
Effects of Transcranial Ultrasound Stimulation on Trigeminal Blink Reflex Excitability

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
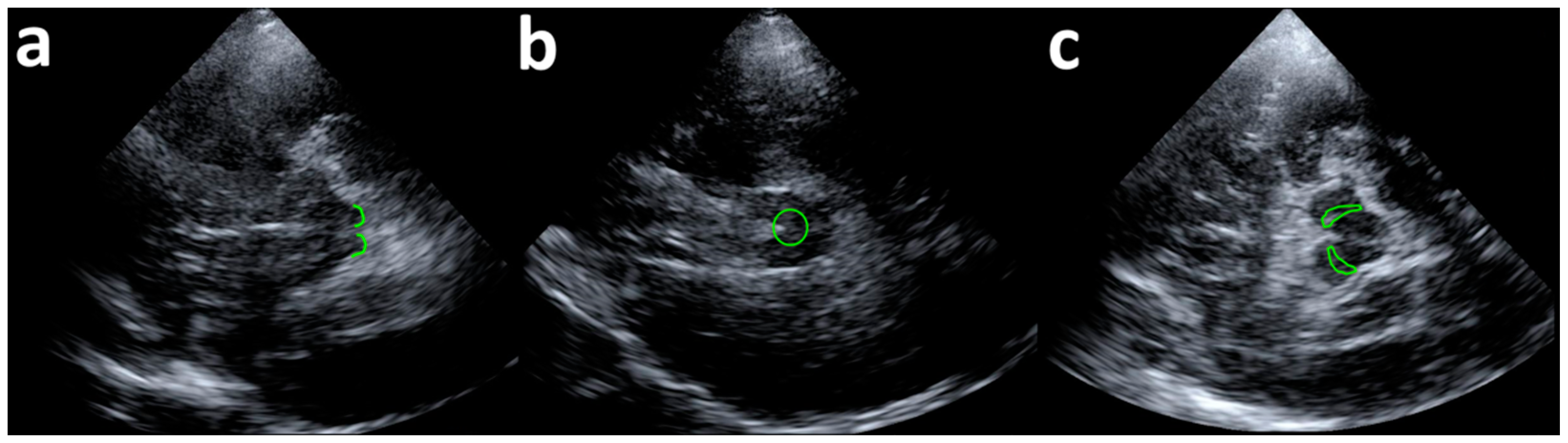
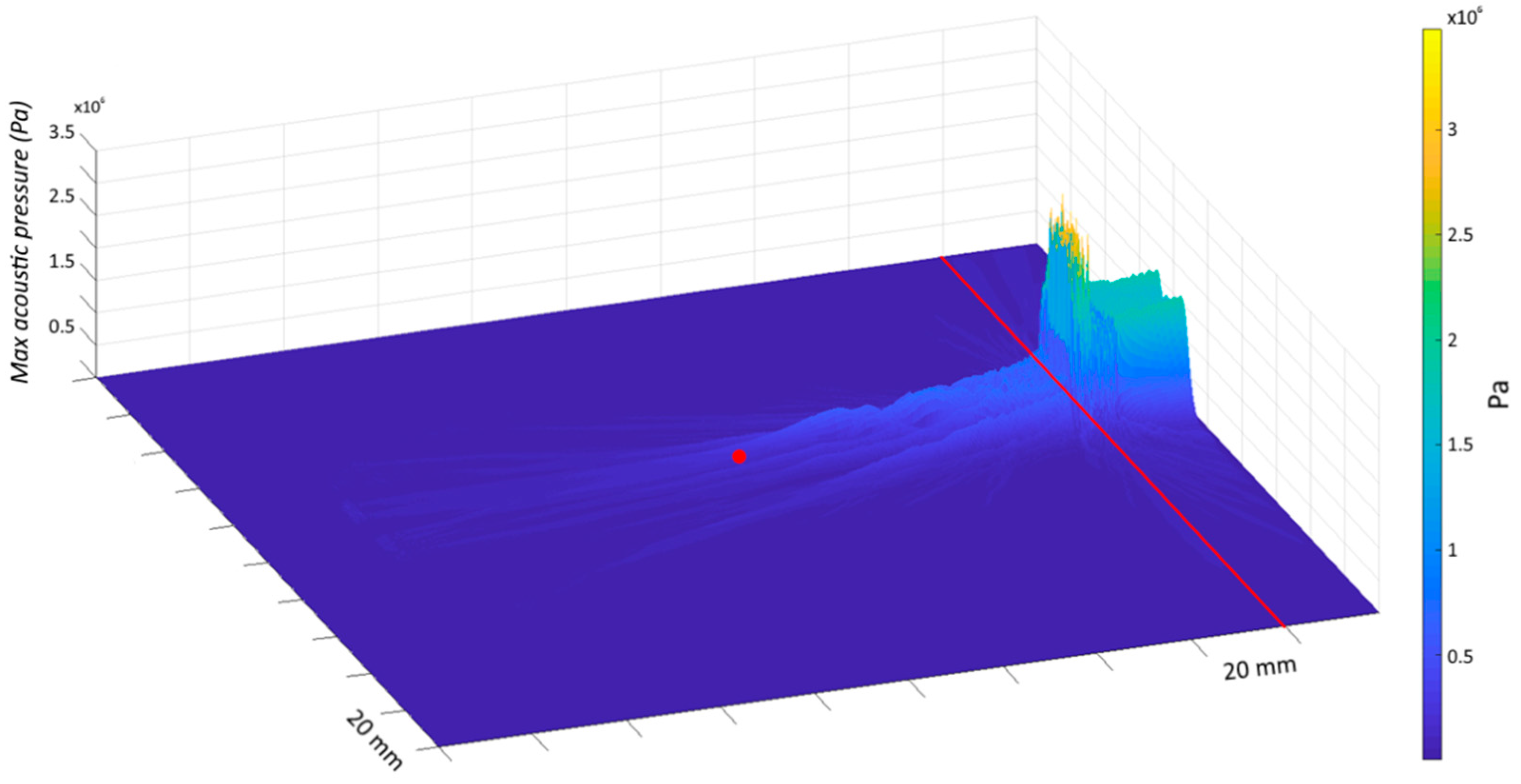
2.2. Transcranial Ultrasound Stimulation
2.3. Blink Reflex and R2 Recovery Cycle
2.4. Experimental Procedure
2.5. Statistical Analysis
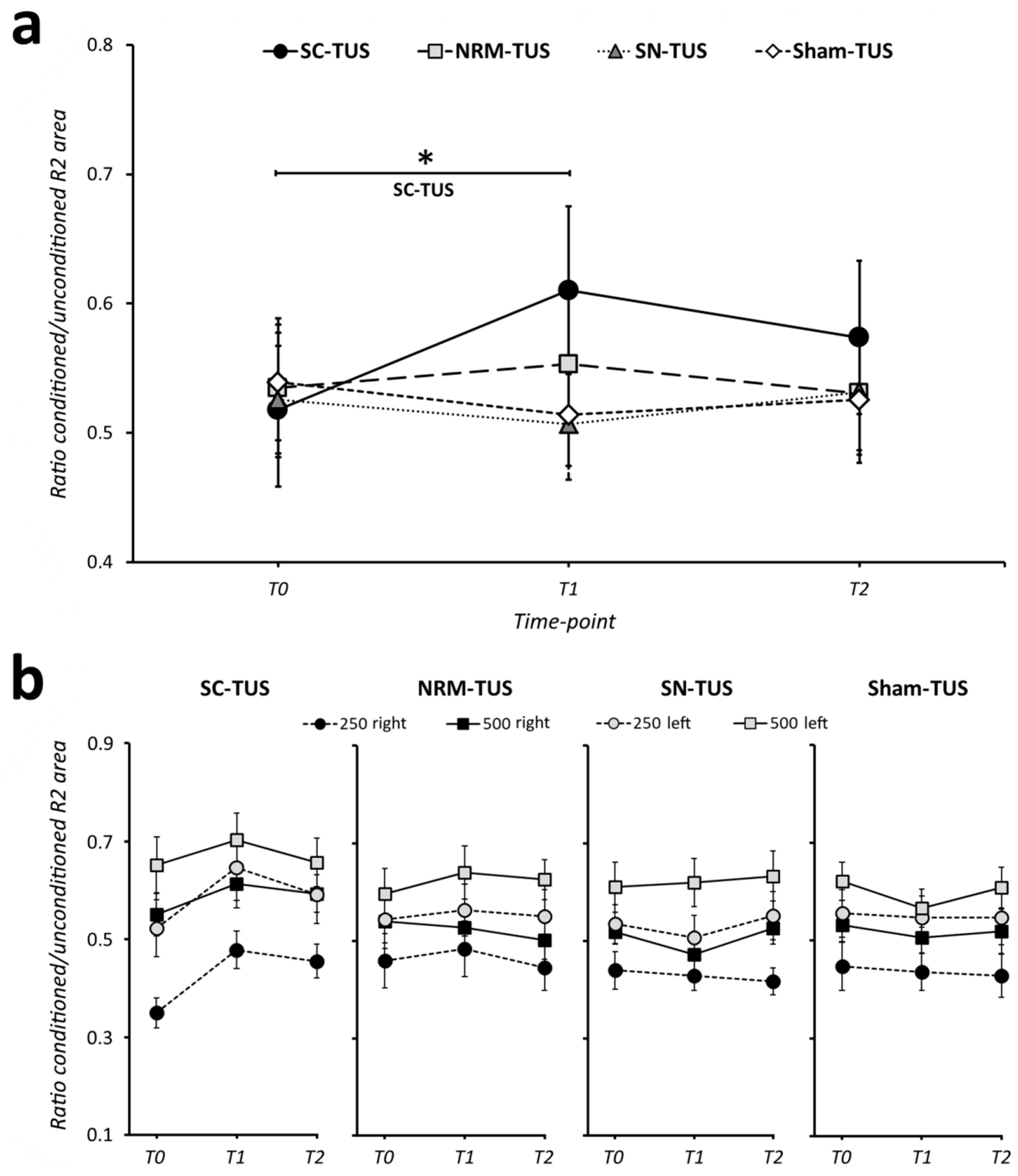
3. Results
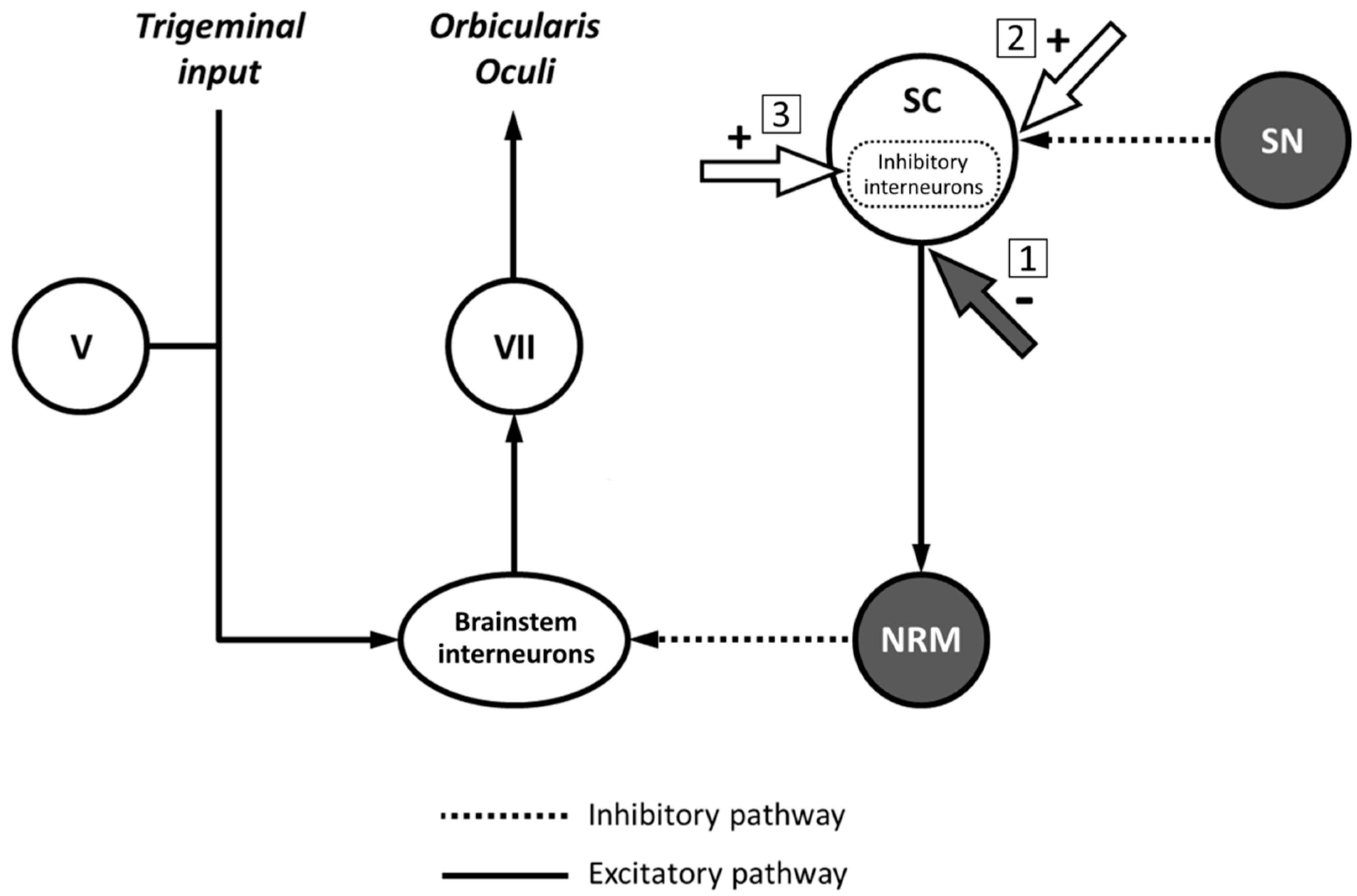
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tyler, W.J.; Tufail, Y.; Finsterwald, M.; Tauchmann, M.L.; Olson, E.J.; Majestic, C. Remote Excitation of Neuronal Circuits Using Low-Intensity, Low-Frequency Ultrasound. PLoS ONE 2008, 3, e3511. [Google Scholar] [CrossRef] [PubMed]
- Tufail, Y.; Matyushov, A.; Baldwin, N.; Tauchmann, M.L.; Georges, J.; Yoshihiro, A.; Tillery, S.I.H.; Tyler, W.J. Transcranial Pulsed Ultrasound Stimulates Intact Brain Circuits. Neuron 2010, 66, 681–694. [Google Scholar] [CrossRef] [Green Version]
- Blackmore, J.; Shrivastava, S.; Sallet, J.; Butler, C.R.; Cleveland, R.O. Ultrasound Neuromodulation: A Review of Results, Mechanisms and Safety. Ultrasound Med. Biol. 2019, 45, 1509–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubanek, J.; Shukla, P.; Das, A.; Baccus, S.A.; Goodman, M.B. Ultrasound Elicits Behavioral Responses through Mechanical Effects on Neurons and Ion Channels in a Simple Nervous System. J. Neurosci. 2018, 38, 3081–3091. [Google Scholar] [CrossRef] [Green Version]
- Krasovitski, B.; Frenkel, V.; Shoham, S.; Kimmel, E. Intramembrane Cavitation as a Unifying Mechanism for Ultrasound-Induced Bioeffects. Proc. Natl. Acad. Sci. USA 2011, 108, 3258–3263. [Google Scholar] [CrossRef] [Green Version]
