
Disrupted Functional Rich-Club Organization of the Brain Networks in Children with Attention-Deficit/Hyperactivity Disorder, a Resting-State EEG Study

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
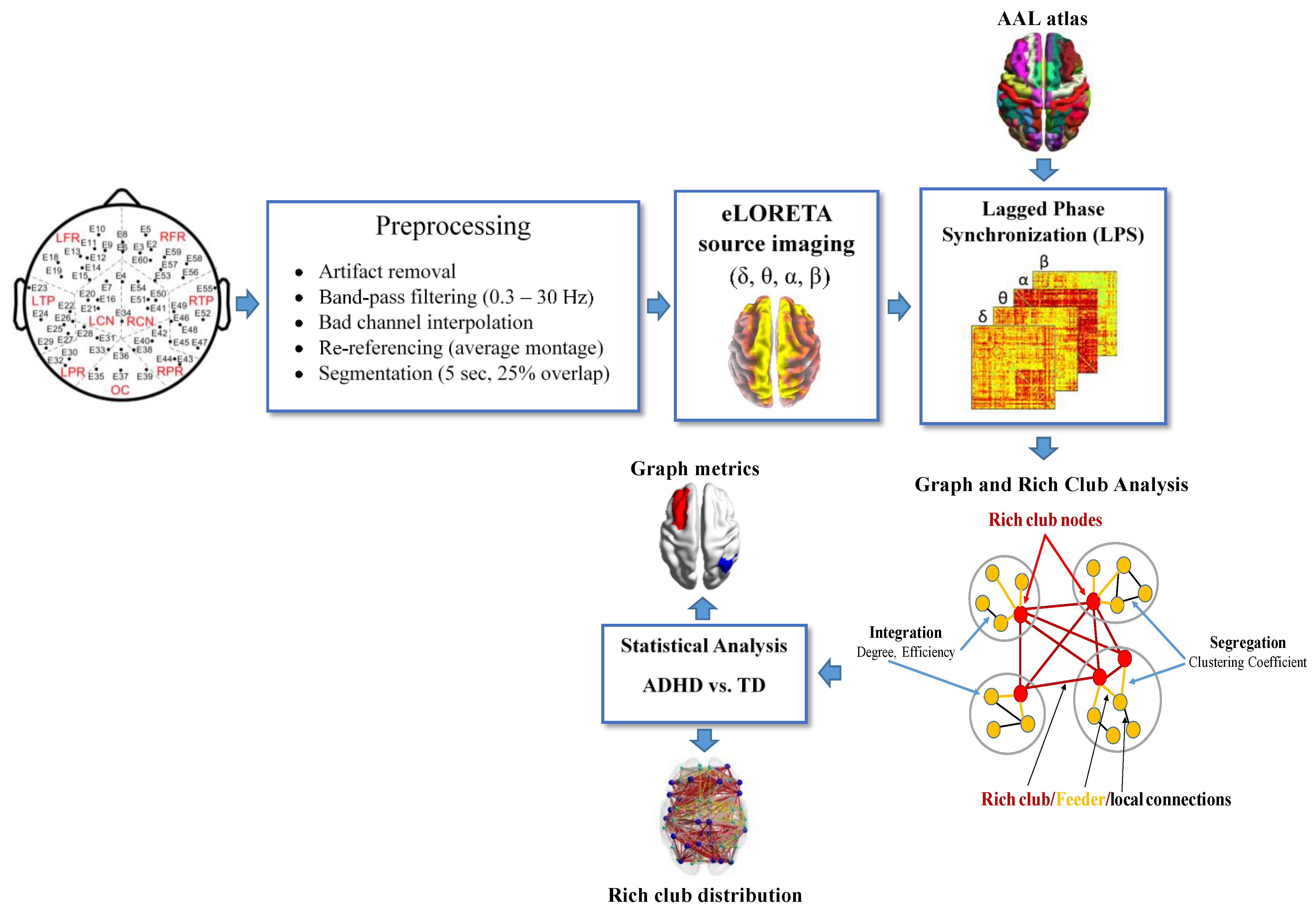
2. Materials and Methods
