurosci. 2011, 23, 2481–2493. [Google Scholar] [CrossRef] [PubMed]
- Recio, G.; Shmuilovich, O.; Sommer, W. Should I smile or should I frown? An ERP study on the voluntary control of emotion-related facial expressions. Psychophysiology 2014, 51, 789–799. [Google Scholar] [CrossRef]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comerchero, M.D.; Polich, J. P3a and P3b from typical auditory and visual stimuli. Clin. Neurophysiol. 1999, 110, 24–30. [Google Scholar] [CrossRef]
- Gajewski, P.D.; Falkenstein, M. Effects of task complexity on ERP components in Go/Nogo tasks. Int. J. Psychophysiol. 2013, 87, 273–278. [Google Scholar] [CrossRef]
- Benikos, N.; Johnstone, S.J.; Roodenrys, S.J. Varying task difficulty in the Go/Nogo task: The effects of inhibitory control, arousal, and perceived effort on ERP components. Int. J. Psychophysiol. 2013, 87, 262–272. [Google Scholar] [CrossRef] [Green Version]
- Falkenstein, M.; Hoormann, J.; Hohnsbein, J. ERP components in Go/Nogo tasks and their relation to inhibition. Acta Psychol. 1999, 101, 267–291. [Google Scholar] [CrossRef]
- Kok, A. Effects of degradation of visual stimuli on components of the event-related potential (ERP) in go/nogo reaction tasks. Biol. Psychol. 1986, 23, 21–38. [Google Scholar] [CrossRef]
- Eimer, M. Effects of attention and stimulus probability on ERPs in a Go/Nogo task. Biol. Psychol. 1993, 35, 123–138. [Google Scholar] [CrossRef]
- Falkenstein, M.; Koshlykova, N.A.; Kiroj, V.N.; Hoormann, J.; Hohnsbein, J. Late ERP components in visual and auditory Go/Nogo tasks. Electroencephalogr. Clin. Neurophysiol. Evoked Potentials 1995, 96, 36–43. [Google Scholar] [CrossRef]
- Albert, J.; López-Martín, S.; Hinojosa, J.A.; Carretié, L. Spatiotemporal characterization of response inhibition. NeuroImage 2013, 76, 272–281. [Google Scholar] [CrossRef]
- Boecker, M.; Gauggel, S.; Drueke, B. Stop or stop-change—Does it make any difference for the inhibition process? Int. J. Psychophysiol. 2013, 87, 234–243. [Google Scholar] [CrossRef]
- Smith, J.L.; Johnstone, S.J.; Barry, R.J. Movement-related potentials in the Go/NoGo task: The P3 reflects both cognitive and motor inhibition. Clin. Neurophysiol. 2008, 119, 704–714. [Google Scholar] [CrossRef]
- Oldenburg, J.F.E.; Roger, C.; Assecondi, S.; Verbruggen, F.; Fias, W. Repetition priming in the stop signal task: The electrophysiology of sequential effects of stopping. Neuropsychologia 2012, 50, 2860–2868. [Google Scholar] [CrossRef]
- Wessel, J.R.; Aron, A.R. It’s not too late: The onset of the frontocentral P3 indexes successful response inhibition in the stop-signal paradigm. Psychophysiology 2015, 52, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Ramautar, J.R.; Kok, A.; Ridderinkhof, K.R. Effects of stop-signal modality on the N2/P3 complex elicited in the stop-signal paradigm. Biol. Psychol. 2006, 72, 96–109. [Google Scholar] [CrossRef]
- Dimoska, A.; Johnstone, S.J.; Barry, R.J. The auditory-evoked N2 and P3 components in the stop-signal task: Indices of inhibition, response-conflict or error-detection? Brain Cogn. 2006, 62, 98–112. [Google Scholar] [CrossRef]
- Ramautar, J.R.; Kok, A.; Ridderinkhof, K.R. Effects of stop-signal probability in the stop-signal paradigm: The N2/P3 complex further validated. Brain Cogn. 2004, 56, 234–252. [Google Scholar] [CrossRef]
- Hong, X.; Wang, Y.; Sun, J.; Li, C.; Tong, S. Segregating Top-Down Selective Attention from Response Inhibition in a Spatial Cueing Go NoGo Task: An ERP and Source Localization Study. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huster, R.J.; Westerhausen, R.; Pantev, C.; Konrad, C. The role of the cingulate cortex as neural generator of the N200 and P300 in a tactile response inhibition task. Hum. Brain Mapp. 2009, 31, 1260–1271. [Google Scholar] [CrossRef]
- Rae, C.L.; Hughes, L.E.; Weaver, C.; Anderson, M.C.; Rowe, J.B. Selection and stopping in voluntary action: A meta-analysis and combined fMRI study. NeuroImage 2014, 86, 381–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubia, K.; Russell, T.; Overmeyer, S.; Brammer, M.J.; Bullmore, E.T.; Sharma, T.; Simmons, A.; Williams, S.C.R.; Giampietroa, V.; Andrew, C.M.; et al. Mapping motor inhibition: Conjunctive brain activations across different versions of Go/No-Go and stop tasks. NeuroImage 2001, 13, 250–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasi, G.; Goldberg, T.E.; Weickert, T.; Das, S.; Kohn, P.; Zoltick, B.; Bertolino, A.; Callicott, J.H.; Weinberger, D.R.; Mattay, V.S. Brain regions underlying response inhibition and interference monitoring and suppression. Eur. J. Neurosci. 2006, 23, 1658–1664. [Google Scholar] [CrossRef]