- Di Biase, L.; Falato, E.; Di Lazzaro, V. Transcranial Focused Ultrasound (TFUS) and Transcranial Unfocused Ultrasound (TUS) Neuromodulation: From Theoretical Principles to Stimulation Practices. Front. Neurol. 2019, 10, 549. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhang, J.; Yu, J.; Smith, C.; Feng, W. Brain Modulatory Effects by Low-Intensity Transcranial Ultrasound Stimulation (TUS): A Systematic Review on Both Animal and Human Studies. Front. Neurosci. 2019, 13, 696. [Google Scholar] [CrossRef]
- Legon, W.; Bansal, P.; Tyshynsky, R.; Ai, L.; Mueller, J.K. Transcranial Focused Ultrasound Neuromodulation of the Human Primary Motor Cortex. Sci. Rep. 2018, 8, 10007. [Google Scholar] [CrossRef] [Green Version]
- Fomenko, A.; Chen, K.-H.S.; Nankoo, J.-F.; Saravanamuttu, J.; Wang, Y.; El-Baba, M.; Xia, X.; Seerala, S.S.; Hynynen, K.; Lozano, A.M.; et al. Systematic Examination of Low-Intensity Ultrasound Parameters on Human Motor Cortex Excitability and Behavior. Elife 2020, 9. [Google Scholar] [CrossRef]
- Gibson, B.C.; Sanguinetti, J.L.; Badran, B.W.; Yu, A.B.; Klein, E.P.; Abbott, C.C.; Hansberger, J.T.; Clark, V.P. Increased Excitability Induced in the Primary Motor Cortex by Transcranial Ultrasound Stimulation. Front. Neurol. 2018, 9, 1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legon, W.; Sato, T.F.; Opitz, A.; Mueller, J.; Barbour, A.; Williams, A.; Tyler, W.J. Transcranial Focused Ultrasound Modulates the Activity of Primary Somatosensory Cortex in Humans. Nat. Neurosci. 2014, 17, 322–329. [Google Scholar] [CrossRef]