2.1. Participants
2.2. Data Acquisition and Preprocessing
2.3. EEG Source Analysis
2.4. Functional Connectivity Analysis
2.5. Rich-Club Organization
2.6. Statistical Analysis
3. Results
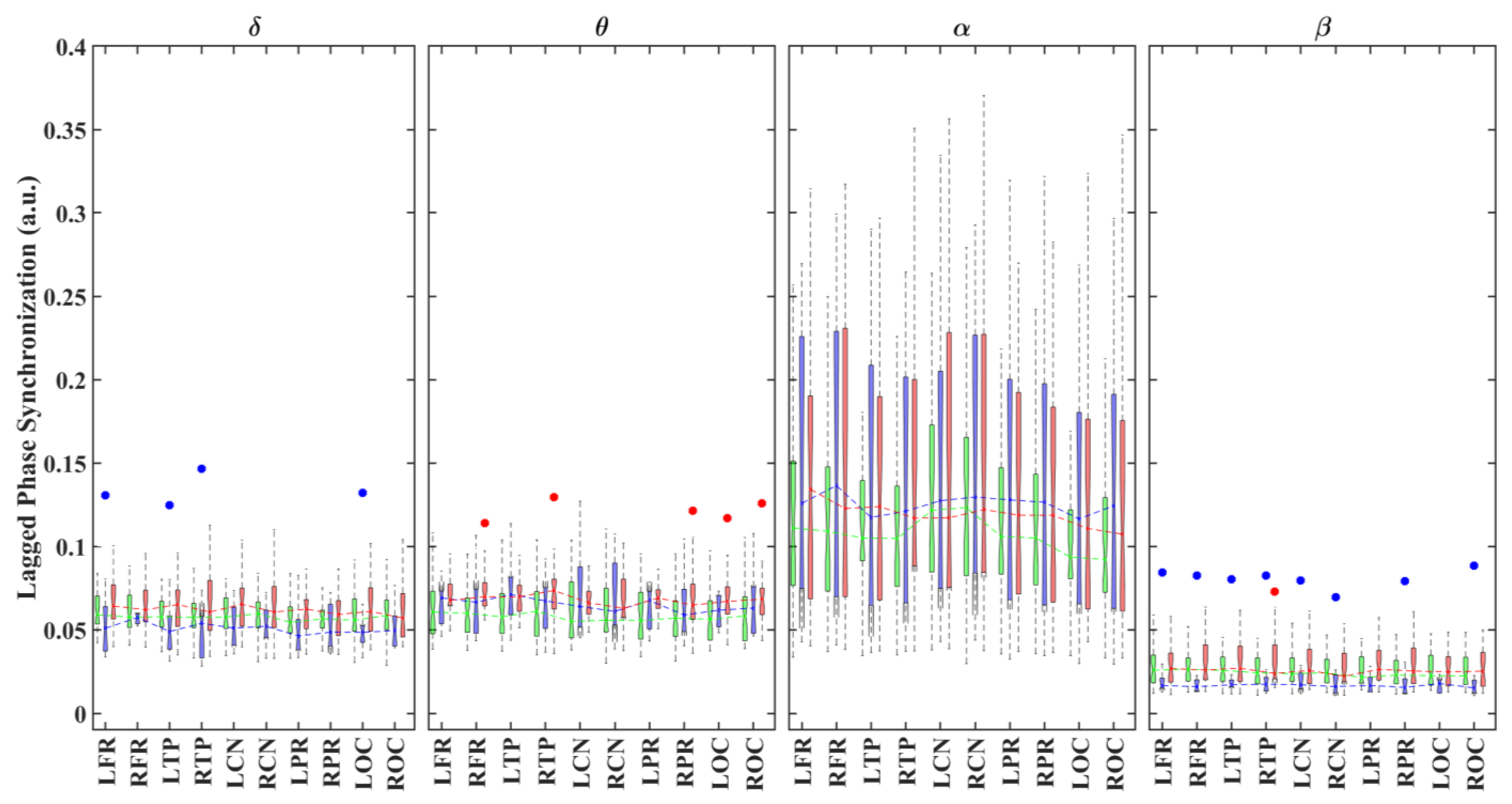
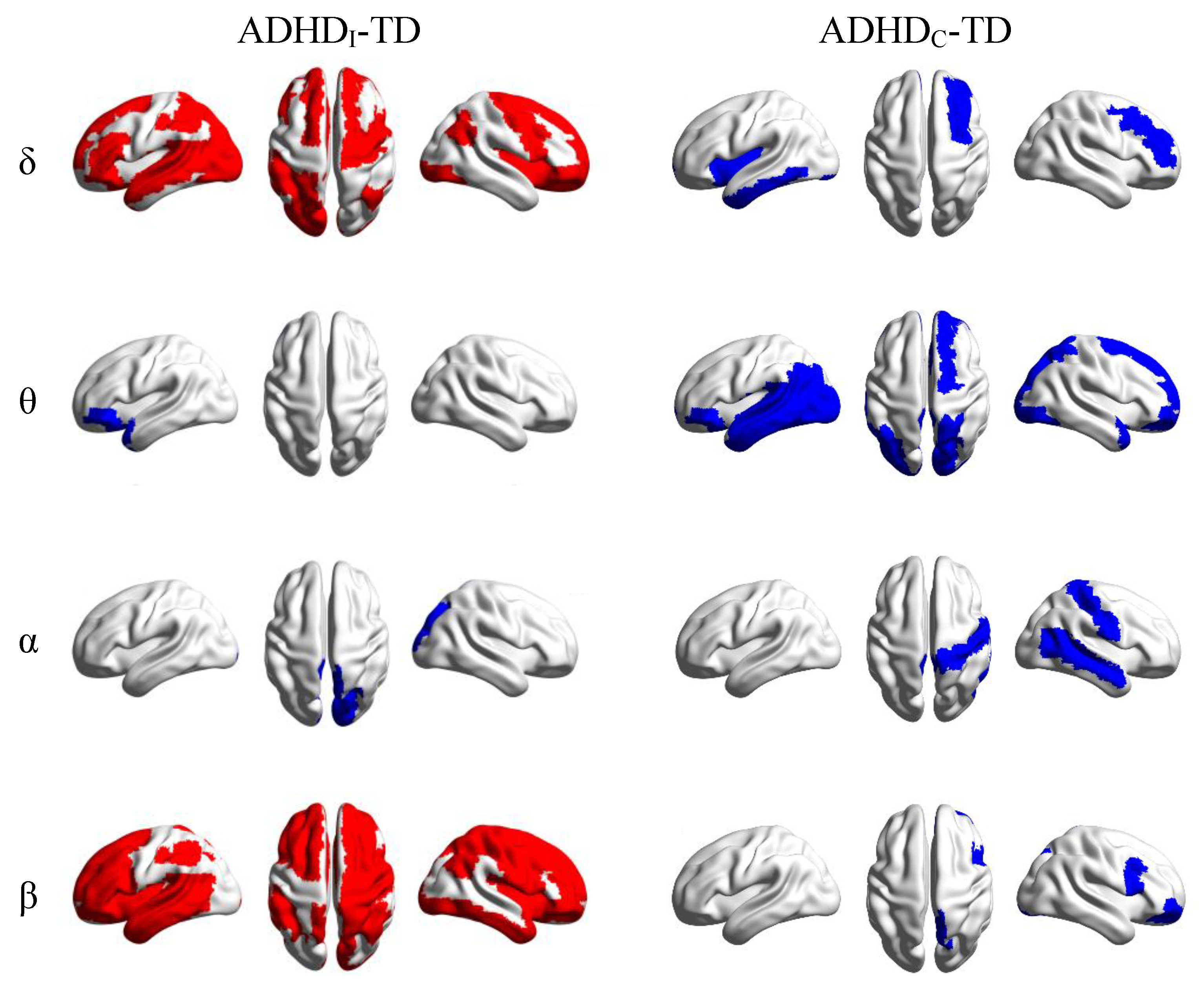
