
Effect of Xenon Treatment on Gene Expression in Brain Tissue after Traumatic Brain Injury in Rats

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. TBI
2.3. Xenon Treatment
2.4. Limb-Placing Test
2.5. Sampling
2.6. RNA Purification
2.7. Multiplex Gene Expression Analysis
2.8. Statistics
3. Results
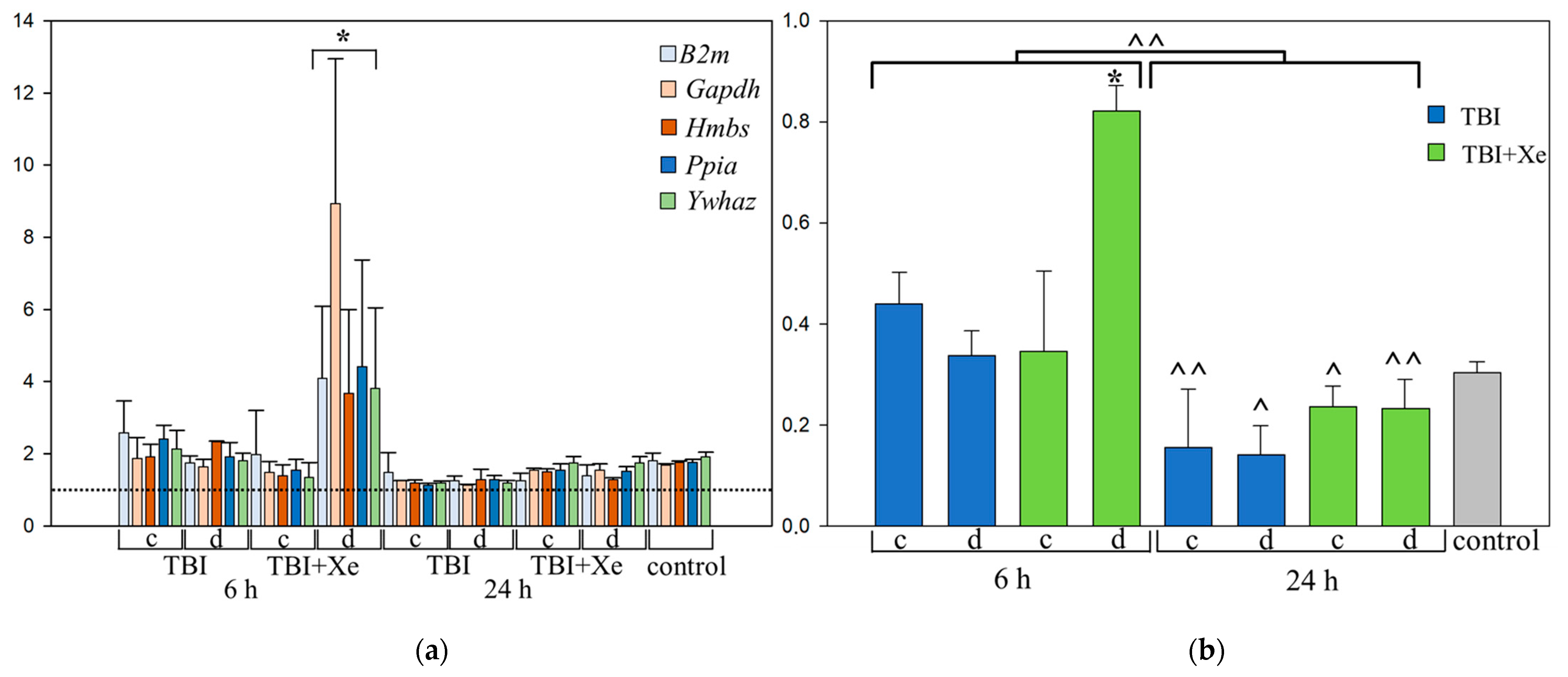
3.1. Housekeeping Gene Selection
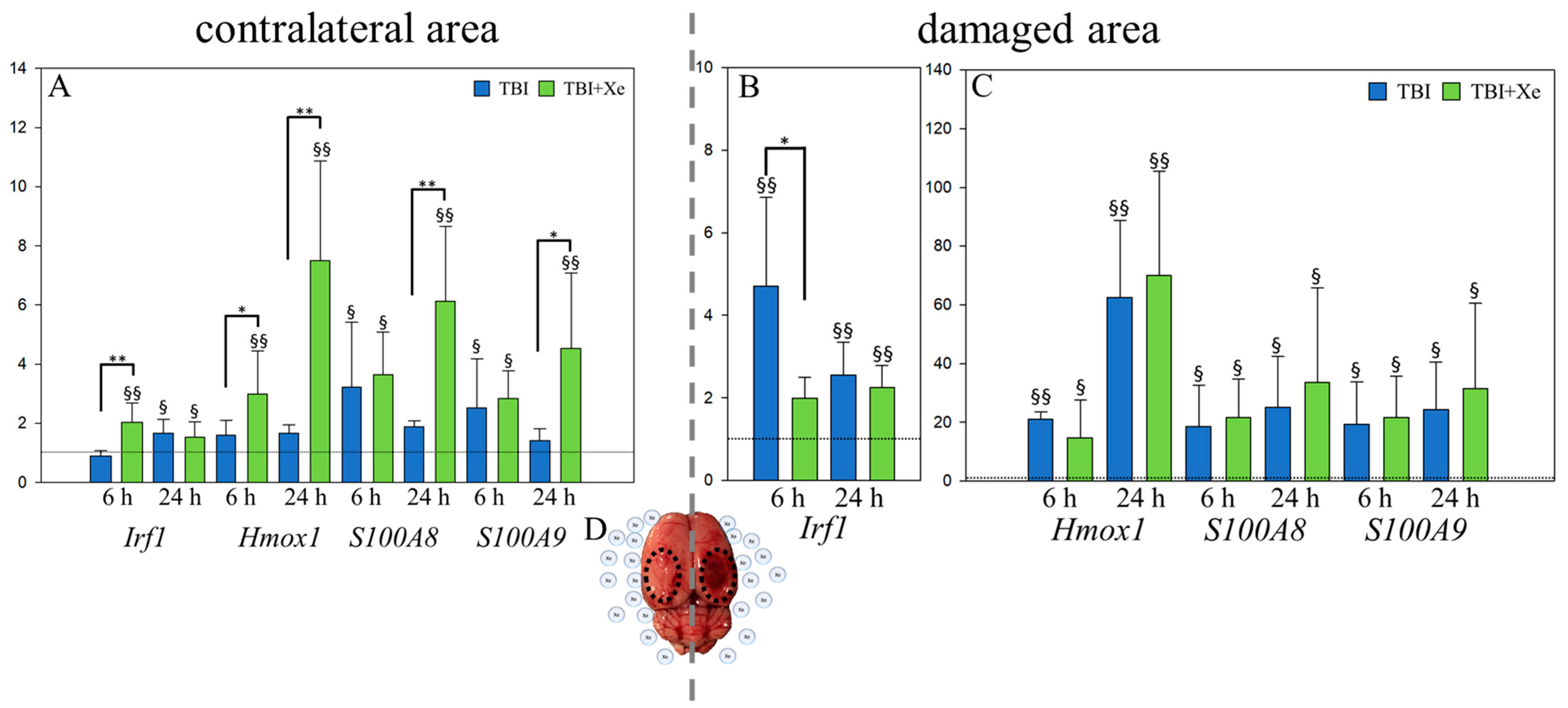
3.2. Xenon and Targeted Genes
3.3. Limb-Placing Test
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Wang, Y.Z.; Li, T.T.; Cao, H.L.; Yang, W.C. Recent advances in the neuroprotective effects of medical gases. Med. Gas Res. 2019, 9, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, R.; Franks, N.P. Bench-to-bedside review: Molecular pharmacology and clinical use of inert gases in anesthesia and neuroprotection. Crit. Care 2010, 14, 229. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, J.H.; Loomis, W.F.; Tobias, C.A.; Turpin, F.H. Preliminary observations on the narcotic effect of xenon with a review of values for solubilities of gases in water and oils. J. Physiol. 1946, 105, 197–204. [Google Scholar] [CrossRef]
- Jin, Z.; Piazza, O.; Ma, D.; Scarpati, G.; De Robertis, E. Xenon anesthesia and beyond: Pros and cons. Minerva Anestesiol. 2019, 85, 83–89. [Google Scholar] [CrossRef]
- Amer, A.R.; Oorschot, D.E. Xenon Combined with Hypothermia in Perinatal Hypoxic-Ischemic Encephalopathy: A Noble Gas, a Noble Mission. Pediatr. Neurol. 2018, 84, 5–10. [Google Scholar] [CrossRef]
- Campos-Pires, R.; Hirnet, T.; Valeo, F.; Ong, B.E.; Radyushkin, K.; Aldhoun, J.; Saville, J.; Edge, C.J.; Franks, N.P.; Thal, S.C.; et al. Xenon improves long-term cognitive function, reduces neuronal loss and chronic neuroinflammation, and improves survival after traumatic brain injury in mice. Br. J. Anaesth. 2019, 123, 60–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veldeman, M.; Coburn, M.; Rossaint, R.; Clusmann, H.; Nolte, K.; Kremer, B.; Höllig, A. Xenon Reduces Neuronal Hippocampal Damage and Alters the Pattern of Microglial Activation after Experimental Subarachnoid Hemorrhage: A Randomized Controlled Animal Trial. Front. Neurol. 2017, 8, 511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos-Pires, R.; Onggradito, H.; Ujvari, E.; Karimi, S.; Valeo, F.; Aldhoun, J.; Edge, C.J.; Franks, N.P.; Dickinson, R. Xenon treatment after severe traumatic brain injury improves locomotor outcome, reduces acute neuronal loss and enhances early beneficial neuroinflammation: A randomized, blinded, controlled animal study. Crit. Care 2020, 24, 667. [Google Scholar] [CrossRef]