- Lee, W.; Kim, H.; Jung, Y.; Song, I.-U.; Chung, Y.A.; Yoo, S.-S. Image-Guided Transcranial Focused Ultrasound Stimulates Human Primary Somatosensory Cortex. Sci. Rep. 2015, 5, 8743. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Chung, Y.A.; Jung, Y.; Song, I.-U.; Yoo, S.-S. Simultaneous Acoustic Stimulation of Human Primary and Secondary Somatosensory Cortices Using Transcranial Focused Ultrasound. BMC Neurosci. 2016, 17, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legon, W.; Ai, L.; Bansal, P.; Mueller, J.K. Neuromodulation with Single-Element Transcranial Focused Ultrasound in Human Thalamus. Hum. Brain Mapp. 2018, 39, 1995–2006. [Google Scholar] [CrossRef] [Green Version]
- Berardelli, A.; Cruccu, G.; Manfredi, M.; Rothwell, J.C.; Day, B.L.; Marsden, C.D. The Corneal Reflex and the R2 Component of the Blink Reflex. Neurology 1985, 35, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Cruccu, G.; Iannetti, G.D.; Marx, J.J.; Thoemke, F.; Truini, A.; Fitzek, S.; Galeotti, F.; Urban, P.P.; Romaniello, A.; Stoeter, P.; et al. Brainstem Reflex Circuits Revisited. Brain 2005, 128, 386–394. [Google Scholar] [CrossRef]
- Valls-Solé, J. Neurophysiological Assessment of Trigeminal Nerve Reflexes in Disorders of Central and Peripheral Nervous System. Clin. Neurophysiol. 2005, 116, 2255–2265. [Google Scholar] [CrossRef] [PubMed]
- Cruccu, G.; Deuschl, G. The Clinical Use of Brainstem Reflexes and Hand-Muscle Reflexes. Clin. Neurophysiol. 2000, 111, 371–387. [Google Scholar] [CrossRef]
- Boelhouwer, A.J. Blink Reflexes and Preparation. Biol. Psychol. 1982, 14, 277–285. [Google Scholar] [CrossRef]
- Valls-Solé, J.; Valldeoriola, F.; Tolosa, E.; Marti, M.J. Distinctive Abnormalities of Facial Reflexes in Patients with Progressive Supranuclear Palsy. Brain 1997, 120 Pt 10, 1877–1883. [Google Scholar] [CrossRef] [Green Version]