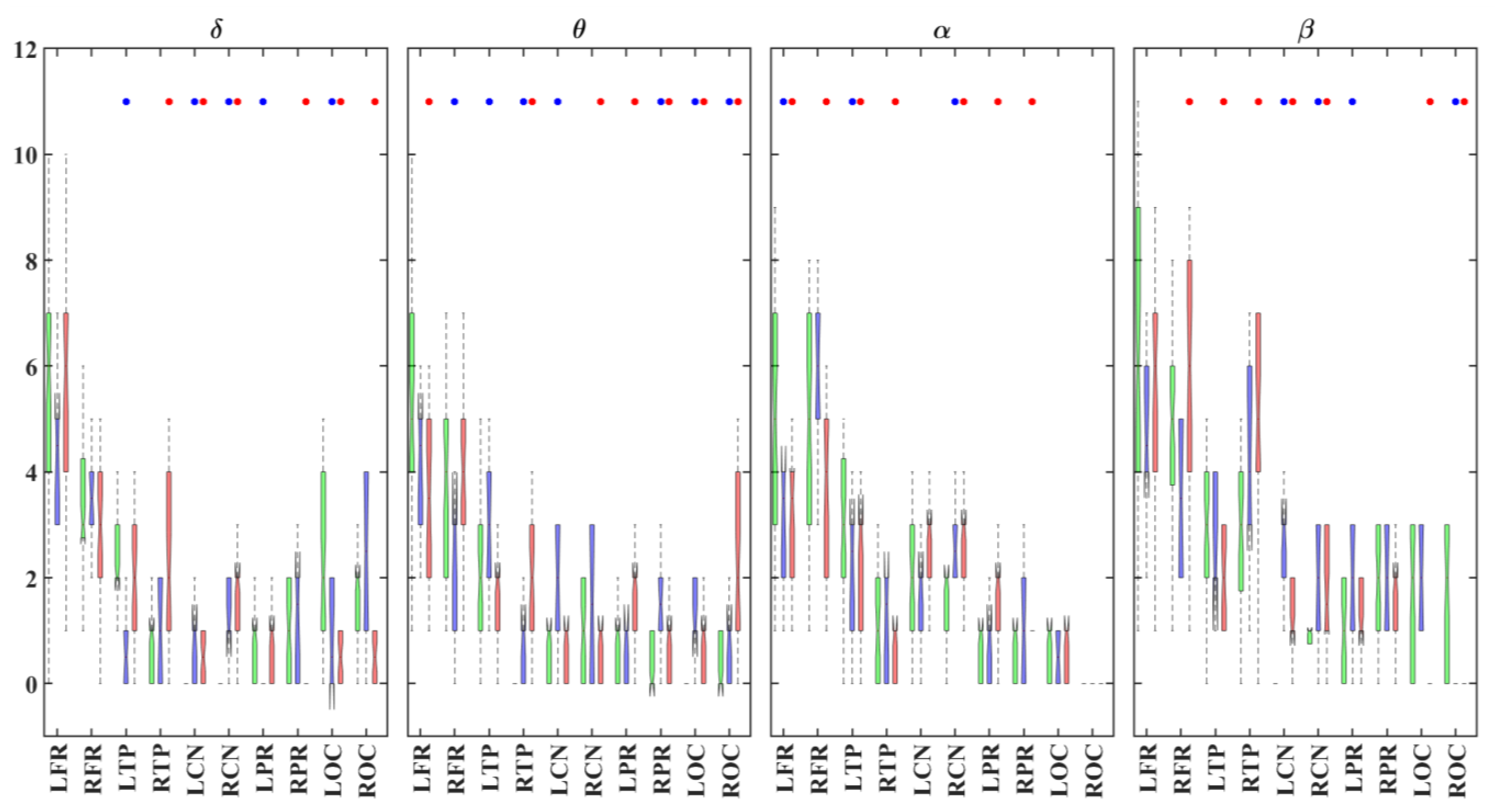
3.1. Alterations in Functional Connectivity
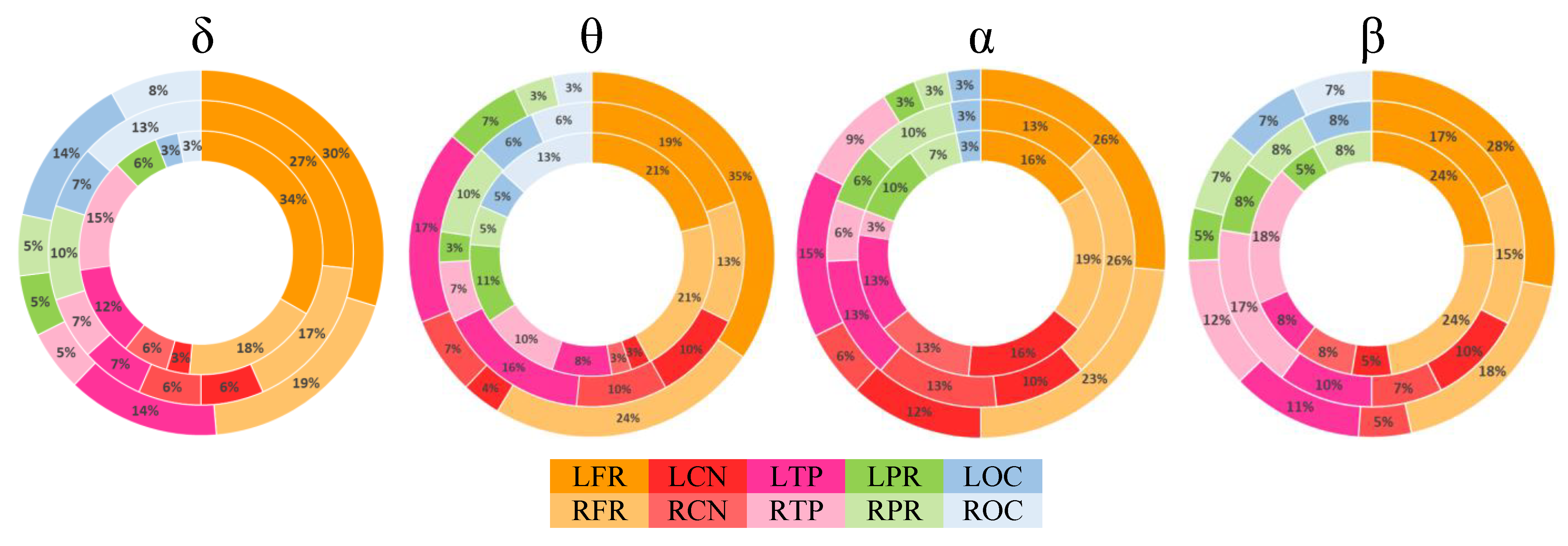
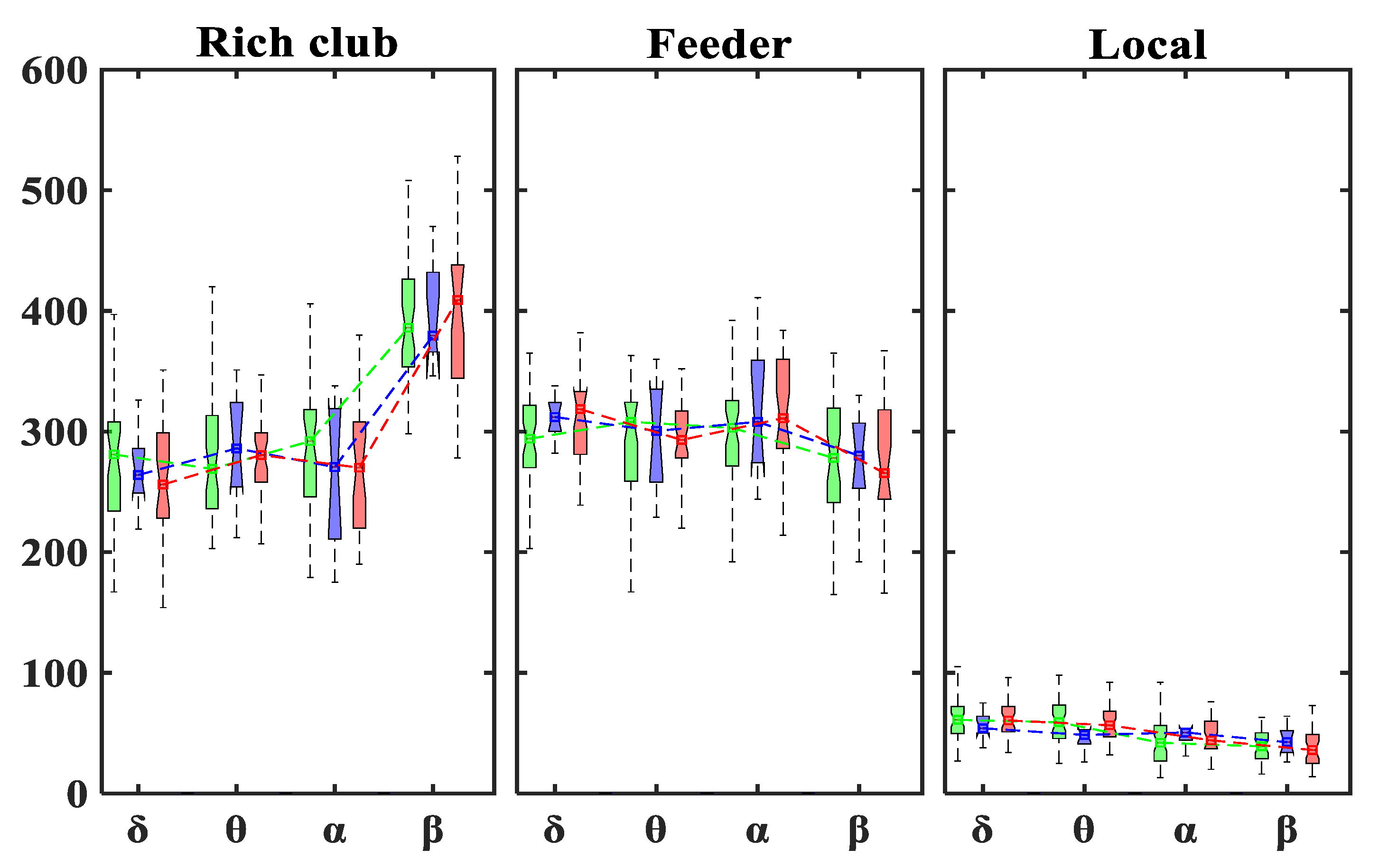
3.2. Rich-Club Organization
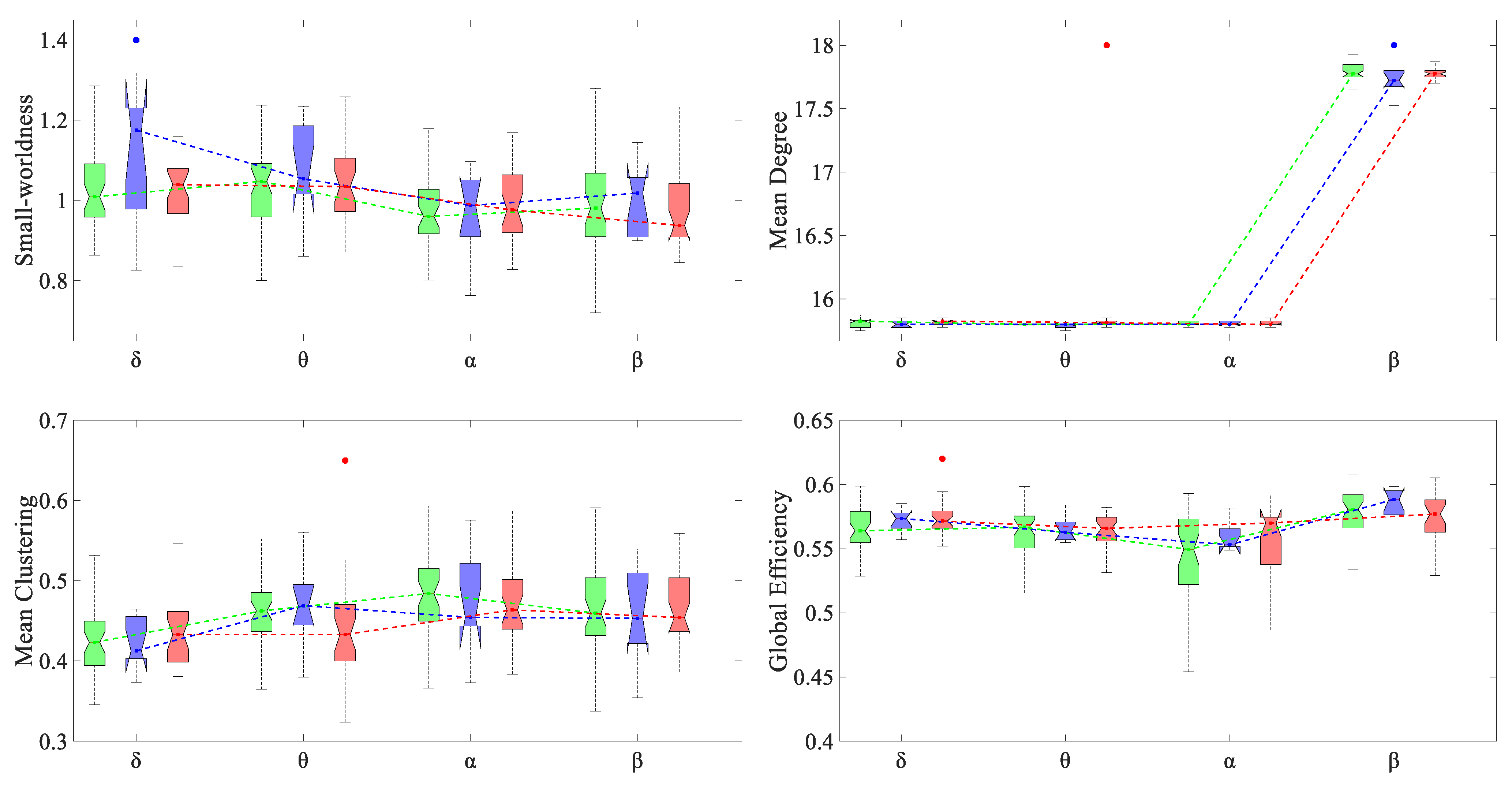
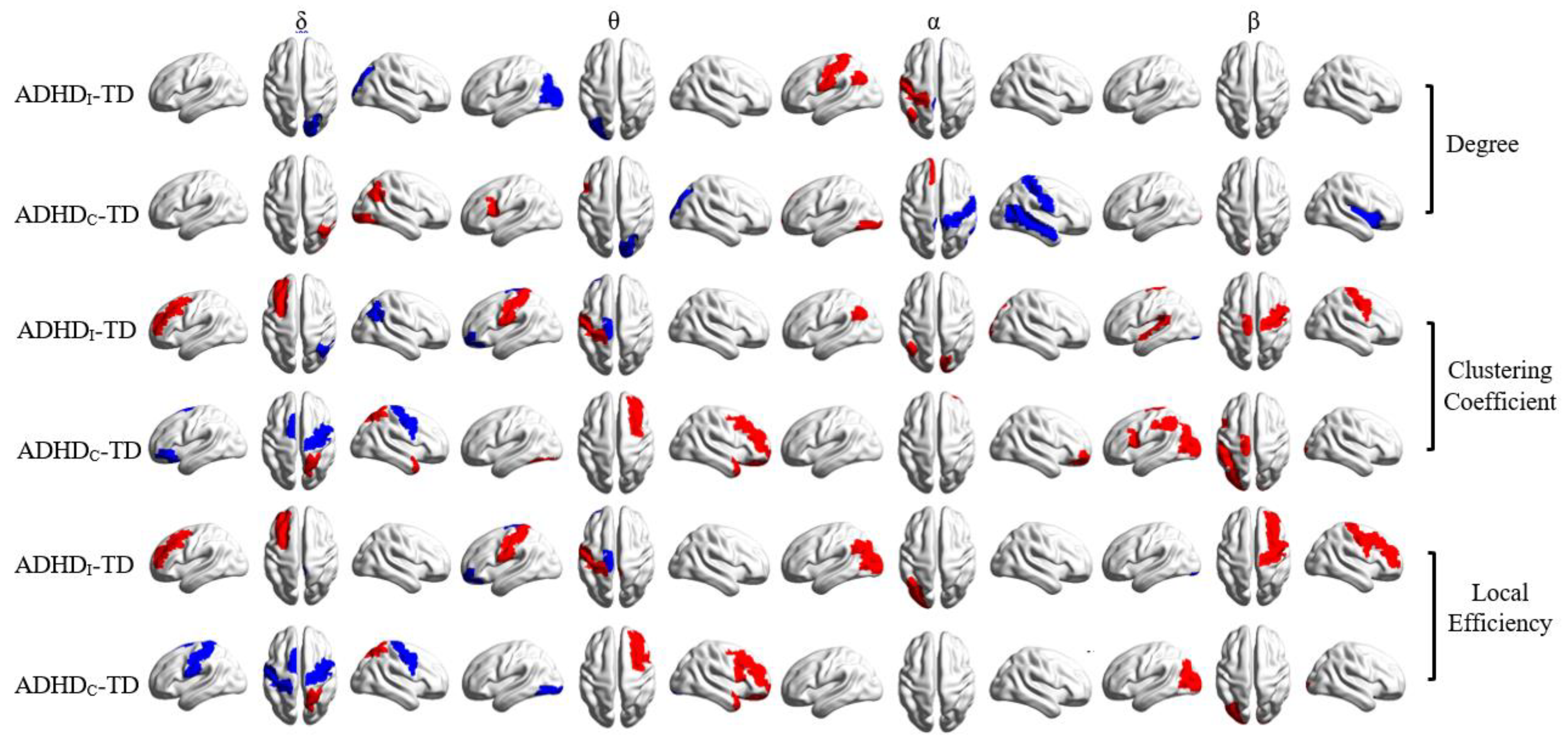