- Franks, N.P.; Dickinson, R.; de Sousa, S.L.; Hall, A.C.; Lieb, W.R. How does xenon produce anaesthesia? Nature 1998, 396, 324. [Google Scholar] [CrossRef]
- Dickinson, R.; Peterson, B.K.; Banks, P.; Simillis, C.; Martin, J.C.; Valenzuela, C.A.; Maze, M.; Franks, N.P. Competitive inhibition at the glycine site of the N-methyl-D-aspartate receptor by the anesthetics xenon and isoflurane: Evidence from molecular modeling and electrophysiology. Anesthesiology 2007, 107, 756–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruss, M.; Bushell, T.J.; Bright, D.P.; Lieb, W.R.; Mathie, A.; Franks, N.P. Two-pore-domain K+ channels are a novel target for the anesthetic gases xenon, nitrous oxide, and cyclopropane. Mol. Pharmacol. 2004, 65, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bantel, C.; Maze, M.; Trapp, S. Noble gas xenon is a novel adenosine triphosphate-sensitive potassium channel opener. Anesthesiology 2010, 112, 623–630. [Google Scholar] [CrossRef] [Green Version]
- Fahlenkamp, A.V.; Coburn, M.; Haase, H.; Kipp, M.; Ryang, Y.M.; Rossaint, R.; Beyer, C. Xenon enhances LPS-induced IL-1β expression in microglia via the extracellular signal-regulated kinase 1/2 pathway. J. Mol. Neurosci. 2011, 45, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Lavaur, J.; Le Nogue, D.; Lemaire, M.; Pype, J.; Farjot, G.; Hirsch, E.C.; Michel, P.P. The noble gas xenon provides protection and trophic stimulation to midbrain dopamine neurons. J. Neurochem. 2017, 142, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhao, H.; Weng, H.; Ma, D. Lasting effects of general anesthetics on the brain in the young and elderly: “mixed picture” of neurotoxicity, neuroprotection and cognitive impairment. J. Anesth. 2019, 33, 321–335. [Google Scholar] [CrossRef] [Green Version]
- Breuer, T.; Emontzpohl, C.; Coburn, M.; Benstoem, C.; Rossaint, R.; Marx, G.; Schälte, G.; Bernhagen, J.; Bruells, C.S.; Goetzenich, A.; et al. Xenon triggers pro-inflammatory effects and suppresses the anti-inflammatory response compared to sevoflurane in patients undergoing cardiac surgery. Crit. Care 2015, 19, 365. [Google Scholar] [CrossRef] [Green Version]
- Feeney, D.M.; Boyeson, M.G.; Linn, R.T.; Murray, H.M.; Dail, W.G. Responses to cortical injury: I. Methodology and local effects of contusions in the rat. Brain Res. 1981, 211, 67–77. [Google Scholar] [CrossRef]