- Basso, M.A.; Evinger, C. An Explanation for Reflex Blink Hyperexcitability in Parkinson’s Disease. II. Nucleus Raphe Magnus. J. Neurosci. 1996, 16, 7318–7330. [Google Scholar] [CrossRef] [Green Version]
- Basso, M.A.; Powers, A.S.; Evinger, C. An Explanation for Reflex Blink Hyperexcitability in Parkinson’s Disease. I. Superior Colliculus. J. Neurosci. 1996, 16, 7308–7317. [Google Scholar] [CrossRef]
- Valls-Sole, J. Assessment of Excitability in Brainstem Circuits Mediating the Blink Reflex and the Startle Reaction. Clin. Neurophysiol. 2012, 123, 13–20. [Google Scholar] [CrossRef]
- Bologna, M.; Fabbrini, G.; Marsili, L.; Defazio, G.; Thompson, P.D.; Berardelli, A. Facial Bradykinesia. J. Neurol. Neurosurg. Psychiatry 2013, 84, 681–685. [Google Scholar] [CrossRef]
- Bologna, M.; Berardelli, A. Brainstem Avenues in Parkinson’s Disease Research. Clin. Neurophysiol. 2019, 130, 554–555. [Google Scholar] [CrossRef]
- Vo, L.; Drummond, P.D. Coexistence of Ipsilateral Pain-Inhibitory and Facilitatory Processes after High-Frequency Electrical Stimulation. Eur. J. Pain 2014, 18, 376–385. [Google Scholar] [CrossRef] [Green Version]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Kern, R.; Perren, F.; Kreisel, S.; Szabo, K.; Hennerici, M.; Meairs, S. Multiplanar Transcranial Ultrasound Imaging: Standards, Landmarks and Correlation with Magnetic Resonance Imaging. Ultrasound Med. Biol. 2005, 31, 311–315. [Google Scholar] [CrossRef]
- Becker, G.; Berg, D. Neuroimaging in Basal Ganglia Disorders: Perspectives for Transcranial Ultrasound. Mov. Disord. 2001, 16, 23–32. [Google Scholar] [CrossRef]
- Treeby, B.E.; Cox, B.T. K-Wave: MATLAB Toolbox for the Simulation and Reconstruction of Photoacoustic Wave Fields. J. Biomed. Opt. 2010, 15, 021314. [Google Scholar] [CrossRef]
