
Effect of Allopregnanolone on Spatial Memory and Synaptic Proteins in Animal Model of Metabolic Syndrome

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Induction of Metabolic Syndrome
2.3. Experimental Design
2.4. Assessment of Spatial Working Memory
2.4.1. Y-Maze Task
2.4.2. Apparatus
2.5. In Vivo Measurement of the Arterial Blood Pressure
2.6. Biochemical Measurements
2.7. Gene Expression by Real-Time PCR of Synaptophysin and Growth Associated Protein 43 (GAP-43)
2.8. Statistical Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kassi, E.; Pervanidou, P.; Kaltsas, G.; Chrousos, G. Metabolic syndrome: Definitions and controversies. BMC Med. 2011, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Yates, K.F.; Sweat, V.; Yau, P.L.; Turchiano, M.M.; Convit, A. Impact of metabolic syndrome on cognition and brain: A selected review of the literature. Arter. Thromb. Vasc. Biol. 2012, 32, 2060–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genazzani, A.D.; Strucchi, C.; Luisi, M.; Casarosa, E.; Lanzoni, C.; Baraldi, E.; Ricchieri, F.; Mehmeti, H.; Genazzani, A.R. Metformin administration modulates neurosteroids secretion in non-obese amenorrhoic patients with polycystic ovary syndrome. Gynecol. Endocrinol. 2006, 22, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Head, G.A.; Jackson, K.L.; Gueguen, C. Potential Therapeutic Use of Neurosteroids for Hypertension. Front. Physiol. 2019, 10, 1477. [Google Scholar] [CrossRef] [Green Version]
- Lloyd-Evans, E.; Waller-Evans, H. Biosynthesis and signalling functions of central and peripheral nervous system neurosteroids in health and disease. Essays Biochem. 2020, 64, 591–606. [Google Scholar] [CrossRef] [PubMed]
- Parks, E.E.; Logan, S.; Yeganeh, A.; Farley, J.A.; Owen, D.B.; Sonntag, W.E. Interleukin 6 reduces allopregnanolone synthesis in the brain and contributes to age-related cognitive decline in mice. J. Lipid Res. 2020, 61, 1308–1319. [Google Scholar] [CrossRef] [PubMed]
- Noorbakhsh, F.; Baker, G.B.; Power, C. Allopregnanolone and neuroinflammation: A focus on multiple sclerosis. Front. Cell. Neurosci. 2014, 8, 134. [Google Scholar] [CrossRef] [Green Version]
- Gajda, A.; Pellizzon, M.; Ricci, M.; Ulman, E. Diet-Induced Metabolic Syndrome in Rodent Models. Animal Lab News. 2007. Available online: http://www.eps-cjgroup.com/lsg/service/researchdiets/pdf/Diet-Induced%20Metabolic%20Syndrome%20models.pdf (accessed on 1 September 2020).
- Nagai, Y.; Yonemitsu, S.; Erion, D.M.; Iwasaki, T.; Stark, R.; Weismann, D.; Dong, J.; Zhang, D.; Jurczak, M.J.; Löffler, M.G.; et al. The role of peroxisome proliferator-activated receptor gamma coactivator-1 beta in the pathogenesis of fructose-induced insulin resistance. Cell Metab. 2009, 9, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Samuelsson, A.M.; Matthews, P.A.; Argenton, M.; Christie, M.R.; McConnell, J.M.; Jansen, E.H. Diet-induced obesity in female mice leads to offspring hyperphagia, adiposity, hypertension, and insulin resistance: A novel murine model of developmental programming. Hypertension 2008, 51, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Maguire, J.; Mody, I. GABAAR plasticity during pregnancy: Relevance to postpartum depression. Neuron 2008, 59, 207–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afrazi, S.; Esmaeili-Mahani, S.; Sheibani, V.; Abbasnejad, M. Neurosteroid allopregnanolone attenuates high glucose-induced apoptosis and prevents experimental diabetic neuropathic pain: In vitro and in vivo studies. J. Steroid. Biochem. Mol. Biol. 2014, 139, 98–103. [Google Scholar] [CrossRef]