- Jolkkonen, J.; Puurunen, K.; Rantakömi, S.; Härkönen, A.; Haapalinna, A.; Sivenius, J. Behavioral effects of the α2-adrenoceptor antagonist, atipamezole, after focal cerebral ischemia in rats. Eur. J. Pharmacol. 2000, 400, 211–219. [Google Scholar] [CrossRef]
- Sundaram, V.K.; Sampathkumar, N.K.; Massaad, C.; Grenier, J. Optimal use of statistical methods to validate reference gene stability in longitudinal studies. PLoS ONE 2019, 14, e0219440. [Google Scholar] [CrossRef] [Green Version]
- Gubern, C.; Hurtado, O.; Rodríguez, R.; Morales, J.R.; Romera, V.G.; Moro, M.A.; Lizasoain, I.; Serena, J.; Mallolas, J. Validation of housekeeping genes for quantitative real-time PCR in in-vivo and in-vitro models of cerebral ischaemia. BMC Mol. Biol. 2009, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Swijsen, A.; Nelissen, K.; Janssen, D.; Rigo, J.M.; Hoogland, G. Validation of reference genes for quantitative real-time PCR studies in the dentate gyrus after experimental febrile seizures. BMC Res. Notes 2012, 5, 685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gholami, K.; Loh, S.Y.; Salleh, N.; Lam, S.K.; Hoe, S.Z. Selection of suitable endogenous reference genes for qPCR in kidney and hypothalamus of rats under testosterone influence. PLoS ONE 2017, 12, e0176368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Iashchishyn, I.A.; Kara, J.; Foderà, V.; Vetri, V.; Sancataldo, G.; Marklund, N.; Morozova-Roche, L.A. Proinflammatory and amyloidogenic S100A9 induced by traumatic brain injury in mouse model. Neurosci. Lett. 2019, 699, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Ping, S.; Kyle, M.; Longo, J.; Chin, L.; Zhao, L.R. S100 Calcium-Binding Protein A9 Knockout Contributes to Neuroprotection and Functional Improvement after Traumatic Brain Injury. J. Neurotrauma 2020, 37, 950–965. [Google Scholar] [CrossRef]
- Sun, J.; Pan, X.; Christiansen, L.I.; Yuan, X.L.; Skovgaard, K.; Chatterton, D.E.W.; Kaalund, S.S.; Gao, F.; Sangild, P.T.; Pankratova, S. Necrotizing enterocolitis is associated with acute brain responses in preterm pigs. J. Neuroinflamm. 2018, 15, 180. [Google Scholar] [CrossRef]
- Khlusov, I.A.; Naumov, S.A.; Vovk, S.M.; Kornetov, N.A.; Shpisman, M.N.; Lukinov, A.V.; Naumov, A.V. Vliianie ksenona na kletki i retseptory [Xenon effects on cells and receptors]. Vestn. Ross. Akad. Med. Nauk. 2003, 9, 32–37. (In Russian) [Google Scholar]