- Strudwick Caviness, V.; Theodore Lange, N.; Makris, N.; Reed Herbert, M.; Nelson Kennedy, D. MRI-Based Brain Volumetrics: Emergence of a Developmental Brain Science. Brain Dev. 1999, 21, 289–295. [Google Scholar] [CrossRef]
- Fong, P.-Y.; Edwards, M.J.; Lu, C.-S.; Chen, R.-S.; Rothwell, J.C.; Bhatia, K.P.; Huang, Y.-Z. Abnormal Blink Reflex Recovery Cycle in Manifesting and Nonmanifesting Carriers of the DYT1 Gene Mutation. Neuroreport 2016, 27, 1046–1049. [Google Scholar] [CrossRef]
- Conte, A.; Fabbrini, G.; Belvisi, D.; Marsili, L.; Di Stasio, F.; Berardelli, A. Electrical Activation of the Orbicularis Oculi Muscle Does Not Increase the Effectiveness of Botulinum Toxin Type A in Patients with Blepharospasm. Eur. J. Neurol. 2010, 17, 449–455. [Google Scholar] [CrossRef]
- Paparella, G.; Di Stefano, G.; Fasolino, A.; Di Pietro, G.; Colella, D.; Truini, A.; Cruccu, G.; Berardelli, A.; Bologna, M. Painful Stimulation Increases Spontaneous Blink Rate in Healthy Subjects. Sci. Rep. 2020, 10, 20014. [Google Scholar] [CrossRef]
- Bologna, M.; Agostino, R.; Gregori, B.; Belvisi, D.; Manfredi, M.; Berardelli, A. Metaplasticity of the Human Trigeminal Blink Reflex. Eur. J. Neurosci. 2010, 32, 1707–1714. [Google Scholar] [CrossRef]
- Kimura, J.; Harada, O. Excitability of the Orbicularis Oculi Reflex in All Night Sleep: Its Suppression in Non-Rapid Eye Movement and Recovery in Rapid Eye Movement Sleep. Electroencephalogr. Clin. Neurophysiol. 1972, 33, 369–377. [Google Scholar] [CrossRef]
- Boelhouwer, A.J.; Brunia, C.H. Blink Reflexes and the State of Arousal. J. Neurol. Neurosurg. Psychiatry 1977, 40, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Goossens, H.H.; Van Opstal, A.J. Blink-Perturbed Saccades in Monkey. II. Superior Colliculus Activity. J. Neurophysiol. 2000, 83, 3430–3452. [Google Scholar] [CrossRef] [Green Version]