- Irvine, R.J.; White, J.; Chan, R. The influence of restraint on blood pressure in the rat. J. Pharmacol. Toxicol. 1997, 38, 157–162. [Google Scholar] [CrossRef]
- Passing, H.; Bablok, W. A new biometrical procedure for testing the equality of measurements from two different analytical methods. Application of linear regression procedures for method comparison studies in clinical chemistry, Part I. J. Clin. Chem. Clin. Biochem. 1983, 21, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Becker, R.H.A.; Frick, A.D.; Burger, F.; Potgieter, J.H.; Scholtz, H. Insulin glulisine, a new rapid-acting insulin analogue, displays a rapid time-action profile in obese non-diabetic subjects. Exp. Clin. Endocrinol. Diabetes 2005, 113, 435–443. [Google Scholar] [CrossRef]
- Allain, C.C.; Poon, L.S.; Chan, C.S.; Richmond, W.; Fu, P.C. Enzymatic determination of total serum cholesterol. Clin. Chem. 1974, 20, 470–475. [Google Scholar] [CrossRef]
- Glick, M.R.; Ryder, K.W.; Jackson, S.A. Graphical comparisons of interferences in clinical chemistry instrumentation. Clin. Chem. 1986, 32, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Lopez-virella, M.F.; Stone, P.; Ellis, S.; Collwel, J. Cholesterol determination in high density lipoprotein by three different methods. Clin. Chem. 1977, 23, 882. [Google Scholar] [CrossRef]
- Treviño, S.; Vázquez-Roque, R.A.; López-López, G.; Perez-Cruz, C.; Moran, C.; Handal-Silva, A.; González-Vergara, E.; Flores, G.; Guevara, J.; Díaz, A. Metabolic syndrome causes recognition impairments and reduced hippocampal neuronal plasticity in rats. J. Chem. Neuroanat. 2017, 82, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Haberman, R.P.; Colantuoni, C.; Koh, M.T.; Gallagher, M. Behaviorally activated mRNA expression profiles produce signatures of learning and enhanced inhibition in aged rats with preserved memory. PLoS ONE 2013, 8, e83674. [Google Scholar] [CrossRef] [Green Version]
- Arai, K.; Matsuki, N.; Ikegaya, Y.; Nishiyama, N. Deterioration of spatial learning performances in lipopolysaccharide-treated mice. Jpn. J. Pharmacol. 2001, 87, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Dallagi, Y.; Rahali, D.; Perrotte, M.; Dkhili, H.; Korsan, A.; El May, M.V.; El Fazaa, S.; Ramassamy, C.; El Golli, N. Date seeds alleviate behavioural and neuronal complications of metabolic syndrome in rats. Arch. Physiol. Biochem. 2020, 8, 1–15. [Google Scholar] [CrossRef]
- Treviño, S.; Aguilar-Alonso, P.; Flores Hernandez, J.A.; Brambila, E.; Guevara, J.; Flores, G.; Lopez-Lopez, G.; Muñoz-Arenas, G.; Morales-Medina, J.C.; Toxqui, V. A high calorie diet causes memory loss, metabolic syndrome and oxidative stress into hippocampus and temporal cortex of rats. Synapse 2015, 69, 421–433. [Google Scholar] [CrossRef]