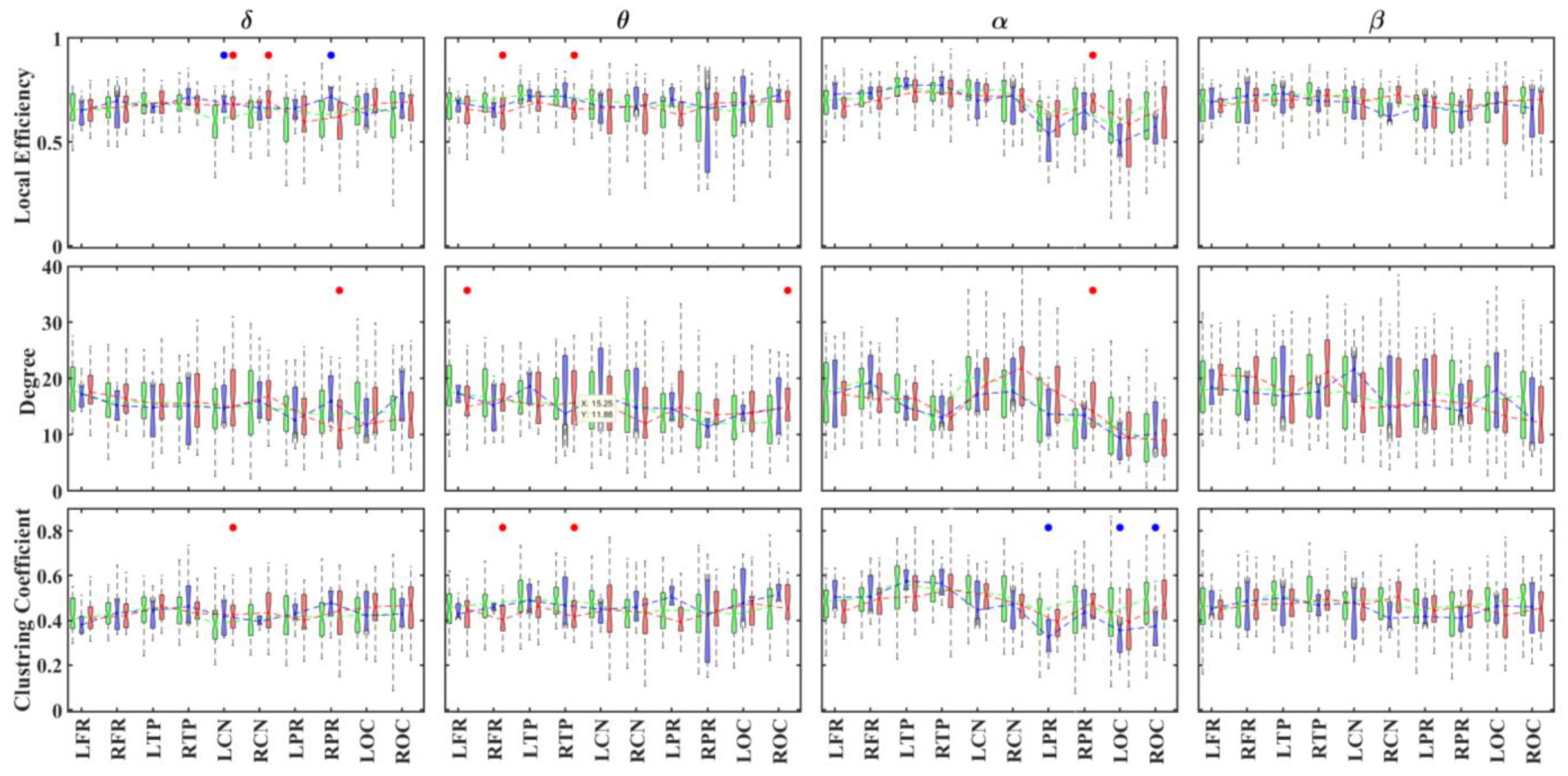
3.3. Alteration in Global and Local Topological Properties
4. Discussion
Technical Considerations and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bálint, S.; Czobor, P.; Komlósi, S.; Meszaros, A.; Simon, V.; Bitter, I. Attention deficit hyperactivity disorder (ADHD): Gender-and age-related differences in neurocognition. Psychol. Med. 2009, 39, 1337. [Google Scholar] [CrossRef] [PubMed]
- Ayano, G.; Yohannes, K.; Abraha, M. Epidemiology of attention-deficit/hyperactivity disorder (ADHD) in children and adolescents in Africa: A systematic review and meta-analysis. Ann. Gen. Psychiatry 2020, 19, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Text Revision, 4th ed.; American Psychiatric Association: Washington DC, USA, 2000.