- Gnadt, J.W.; Lu, S.M.; Breznen, B.; Basso, M.A.; Henriquez, V.M.; Evinger, C. Influence of the Superior Colliculus on the Primate Blink Reflex. Exp. Brain Res. 1997, 116, 389–398. [Google Scholar] [CrossRef]
- Tardif, E.; Delacuisine, B.; Probst, A.; Clarke, S. Intrinsic Connectivity of Human Superior Colliculus. Exp. Brain Res. 2005, 166, 316–324. [Google Scholar] [CrossRef] [Green Version]
- May, P.J. The Mammalian Superior Colliculus: Laminar Structure and Connections. Prog. Brain Res. 2006, 151, 321–378. [Google Scholar] [CrossRef]
- Khraiche, M.L.; Phillips, W.B.; Jackson, N.; Muthuswamy, J. Ultrasound Induced Increase in Excitability of Single Neurons. In Proceedings of the Annual International Conference IEEE Engineering in Medicine and Biology Society, Vancouver, BC, Canada, 21–24 August 2008; pp. 4246–4249. [Google Scholar]
- Kim, H.-B.; Swanberg, K.M.; Han, H.-S.; Kim, J.-C.; Kim, J.-W.; Lee, S.; Lee, C.J.; Maeng, S.; Kim, T.-S.; Park, J.-H. Prolonged Stimulation with Low-Intensity Ultrasound Induces Delayed Increases in Spontaneous Hippocampal Culture Spiking Activity. J. Neurosci. Res. 2017, 95, 885–896. [Google Scholar] [CrossRef]
- Lin, W.-T.; Chen, R.-C.; Lu, W.-W.; Liu, S.-H.; Yang, F.-Y. Protective Effects of Low-Intensity Pulsed Ultrasound on Aluminum-Induced Cerebral Damage in Alzheimer’s Disease Rat Model. Sci. Rep. 2015, 5, 9671. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.-Y.; Lu, W.-W.; Lin, W.-T.; Chang, C.-W.; Huang, S.-L. Enhancement of Neurotrophic Factors in Astrocyte for Neuroprotective Effects in Brain Disorders Using Low-Intensity Pulsed Ultrasound Stimulation. Brain Stimul. 2015, 8, 465–473. [Google Scholar] [CrossRef]