- Muñoz-Arenas, G.; Pulido, G.; Treviño, S.; Vázquez-Roque, R.; Flores, G.; Moran, C.; Handal-Silva, A.; Guevara, J.; Venegas, B.; Díaz, A. Effects of metformin on recognition memory and hippocampal neuroplasticity in rats with metabolic syndrome. Synapse 2020, 74, e22153. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, A.R.; Tong, X.Y.; Shamaladevi, N.; Barcelona, S.; Gaidosh, G.; Agarwal, A.; Norenberg, M.D. Defective synthesis and release of astrocytic thrombospondin-1 mediates the neuronal TDP-43 proteinopathy, resulting in defects in neuronal integrity associated with chronic traumatic encephalopathy: In vitro studies. J. Neurochem. 2017, 140, 645–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.-K.; Carreras, I.; Aytan, N.; Jenkins-Sahlin, E.; Dedeoglu, A.; Jenkins, B.G. The effects of aging, housing and ibuprofen treatment on brain neurochemistry in a triple transgene Alzheimer’s disease mouse model using magnetic resonance spectroscopy and imaging. Brain Res. 2014, 1590, 85–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallérac, G.; Rouach, N. Astrocytes as new targets to improve cognitive functions. Prog. Neurobiol. 2016, 144, 48–67. [Google Scholar] [CrossRef]
- Elberry, D.A.; Amin, S.N.; Esmail, R.S.E.N.; Rashed, L.A.; Gamal, M.M. Effect of undifferentiated versus hepatogenic partially differentiated mesenchymal stem cells on hepatic and cognitive functions in liver cirrhosis. Excli J. 2016, 15, 652–670. [Google Scholar] [CrossRef]
- Zhao, Y.; Pu, D.; Sun, Y.; Chen, J.; Luo, C.; Wang, M.; Zhou, J.; Lv, A.; Zhu, S.; Liao, Z. High glucose-induced defective thrombospondin-1 release from astrocytes via TLR9 activation contributes to the synaptic protein loss. Exp. Cell Res. 2018, 363, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Stranahan, A.M.; Cutler, R.G.; Button, C.; Telljohann, R.; Mattson, M.P. Diet-induced elevations in serum cholesterol are associated with alterations in hippocampal lipid metabolism and increased oxidative stress. J. Neurochem. 2011, 118, 611–615. [Google Scholar] [CrossRef] [Green Version]
- Funari, V.A.; Crandall, J.E.; Tolan, D.R. Fructose metabolism in the cerebellum. Cerebellum 2007, 6, 130. [Google Scholar] [CrossRef]
- Rivera, D.S.; Lindsay, C.B.; Codocedo, J.F.; Carreño, L.E.; Cabrera, D.; Arrese, M.A.; Vio, C.P.; Bozinovic, F.; Inestrosa, N.C. Long-Term, Fructose-Induced Metabolic Syndrome-Like Condition Is Associated with Higher Metabolism, Reduced Synaptic Plasticity and Cognitive Impairment in Octodon degus. Mol. Neurobiol. 2018, 55, 9169–9187. [Google Scholar] [CrossRef] [PubMed]
- Nemes, A.D.; Ayasoufi, K.; Ying, Z.; Zhou, Q.-G.; Suh, H.; Najm, I.M. Growth Associated Protein 43 (GAP-43) as a Novel Target for the Diagnosis, Treatment and Prevention of Epileptogenesis. Sci. Rep. 2017, 7, 17702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holahan, M.R. A Shift from a Pivotal to Supporting Role for the Growth-Associated Protein (GAP-43) in the Coordination of Axonal Structural and Functional Plasticity. Front. Cell. Neurosci. 2017, 11, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Cheng, X.; Luo, X.; Yang, X.; Pang, Y.; Zhang, X.; Zhang, Y.; Liu, Y. Luteolin Ameliorates Cognitive Impairments by Suppressing the Expression of Inflammatory Cytokines and Enhancing Synapse-Associated Proteins GAP-43 and SYN Levels in Streptozotocin-Induced Diabetic Rats. Neurochem. Res. 2018, 43, 1905–1913. [Google Scholar] [CrossRef] [PubMed]