- Nigg, J.T. Is ADHD a disinhibitory disorder? Psychol. Bull. 2001, 127, 571. [Google Scholar] [CrossRef]
- Ahmadi, M.; Kazemi, K.; Kuc, K.; Cybulska-Klosowicz, A.; Zakrzewska, M.; Racicka-Pawlukiewicz, E.; Helfroush, M.S.; Aarabi, A. Cortical source analysis of resting state EEG data in children with attention deficit hyperactivity disorder. Clin. Neurophysiol. 2020, 131, 2115–2130. [Google Scholar] [CrossRef]
- Newson, J.J.; Thiagarajan, T.C. EEG frequency bands in psychiatric disorders: A review of resting state studies. Front. Hum. Neurosci. 2019, 12, 521. [Google Scholar] [CrossRef] [PubMed]
- Furlong, S. Resting-State EEG Connectivity in Young Children with ADHD: A Potential Neural Marker. Ph.D. Thesis, The University of North Carolina at Chapel Hill, Chapel Hill, NC, USA, 2018. [Google Scholar]
- Michelini, G.; Jurgiel, J.; Bakolis, I.; Cheung, C.H.; Asherson, P.; Loo, S.K.; Kuntsi, J.; Mohammad-Rezazadeh, I. Atypical functional connectivity in adolescents and adults with persistent and remitted ADHD during a cognitive control task. Transl. Psychiatry 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Ghaderi, A.H.; Nazari, M.A.; Shahrokhi, H.; Darooneh, A.H. Functional brain connectivity differences between different ADHD presentations: Impaired functional segregation in ADHD-combined presentation but not in ADHD-inattentive presentation. Basic Clin. Neurosci. 2017, 8, 267. [Google Scholar] [CrossRef]
- Fair, D.A.; Posner, J.; Nagel, B.J.; Bathula, D.; Dias, T.G.C.; Mills, K.L.; Blythe, M.S.; Giwa, A.; Schmitt, C.F.; Nigg, J.T. Atypical default network connectivity in youth with attention-deficit/hyperactivity disorder. Biol. Psychiatry 2010, 68, 1084–1091. [Google Scholar] [CrossRef] [Green Version]
- Tomasi, D.; Volkow, N.D. Abnormal functional connectivity in children with attention-deficit/hyperactivity disorder. Biol. Psychiatry 2012, 71, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Jiang, T.; Wang, Y.; Zang, Y.; He, Y.; Liang, M.; Sui, M.; Cao, Q.; Hu, S.; Peng, M. Altered resting-state functional connectivity patterns of anterior cingulate cortex in adolescents with attention deficit hyperactivity disorder. Neurosci. Lett. 2006, 400, 39–43. [Google Scholar] [CrossRef]
- Peterson, B.S.; Potenza, M.N.; Wang, Z.; Zhu, H.; Martin, A.; Marsh, R.; Plessen, K.J.; Yu, S. An FMRI study of the effects of psychostimulants on default-mode processing during Stroop task performance in youths with ADHD. Am. J. Psychiatry 2009, 166, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Cubillo, A.; Halari, R.; Ecker, C.; Giampietro, V.; Taylor, E.; Rubia, K. Reduced activation and inter-regional functional connectivity of fronto-striatal networks in adults with childhood Attention-Deficit Hyperactivity Disorder (ADHD) and persisting symptoms during tasks of motor inhibition and cognitive switching. J. Psychiatr. Res. 2010, 44, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Monk, C.S.; Peltier, S.J.; Wiggins, J.L.; Weng, S.-J.; Carrasco, M.; Risi, S.; Lord, C. Abnormalities of intrinsic functional connectivity in autism spectrum disorders. Neuroimage 2009, 47, 764–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Tao, F.; Zuo, C.; Kanji, M.; Hu, M.; Wang, D. Disrupted Resting Frontal–Parietal Attention Network Topology is Associated with a Clinical Measure in Children with Attention-Deficit/Hyperactivity Disorder. Front. Psychiatry 2019, 10, 300. [Google Scholar] [CrossRef]
- Murias, M.; Webb, S.J.; Greenson, J.; Dawson, G. Resting state cortical connectivity reflected in EEG coherence in individuals with autism. Biol. Psychiatry 2007, 62, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.R.; Barry, R.J.; McCarthy, R.; Selikowitz, M.; Johnstone, S.J.; Hsu, C.-I.; Magee, C.A.; Lawrence, C.A.; Croft, R.J. Coherence in children with attention-deficit/hyperactivity disorder and excess beta activity in their EEG. Clin. Neurophysiol. 2007, 118, 1472–1479. [Google Scholar] [CrossRef]
- Dupuy, F.E.; Clarke, A.R.; Barry, R.J.; McCarthy, R.; Selikowitz, M. EEG coherence in girls with attention-deficit/hyperactivity disorder: Stimulant effects in good responders. Int. J. Psychophysiol. 2008, 70, 151–157. [Google Scholar] [CrossRef]
- Barry, R.J.; Clarke, A.R.; McCarthy, R.; Selikowitz, M.; Johnstone, S.J. EEG coherence adjusted for inter-electrode distance in children with attention-deficit/hyperactivity disorder. Int. J. Psychophysiol. 2005, 58, 12–20. [Google Scholar] [CrossRef]
- Liu, T.; Chen, Y.; Lin, P.; Wang, J. Small-world brain functional networks in children with attention-deficit/hyperactivity disorder revealed by EEG synchrony. Clin. EEG Neurosci. 2015, 46, 183–191. [Google Scholar] [CrossRef]
- Barry, R.J.; Clarke, A.R.; McCarthy, R.; Selikowitz, M. EEG coherence in attention-deficit/hyperactivity disorder: A comparative study of two DSM-IV types. Clin. Neurophysiol. 2002, 113, 579–585. [Google Scholar] [CrossRef]
- Wang, B.; Wang, G.; Wang, X.; Cao, R.; Xiang, J.; Yan, T.; Li, H.; Yoshimura, S.; Toichi, M.; Zhao, S. Rich-club analysis in adults with ADHD connectomes reveals an abnormal structural core network. J. Atten. Disord. 2021, 25, 1068–1079. [Google Scholar] [CrossRef]