- Behan, M.; Kime, N.M. Intrinsic Circuitry in the Deep Layers of the Cat Superior Colliculus. Vis. Neurosci. 1996, 13, 1031–1042. [Google Scholar] [CrossRef]
- Kaneda, K.; Isa, K.; Yanagawa, Y.; Isa, T. Nigral Inhibition of GABAergic Neurons in Mouse Superior Colliculus. J. Neurosci. 2008, 28, 11071–11078. [Google Scholar] [CrossRef] [Green Version]
- Phongphanphanee, P.; Mizuno, F.; Lee, P.H.; Yanagawa, Y.; Isa, T.; Hall, W.C. A Circuit Model for Saccadic Suppression in the Superior Colliculus. J. Neurosci. 2011, 31, 1949–1954. [Google Scholar] [CrossRef] [Green Version]
- Villalobos, C.A.; Wu, Q.; Lee, P.H.; May, P.J.; Basso, M.A. Parvalbumin and GABA Microcircuits in the Mouse Superior Colliculus. Front. Neural Circuits 2018, 12, 35. [Google Scholar] [CrossRef]
- Mao, J.B.; Evinger, C. Long-Term Potentiation of the Human Blink Reflex. J. Neurosci. 2001, 21, RC151. [Google Scholar] [CrossRef]
- Kubanek, J. Neuromodulation with Transcranial Focused Ultrasound. Neurosurg. Focus 2018, 44, E14. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Shapiro, M.G.; Tsao, D.Y. Ultrasonic Neuromodulation Causes Widespread Cortical Activation via an Indirect Auditory Mechanism. Neuron 2018, 98, 1031–1041.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiller, P.H.; True, S.D.; Conway, J.L. Effects of Frontal Eye Field and Superior Colliculus Ablations on Eye Movements. Science 1979, 206, 590–592. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors and Interactions | F | d,f | p |
|---|---|---|---|
| ‘SIDE’ | 11.92 | 1,15 | <0.01 |
| ‘CONDITION’ | 1.02 | 3,45 | 0.39 |