- Sandelius, Å.; Portelius, E.; Källén, Å.; Zetterberg, H.; Rot, U.; Olsson, B.; Toledo, J.B.; Shaw, L.M.; Lee, V.M.Y.; Irwin, D.J.; et al. Elevated CSF GAP-43 is Alzheimer’s disease specific and associated with tau and amyloid pathology. Alzheimers Dement. 2019, 15, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Gorup, D.; Bohaček, I.; Miličević, T.; Pochet, R.; Mitrečić, D.; Križ, J.; Gajović, S. Increased expression and colocalization of GAP43 and CASP3 after brain ischemic lesion in mouse. Neurosci. Lett. 2015, 15, 176–182. [Google Scholar] [CrossRef]
- Sandelius, Å.; Cullen, N.C.; Källén, Å.; Rosengren, L.; Jensen, C.; Kostanjevecki, V.; Vandijck, M.; Zetterberg, H.; Blennow, K. Transient increase in CSF GAP-43 concentration after ischemic stroke. BMC Neurol. 2018, 18, 202. [Google Scholar] [CrossRef] [Green Version]
- Page, K.A.; Chan, O.; Arora, J.; Belfort-DeAguiar, R.; Dzuira, J.; Roehmholdt, B.; Cline, G.W.; Naik, S.; Sinha, R.; Constable, R.T.; et al. Effects of Fructose vs Glucose on Regional Cerebral Blood Flow in Brain Regions Involved with Appetite and Reward Pathways. JAMA 2013, 309, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo-Ochoa, E.; Hernández-Ortega, K.; Ferrera, P.; Morimoto, S.; Arias, C. Short-Term High-Fat-and-Fructose Feeding Produces Insulin Signaling Alterations Accompanied by Neurite and Synaptic Reduction and Astroglial Activation in the Rat Hippocampus. J. Cereb. Blood Flow Metab. 2014, 34, 1001–1008. [Google Scholar] [CrossRef]
- Hung, C.-C.; Lin, C.-H.; Chang, H.; Wang, C.-Y.; Lin, S.-H.; Hsu, P.-C.; Sun, Y.-Y.; Lin, T.-N.; Shie, F.-S.; Kao, L.-S.; et al. Astrocytic GAP43 Induced by the TLR4/NF-κB/STAT3 Axis Attenuates Astrogliosis-Mediated Microglial Activation and Neurotoxicity. J. Neurosci. 2016, 36, 2027–2043. [Google Scholar] [CrossRef]
- Mastrocola, R.; Nigro, D.; Cento, A.S.; Chiazza, F.; Collino, M.; Aragno, M. High-fructose intake as risk factor for neurodegeneration: Key role for carboxy methyllysine accumulation in mice hippocampal neurons. Neurobiol. Dis. 2016, 89, 65–75. [Google Scholar] [CrossRef]
- Frye, C.A.; Koonce, C.J.; Walf, A.A. Progesterone, compared to medroxyprogesterone acetate, to C57BL/6, but not 5α-reductase mutant, mice enhances object recognition and placement memory and is associated with higher BDNF levels in the hippocampus and cortex. Neurosci. Lett. 2013, 551, 53–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meltzer-Brody, S.; Kanes, S.J. Allopregnanolone in postpartum depression: Role in pathophysiology and treatment. Neurobiol Stress. 2020, 12, 100212. [Google Scholar] [CrossRef] [PubMed]