- Armstrong, S.P.; Banks, P.J.; McKitrick, T.J.; Geldart, C.H.; Edge, C.J.; Babla, R.; Simillis, C.; Franks, N.P.; Dickinson, R. Identification of two mutations (F758W and F758Y) in the N-methyl-D-aspartate receptor glycine-binding site that selectively prevent competitive inhibition by xenon without affecting glycine binding. Anesthesiology 2012, 117, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyllie, J.; Bruinenberg, J.; Roehr, C.C.; Rüdiger, M.; Trevisanuto, D.; Urlesberger, B. European Resuscitation Council Guidelines for Resuscitation 2015: Section 7. Resuscitation and support of transition of babies at birth. Resuscitation 2015, 95, 249–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amano, T.; Meyer, J.S.; Okabe, T.; Shaw, T.; Mortel, K.F. Stable xenon CT cerebral blood flow measurements computed by a single compartment--double integration model in normal aging and dementia. J. Comput. Assist. Tomogr. 1982, 6, 923–932. [Google Scholar] [CrossRef]
- Gerstner, B. Interaction of inflammation and hyperoxia in a rat model of neonatal white matter damage. PLoS ONE 2012, 7, e49023. [Google Scholar]
- Wright, C.J.; Dennery, P.A. Manipulation of gene expression by oxygen: A primer from bedside to bench. Pediatr. Res. 2009, 66, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greco, F.; Wiegert, S.; Baumann, P.; Wellmann, S.; Pellegrini, G.; Cannizzaro, V. Hyperoxia-induced lung structure-function relation, vessel rarefaction, and cardiac hypertrophy in an infant rat model. J. Transl. Med. 2019, 17, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HKG | CV% | SD | Rank |
|---|---|---|---|
| Gapdh | 28.6 | 20.2 | 1 |
| Ppia | 28.8 | 22.1 | 2 |
| Ywhaz | 29.8 | 18.5 | 3 |
| Hmbs | 30.1 | 18.2 | 4 |
| B2m | 40.0 | 19.1 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filev, A.D.; Silachev, D.N.; Ryzhkov, I.A.; Lapin, K.N.; Babkina, A.S.; Grebenchikov, O.A.; Pisarev, V.M. Effect of Xenon Treatment on Gene Expression in Brain Tissue after Traumatic Brain Injury in Rats. Brain Sci. 2021, 11, 889. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070889
Filev AD, Silachev DN, Ryzhkov IA, Lapin KN, Babkina AS, Grebenchikov OA, Pisarev VM. Effect of Xenon Treatment on Gene Expression in Brain Tissue after Traumatic Brain Injury in Rats. Brain Sciences. 2021; 11(7):889. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070889
Chicago/Turabian StyleFilev, Anton D., Denis N. Silachev, Ivan A. Ryzhkov, Konstantin N. Lapin, Anastasiya S. Babkina, Oleg A. Grebenchikov, and Vladimir M. Pisarev. 2021. "Effect of Xenon Treatment on Gene Expression in Brain Tissue after Traumatic Brain Injury in Rats" Brain Sciences 11, no. 7: 889. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070889