| ‘TIME-POINT’ | 2.09 | 2,30 | 0.14 |
| ‘ISI’ | 42.51 | 1,15 | <0.001 |
| ‘SIDE’ × ‘CONDITION’ | 0.13 | 3,45 | 0.94 |
| ‘SIDE’ × ‘TIME-POINT’ | 0.16 | 2,30 | 0.86 |
| ‘CONDITION’ × ‘TIME-POINT’ | 2.92 | 6,90 | 0.01 |
| ‘SIDE’ × ‘ISI’ | 2.40 | 1,15 | 0.14 |
| ‘CONDITION’ × ‘ISI’ | 1.55 | 3,45 | 0.21 |
| ‘TIME-POINT’ × ‘ISI’ | 1.03 | 2,30 | 0.37 |
| ‘SIDE’ × ‘CONDITION’ × ‘TIME-POINT’ | 0.70 | 6,90 | 0.65 |
| ‘SIDE’ × ‘CONDITION’ × ‘ISI’ | 2.53 | 3,45 | 0.07 |
| ‘SIDE’ × ‘TIME-POINT’ × ‘ISI’ | 1.16 | 2,30 | 0.33 |
| ‘CONDITION’ × ‘TIME-POINT’ × ‘ISI’ | 0.69 | 6,90 | 0.66 |
| ‘SIDE’ × ‘CONDITION’ × ‘TIME-POINT’ × ‘ISI’ | 1.33 | 6,90 | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerra, A.; Vicenzini, E.; Cioffi, E.; Colella, D.; Cannavacciuolo, A.; Pozzi, S.; Caccia, B.; Paparella, G.; Di Stefano, G.; Berardelli, A.; et al. Effects of Transcranial Ultrasound Stimulation on Trigeminal Blink Reflex Excitability. Brain Sci. 2021, 11, 645. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11050645
Guerra A, Vicenzini E, Cioffi E, Colella D, Cannavacciuolo A, Pozzi S, Caccia B, Paparella G, Di Stefano G, Berardelli A, et al. Effects of Transcranial Ultrasound Stimulation on Trigeminal Blink Reflex Excitability. Brain Sciences. 2021; 11(5):645. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11050645
Chicago/Turabian StyleGuerra, Andrea, Edoardo Vicenzini, Ettore Cioffi, Donato Colella, Antonio Cannavacciuolo, Silvia Pozzi, Barbara Caccia, Giulia Paparella, Giulia Di Stefano, Alfredo Berardelli, and et al. 2021. "Effects of Transcranial Ultrasound Stimulation on Trigeminal Blink Reflex Excitability" Brain Sciences 11, no. 5: 645. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11050645