- Bonansco, C.; Fuenzalida, M. Plasticity of Hippocampal Excitatory-Inhibitory Balance: Missing the Synaptic Control in the Epileptic Brain. Neural. Plast. 2016, e8607038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Son, Y.; Kim, J.; Lee, S.; Kang, S.; Park, K.; Kim, S.H.; Kim, J.C.; Kim, J.; Takayama, C.; et al. Developmental and degenerative modulation of GABAergic transmission in the mouse hippocampus. Int. J. Dev. Neurosci. 2015, 47, 320–332. [Google Scholar] [CrossRef]
- Whissell, P.D.; Rosenzweig, S.; Lecker, I.; Wang, D.-S.; Wojtowicz, J.M.; Orser, B.A. γ-aminobutyric acid type A receptors that contain the δ subunit promote memory and neurogenesis in the dentate gyrus. Ann. Neurol. 2013, 74, 611–621. [Google Scholar] [CrossRef]
- Napoli, E.; Schneider, A.; Wang, J.Y.; Trivedi, A.; Carrillo, N.R.; Tassone, F.; Rogawski, M.; Hagerman, R.J.; Giulivi, C. Allopregnanolone Treatment Improves Plasma Metabolomic Profile Associated with GABA Metabolism in Fragile X-Associated Tremor/Ataxia Syndrome: A Pilot Study. Mol. Neurobiol. 2019, 56, 3702–3713. [Google Scholar] [CrossRef]
- Gall, W.E.; Beebe, K.; Lawton, K.A.; Adam, K.-P.; Mitchell, M.W.; Nakhle, P.J.; Ryals, J.A.; Milburn, M.V.; Nannipieri, M.; Camastra, S.; et al. Group, for the the RS α-Hydroxybutyrate Is an Early Biomarker of Insulin Resistance and Glucose Intolerance in a Nondiabetic Population. PLoS ONE 2010, 5, e10883. [Google Scholar] [CrossRef] [Green Version]
- Frye, C.A.; Lembo, V.F.; Walf, A.A. Progesterone’s Effects on Cognitive Performance of Male Mice Are Independent of Progestin Receptors but Relate to Increases in GABAA Activity in the Hippocampus and Cortex. Front. Endocrinol. 2021, 11, 552805. [Google Scholar] [CrossRef] [PubMed]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Pistell, P.J.; Morrison, C.D.; Gupta, S.; Knight, A.G.; Keller, J.N.; Ingram, D.K.; Bruce-Keller, A.J. Cognitive impairment following high fat diet consumption is associated with brain inflammation. J. Neuroimmunol. 2010, 219, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Ye, X.; Bian, W.; Chen, Z.; Du, J.; Li, M.; Zhou, P.; Cui, H.; Ding, Y.Q.; Qi, S.; et al. Allopregnanolone Modulates GABAAR-Dependent CaMKIIδ3 and BDNF to Protect SH-SY5Y Cells Against 6-OHDA-Induced Damage. Front Cell Neurosci. 2020, 13, 569. [Google Scholar] [CrossRef] [Green Version]
- Almeida, F.B.; Nin, M.S.; Barros, H.M.T. The role of Allopregnanolone in depressive-like behaviors: Focus on neurotrophic proteins. Neurobiol. Stress. 2020, 12, 100218. [Google Scholar] [CrossRef]
- Almeida, F.B.; Barros, H.M.T.; Pinna, G. Neurosteroids and Neurotrophic Factors: What Is Their Promise as Biomarkers for Major Depression and PTSD? Int. J. Mol. Sci. 2021, 22, 1758. [Google Scholar] [CrossRef]
- Xu, A.-H.; Yang, Y.; Sun, Y.-X.; Zhang, C.-D. Exogenous brain-derived neurotrophic factor attenuates cognitive impairment induced by okadaic acid in a rat model of Alzheimer’s disease. Neural Regen. Res. 2018, 13, 2173–2181. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhu, Y.; He, T.; Li, W.; Li, Q.; Miao, Y. Brain-derived neurotrophic factor inhibits hyperglycemia-induced apoptosis and downregulation of synaptic plasticity-related proteins in hippocampal neurons via the PI3K/Akt pathway. Int. J. Mol. Med. 2019, 43, 294–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampaio, T.B.; Savall, A.S.; Gutierrez, M.E.Z.; Pinton, S. Neurotrophic factors in Alzheimer’s and Parkinson’s diseases: Implications for pathogenesis and therapy. Neural Regen. Res. 2017, 12, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Johnston, P.B.; Ball, B.G.; Brinton, R.D. The Neurosteroid Allopregnanolone Promotes Proliferation of Rodent and Human Neural Progenitor Cells and Regulates Cell-Cycle Gene and Protein Expression. J. Neurosci. 2005, 25, 4706–4718. [Google Scholar] [CrossRef]
- Singh, C.; Liu, L.; Wang, J.M.; Irwin, R.W.; Yao, J.; Chen, S.; Henry, S.; Thompson, R.F.; Brinton, R.D. Allopregnanolone restores hippocampal-dependent learning and memory and neural progenitor survival in aging 3xTgAD and nonTg mice. Neurobiol. Aging 2012, 33, 1493–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haage, D.; Johansson, S. Neurosteroid modulation of synaptic and GABAevoked currents in neurons from the rat medial preoptic nucleus. J. Neurophysiol. 1999, 82, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Dang, H.N.; Yong, J.; Chui, W.S.; Dizon MP, G.; Yaw CK, Y.; Kaufman, D.L. Oral treatment with gamma-aminobutyric acid improves glucose tolerance and insulin sensitivity by inhibiting inflammation in high fat diet-fed mice. PLoS ONE 2011, 6, e25338. [Google Scholar] [CrossRef] [Green Version]
- Xu, E.; Kumar, M.; Zhang, Y.; Ju, W.; Obata, T.; Zhang, N.; Liu, S.; Wendt, A.; Deng, S.; Ebina, Y.; et al. Intra-islet insulin suppresses glucagon release via GABA-GABAA receptor system. Cell Metab. 2006, 3, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Braun, M.; Ramracheya, R.; Bengtsson, M.; Clark, A.; Walker, J.N.; Johnson, P.R.; Rorsman, P. Gamma-Aminobutyric Acid (GABA) Is an Autocrine Excitatory Transmitter in Human Pancreatic Beta-Cells. Diabetes 2010, 59, 1694–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmade, S.J.; Gale, S.E.; Frolov, A.; Mohri, I.; Suzuki, K.; Mellon, S.H.; Walkley, S.U.; Covey, D.F.; Schaffer, J.E.; Ory, D.S. Pregnane X receptor (PXR) activation: A mechanism for neuroprotection in a mouse model of Niemann-Pick C disease. Proc. Natl. Acad. Sci. USA 2006, 103, 13807–13812. [Google Scholar] [CrossRef] [Green Version]
- Roth, A.; Looser, R.; Kaufmann, M.; Blättler, S.M.; Rencurel, F.; Huang, W.; Moore, D.D.; Meyer, U.A. Regulatory cross-talk between drug metabolism and lipid homeostasis: Constitutive androstane receptor and pregnane X receptor increase Insig-1 expression. Mol. Pharmacol. 2008, 73, 1282–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monnerie, H.; Romer, M.; Jensen, B.K.; Millar, J.S.; Jordan-Sciutto, K.L.; Kim, S.F.; Grinspan, J.B. Reduced sterol regulatory element-binding protein (SREBP) processing through site-1 protease (S1P) inhibition alters oligodendrocyte differentiation in vitro. J. Neurochem. 2017, 140, 53–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevenson, E.; Johns, E.; Marques, F.; Jackson, K.; Davern, P.; Evansc, R.; Head, G. Positive allosteric modulation of GABA-A receptors attenuates high blood pressure in Schlager hypertensive mice. J. Hypertens. 