- Sebastian, A.; Pohl, M.F.; Klöppel, S.; Feige, B.; Lange, T.; Stahl, C.; Voss, A.; Klauer, K.C.; Lieb, K.; Tüscher, O. Disentangling common and specific neural subprocesses of response inhibition. NeuroImage 2013, 64, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Brass, M.; Zysset, S.; von Cramon, D.Y. The inhibition of imitative response tendencies. NeuroImage 2001, 14, 1416–1423. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.R.G.; Vilis, T.; Everling, S. Isolation of saccade inhibition processes: Rapid event-related fMRI of saccades and nogo trials. NeuroImage 2008, 39, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Kadota, H.; Sekiguchi, H.; Takeuchi, S.; Miyazaki, M.; Kohno, Y.; Nakajima, Y. The role of the dorsolateral prefrontal cortex in the inhibition of stereotyped responses. Exp. Brain Res. 2010, 203, 593–600. [Google Scholar] [CrossRef]
- Balconi, M.; Vitaloni, S. The tDCS effect on alpha brain oscillation for correct vs. incorrect object use. The contribution of the left DLPFC. Neurosci. Lett. 2012, 517, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.H. Tool use, communicative gesture and cerebral asymmetries in the modern human brain. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1951–1957. [Google Scholar] [CrossRef] [PubMed]
- Haaland, K.Y.; Harrington, D.L.; Knight, R.T. Neural representations of skilled movement. Brain 2000, 123, 2306–2313. [Google Scholar] [CrossRef] [Green Version]
- Martin, K.; Jacobs, S.; Frey, S.H. Handedness-dependent and -independent cerebral asymmetries in the anterior intraparietal sulcus and ventral premotor cortex during grasp planning. NeuroImage 2011, 57, 502–512. [Google Scholar] [CrossRef] [Green Version]
- Janssen, L.; Beuting, M.; Meulenbroek, R.G.J.; Steenbergen, B. Combined effects of planning and execution constraints on bimanual task performance. Exp. Brain Res. 2009, 192, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Janssen, L.; Meulenbroek, R.G.J.; Steenbergen, B. Behavioral evidence for left-hemisphere specialization of motor planning. Exp. Brain Res. 2011, 209, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Mayer, A.R.; Hanlon, F.M.; Shaff, N.A.; Stephenson, D.D.; Ling, J.M.; Dodd, A.B.; Hogeveen, J.; Quinn, D.K.; Ryman, S.G.; Pirio-Richardson, S. Evidence for asymmetric inhibitory activity during motor planning phases of sensorimotor synchronization. Cortex 2020, 129, 314–328. [Google Scholar] [CrossRef] [PubMed]
- Kaan, E.; Harris, A.; Gibson, E.; Holcomb, P. The P600 as an index of syntactic integration difficulty. Lang. Cogn. Process. 2000, 15, 159–201. [Google Scholar] [CrossRef]
- Hagoort, P.; Brown, C.M. ERP effects of listening to speech compared to reading: The P600/SPS to syntactic violations in spoken sentences and rapid serial visual presentation. Neuropsychologia 2000, 38, 1531–1549. [Google Scholar] [CrossRef] [Green Version]
- Frisch, S.; Kotz, S.A.; Von Cramon, D.Y.; Friederici, A.D. Why the P600 is not just a P300: The role of the basal ganglia. Clin. Neurophysiol. 2003, 114, 336–340. [Google Scholar] [CrossRef] [Green Version]
- van Herten, M.; Kolk, H.H.J.; Chwilla, D.J. An ERP study of P600 effects elicited by semantic anomalies. Cogn. Brain Res. 2005, 22, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Bach, P.; Gunter, T.C.; Knoblich, G.; Prinz, W.; Friederici, A.D. N400-like negativities in action perception reflect the activation of two components of an action representation. Soc. Neurosci. 2009, 4, 212–232. [Google Scholar] [CrossRef]
- Gouvea, A.C.; Phillips, C.; Kazanina, N.; Poeppel, D. The linguistic processes underlying the P600. Lang. Cogn. Process. 2010, 25, 149–188. [Google Scholar] [CrossRef]
- Osterhout, L.; Holcomb, P.J. Event-related brain potentials elicited by syntactic anomaly. J. Mem. Lang. 1992, 31, 785–806. [Google Scholar] [CrossRef]
- Maffongelli, L.; Bartoli, E.; Sammler, D.; Kölsch, S.; Campus, C.; Olivier, E.; Fadiga, L.; D’Ausilio, A. Distinct brain signatures of content and structure violation during action observation. Neuropsychologia 2015, 75, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Kuperberg, G.R. Neural mechanisms of language comprehension: Challenges to syntax. Brain Res. 2007, 1146, 23–49. [Google Scholar] [CrossRef]
- Koelsch, S.; Gunter, T.C.; Wittfoth, M.; Sammler, D. Interaction between syntax processing in language and in music: An ERP study. J. Cogn. Neurosci. 2005, 17, 1565–1577. [Google Scholar] [CrossRef]
- Patel, A.D.; Gibson, E.; Ratner, J.; Besson, M.; Holcomb, P.J. Processing syntactic relations in language and music: An event-related potential study. J. Cogn. Neurosci. 1998, 10, 717–733. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Schack, T.; Koester, D. Online Movement Correction in Response to the Unexpectedly Perturbed Initial or Final Action Goals: An ERP and sLORETA Study. Brain Sci. 2021, 11, 641. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11050641
Yu L, Schack T, Koester D. Online Movement Correction in Response to the Unexpectedly Perturbed Initial or Final Action Goals: An ERP and sLORETA Study. Brain Sciences. 2021; 11(5):641. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11050641
Chicago/Turabian StyleYu, Lin, Thomas Schack, and Dirk Koester. 2021. "Online Movement Correction in Response to the Unexpectedly Perturbed Initial or Final Action Goals: An ERP and sLORETA Study" Brain Sciences 11, no. 5: 641. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11050641