2017, 35, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Milivojevic, V.; Fox, H.C.; Sofuoglu, M.; Covault, J.; Sinha, R. Effects of progesterone stimulated Allopregnanolone on craving and stress response in cocaine dependent men and women. Psychoneuroendocrinology 2016, 65, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Shimada, M.; Hasegawa, T.; Nishimura, C.; Kan, H.; Kanno, T.; Nakamura, T.; Matsubayashi, T. Anti-Hypertensive Effect of g-Aminobutyric Acid (GABA)-Rich Chlorella on High-Normal Blood Pressure and Borderline Hypertension in Placebo-Controlled Double Blind Study. Clin. Exp. Hypertens. 2009, 31, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Schillaci, G.; Pirro, M.; Vaudo, G.; Gemelli, F.; Marchesi, S.; Porcellati, C.; Mannarino, E. Prognostic value of the metabolic syndrome in essential hypertension. J. Am. Coll. Cardiol. 2004, 43, 1817–1822. [Google Scholar] [CrossRef] [Green Version]
- Grassi, G.; Seravalle, G.; Dell’Oro, R.; Quarti Trevano, F.; Bombelli, M.; Scopelliti, F.; Facchini, A.; Mancia, G. Comparative effects of candesartan and hydrochlorothiazide on blood pressure, insulin sensitivity, and sympathetic drive in obese hypertensive individuals: Results of the CROSS study. J. Hypertens. 2003, 21, 1761–1769. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Dell’Oro, R.; Quarti Trevano, F.; Scopelliti, F.; Seravalle, G.; Paleari, F.; Gamba, P.L.; Mancia, G. Neuroadrenergic and reflex abnormalities in patients with metabolic syndrome. Diabetologia 2005, 48, 1359–1365. [Google Scholar] [CrossRef] [Green Version]
- Reaven, G.M.; Lithell, H.; Landsberg, L. Hypertension and associated metabolic abnormalities—the role of insulin resistance and the sympathoadrenal system. N. Engl. J. Med. 1996, 334, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, P.; Tappy, L.; Randin, D.; Schneiter, P.; Jequier, E.; Nicod, P.; Scherrer, U. Differential effects of hyperinsulinemia and carbohydrate metabolism on sympathetic nerve activity and muscle blood flow in humans. J. Clin. Investig. 1993, 92, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudbjornsdottir, S.; Friberg, P.; Elam, M.; Attvall, S.; Lonnroth, P.; Wallin, B.G. The effects of metformin and insulin on sympathetic nerve activity, norepinehrine spillover and blood pressure in obese, insulin resistant, normoglycemic, hypertensive men. Blood Press 1994, 3, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Maguire, J.; Stell, B.; Rafizadeh, M.; Mody, I. Ovarian cycle-linked changes in GABAA receptors mediating tonic inhibition alter seizure susceptibility and anxiety. Nat. Neurosci. 2005, 8, 797–804. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer Sequence | Accession Number | |
|---|---|---|
| Synaptophysin | Forward primer: 5′-TCCAATCAGATGTAGTCTGGTCAG-3′ Reverse primer: 5′-AGGCCTTCTCCTGAGCTCTT-3′ | NM_009305.2 |
| GAP-43 | Forward primer: 5′-TTTCCTCTCCTGTCCTGCTC-3′ Reverse primer: 5′-TGGACTTGGGATCTTTCCTG-′3 | NM_008083.2 |
| glyceraldehyde-3-phosphate dehydrogenase (Gapdh) | Forward primer: 5′-GGTCGGTGTGAACGGATTTGG-3′ Reverse primer: 5′-ATGTAGGCCATGAGGTCCACC-3′ | NM_001289726.1 |
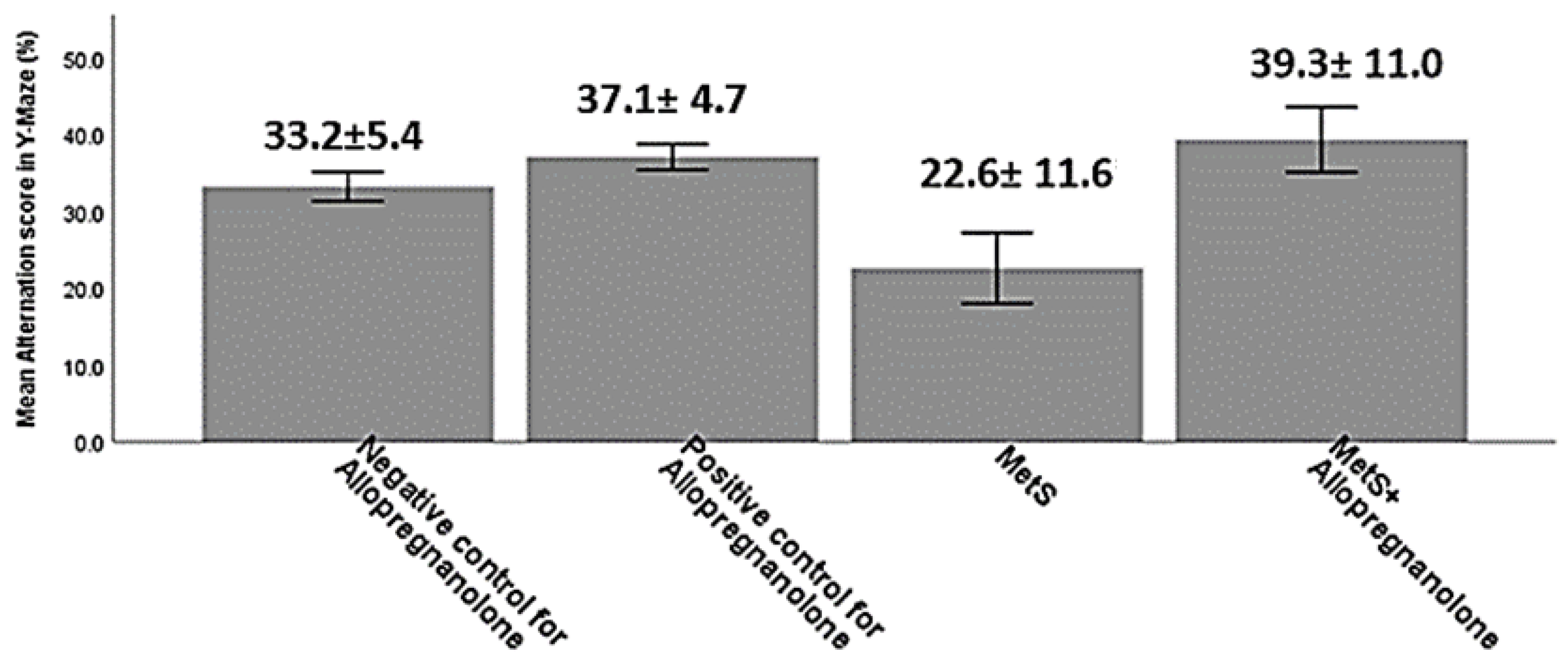
| Measured Parameters | Negative Control for Allopregnanolone | Positive Control for Allopregnanolone | MetS | MetS+ Allopregnanolone | F-Values | p-Value |
|---|---|---|---|---|---|---|
| Serum Glucose (mg/dL) | 97.3 ± 12.9 a | 107.8 ± 8.2 a | 250.8 ± 53.8 b | 189.0 ± 36.6 c | 37.6 | <0.001 |
| Serum Insulin (µIU/L) | 9.2 ± 0.9 a | 14.4 ± 1.1 b | 21.3 ± 3.9 c | 16.7 ± 2.0 b | 40.1 | <0.001 |
| HOMA-IR | 2.20 ± 0.34 a | 3.72 ± 0.44 a | 12.97 ± 2.8 b | 7.85 ± 2.03 c | 33.5 | <0.001 |
| Serum Triglycerides (mg/dL) | 75.5 ± 14.9 a | 91.2 ± 22.9 a | 143.4 ± 17.5 b | 126.7 ± 5.6 b | 28.7 | <0.001 |
| Serum Cholesterol (mg/dL) | 130.9 ± 3.9 a | 146.2 ± 25.4 a | 204.6 ± 16.3 b | 153.9 ± 45.1 a | 11.1 | <0.001 |
| Serum HDL (mg/dL) | 58.3 ± 3.4 a | 56.9 ± 9.1 a | 28.6 ± 3.2 b | 43.1 ± 5.1 c | 47.1 | <0.001 |
| Hippocampal Synaptophysin (relative expression) | 1.8 ± 0.2 a | 1.3 ± 0.1 b | 0.1 ± 0.03 c | 0.9 ± 0.1 d | 155.1 | <0.001 |
| Hippocampal GAP43 (relative expression) | 1.1 ± 0.03 a | 1.2 ± 0.3 a | 4.5 ± 0.6 b | 2.7 ± 0.5 c | 112.9 | <0.001 |
| Measured Parameters | Negative Control for Allopregnanolone | Positive Control for Allopregnanolone | MetS | MetS+Allopregnanolone | F-Value | p-Value |
|---|---|---|---|---|---|---|
| Systolic BP (mmHg) | 116.8 ± 4.1 a | 104.9 ± 4.3 b | 151.5 ± 11.0 c | 118.5 ± 5.1 a | 70.3 | <0.001 |
| Diastolic BP (mmHg) | 84.5 ± 6.6 a | 86.4 ± 3.6 a | 113.6 ± 6.3 b | 90.9 ± 2.3 a | 56.6 | <0.001 |
| MAP (mmHg) | 96.6 ± 4.2 a | 91.5 ± 3.7 a | 128.3 ± 8.7 b | 100.4 ± 3.4 a | 72.6 | <0.001 |
| Biochemical Parameters | Alternation Score in Y-Maze (%) | |
|---|---|---|
| Correlation Coefficient (r) | p Value | |
| Serum Glucose (mg/dL) | −0.086 | 0.640 |
| Serum Insulin ((µIU/L) | −0.334 | 0.062 |
| Serum Triglycerides (mg/dL) | −0.301 | 0.094 |
| Serum Cholesterol (mg/dL) | −0.204 | 0.262 |
| Serum HDL (mg/dL) | 0.317 | 0.077 |
| Hippocampal Synaptophysin (relative expression) | 0.366 | 0.039 |
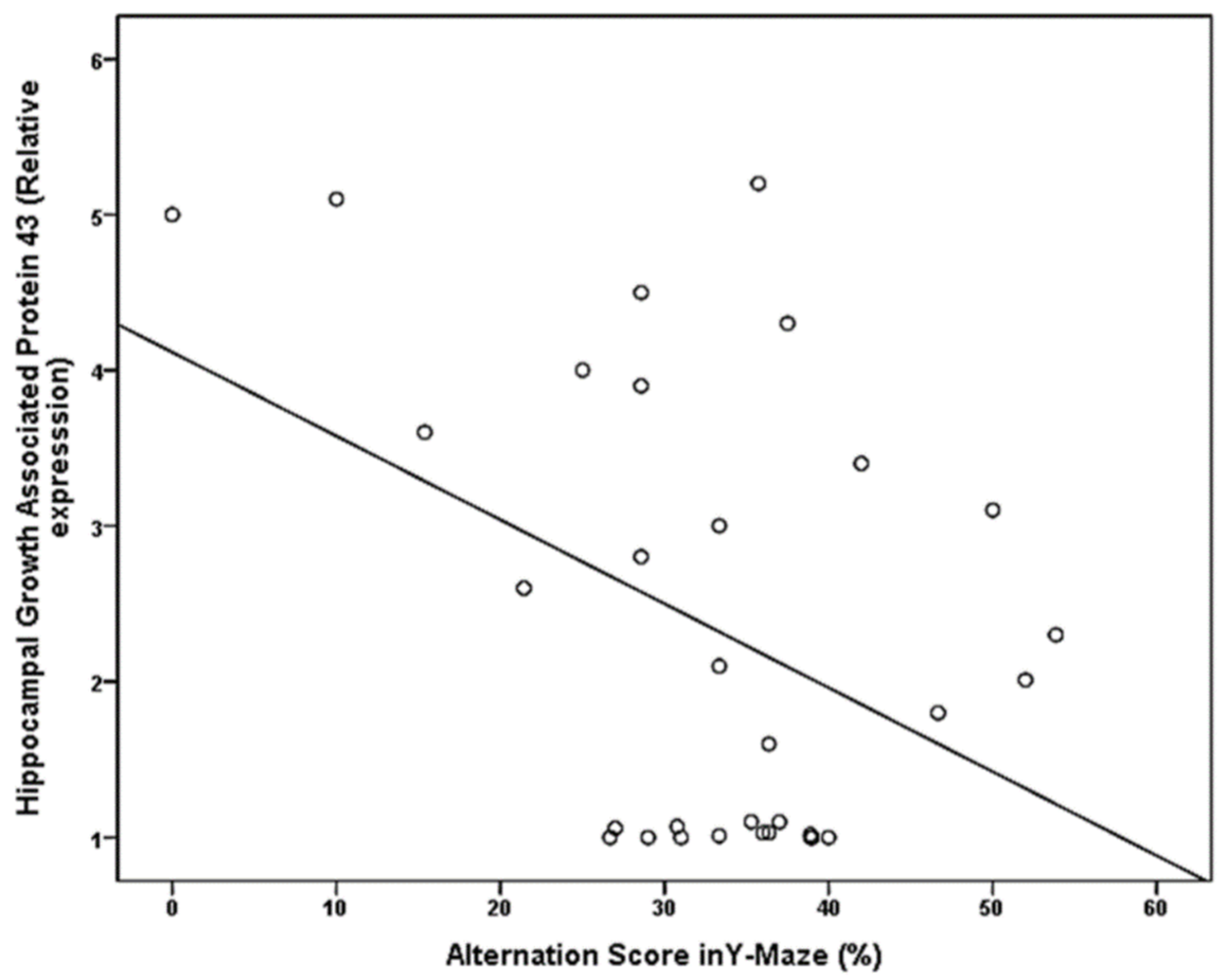
| Hippocampal Growth Associated Protein 43 (GAP43) (relative expression) | −0.414 | 0.019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amin, S.N.; Abdalgeleel, S.A.; Algahtany, M.A.; Shaltout, S.A.; El Gazzar, W.B.; Elberry, D.A. Effect of Allopregnanolone on Spatial Memory and Synaptic Proteins in Animal Model of Metabolic Syndrome. Brain Sci. 2021, 11, 644. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11050644
Amin SN, Abdalgeleel SA, Algahtany MA, Shaltout SA, El Gazzar WB, Elberry DA. Effect of Allopregnanolone on Spatial Memory and Synaptic Proteins in Animal Model of Metabolic Syndrome. Brain Sciences. 2021; 11(5):644. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11050644
Chicago/Turabian StyleAmin, Shaimaa Nasr, Shaimaa Abdalaleem Abdalgeleel, Mubarak Ali Algahtany, Sherif Ahmed Shaltout, Walaa Bayoumie El Gazzar, and Dalia Azmy Elberry. 2021. "Effect of Allopregnanolone on Spatial Memory and Synaptic Proteins in Animal Model of Metabolic Syndrome" Brain Sciences 11, no. 5: 644. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11050644