- Ahmadlou, M.; Adeli, H.; Adeli, A. Graph theoretical analysis of organization of functional brain networks in ADHD. Clin. EEG Neurosci. 2012, 43, 5–13. [Google Scholar] [CrossRef]
- Cao, M.; Shu, N.; Cao, Q.; Wang, Y.; He, Y. Imaging functional and structural brain connectomics in attention-deficit/hyperactivity disorder. Mol. Neurobiol. 2014, 50, 1111–1123. [Google Scholar] [CrossRef]
- Van den Heuvel, M.P.; Sporns, O. An anatomical substrate for integration among functional networks in human cortex. J. Neurosci. 2013, 33, 14489–14500. [Google Scholar] [CrossRef]
- Colizza, V.; Flammini, A.; Serrano, M.A.; Vespignani, A. Detecting rich-club ordering in complex networks. Nat. Phys. 2006, 2, 110–115. [Google Scholar] [CrossRef]
- McAuley, J.J.; da Fontoura Costa, L.; Caetano, T.S. Rich-club phenomenon across complex network hierarchies. Appl. Phys. Lett. 2007, 91, 084103. [Google Scholar] [CrossRef] [Green Version]
- de Reus, M.A.; van den Heuvel, M.P. Rich club organization and intermodule communication in the cat connectome. J. Neurosci. 2013, 33, 12929–12939. [Google Scholar] [CrossRef] [Green Version]
- Collin, G.; de Nijs, J.; Pol, H.H.; Cahn, W.; van den Heuvel, M.P. Connectome organization is related to longitudinal changes in general functioning, symptoms and IQ in chronic schizophrenia. Schizophr. Res. 2016, 173, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, L.; Yin, Q.; Dun, W.; Xu, X.; Liu, J.; Zhang, M. Abnormal rich club organization and impaired correlation between structural and functional connectivity in migraine sufferers. Brain Imaging Behav. 2017, 11, 526–540. [Google Scholar] [CrossRef] [PubMed]
- Daianu, M.; Mezher, A.; Mendez, M.F.; Jahanshad, N.; Jimenez, E.E.; Thompson, P.M. Disrupted rich club network in behavioral variant frontotemporal dementia and early-onset A lzheimer’s disease. Hum. Brain Mapp. 2016, 37, 868–883. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Miller, M.; Karalunas, S.; Robertson, C.; Grayson, D.S.; Cary, R.P.; Hawkey, E.; Painter, J.G.; Kriz, D.; Fombonne, E. Structural and functional connectivity of the human brain in autism spectrum disorders and attention-deficit/hyperactivity disorder: A rich club-organization study. Hum. Brain Mapp. 2014, 35, 6032–6048. [Google Scholar] [CrossRef] [Green Version]
- Nunez, P.L.; Srinivasan, R.; Westdorp, A.F.; Wijesinghe, R.S.; Tucker, D.M.; Silberstein, R.B.; Cadusch, P.J. EEG coherency: I: Statistics, reference electrode, volume conduction, Laplacians, cortical imaging, and interpretation at multiple scales. Electroencephalogr. Clin. Neurophysiol. 1997, 103, 499–515. [Google Scholar] [CrossRef]
- Nolte, G.; Bai, O.; Wheaton, L.; Mari, Z.; Vorbach, S.; Hallett, M. Identifying true brain interaction from EEG data using the imaginary part of coherency. Clin. Neurophysiol. 2004, 115, 2292–2307. [Google Scholar] [CrossRef]
- Giertuga, K.; Zakrzewska, M.Z.; Bielecki, M.; Racicka-Pawlukiewicz, E.; Kossut, M.; Cybulska-Klosowicz, A. Age-related changes in resting-state EEG activity in attention deficit/hyperactivity disorder: A cross-sectional study. Front. Hum. Neurosci. 2017, 11, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuPaul, G.J.; Power, T.J.; Anastopoulos, A.D.; Reid, R. ADHD Rating Scale—IV: Checklists, Norms, and Clinical Interpretation; Guilford Press: New York, NY, USA, 1998. [Google Scholar]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput. Intell. Neurosci. 2011, 2011. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods Find. Exp. Clin. Pharm. 2002, 24, 5–12. [Google Scholar]
- Luu, P.; Ferree, T. Determination of the HydroCel Geodesic Sensor Nets’ Average Electrode Positions and Their 10–10 International Equivalents; Electrical Geodesics, Inc.: Eugene, OR, USA, 2005; pp. 1–11. [Google Scholar]
- Mazziotta, J.; Toga, A.; Evans, A.; Fox, P.; Lancaster, J.; Zilles, K.; Woods, R.; Paus, T.; Simpson, G.; Pike, B. A probabilistic atlas and reference system for the human brain: International Consortium for Brain Mapping (ICBM). Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2001, 356, 1293–1322. [Google Scholar] [CrossRef] [PubMed]
- Zinn, M.; Zinn, M.; Valencia, I.; Jason, L.; Montoya, J. Cortical hypoactivation during resting EEG suggests central nervous system pathology in patients with Chronic Fatigue Syndrome. Biol. Psychol. 2018, 136, 87–99. [Google Scholar] [CrossRef]
- Paquette, V.; Beauregard, M.; Beaulieu-Prévost, D. Effect of a psychoneurotherapy on brain electromagnetic tomography in individuals with major depressive disorder. Psychiatry Res. Neuroimaging 2009, 174, 231–239. [Google Scholar] [CrossRef]
- Rizkallah, J.; Amoud, H.; Fraschini, M.; Wendling, F.; Hassan, M. Exploring the correlation between M/EEG source–space and fMRI networks at rest. Brain Topogr. 2020, 33, 151–160. [Google Scholar] [CrossRef]
- Stam, C.J.; Jones, B.; Nolte, G.; Breakspear, M.; Scheltens, P. Small-world networks and functional connectivity in Alzheimer’s disease. Cereb. Cortex 2007, 17, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Canuet, L.; Ishii, R.; Pascual-Marqui, R.D.; Iwase, M.; Kurimoto, R.; Aoki, Y.; Ikeda, S.; Takahashi, H.; Nakahachi, T.; Takeda, M. Resting-state EEG source localization and functional connectivity in schizophrenia-like psychosis of epilepsy. PLoS ONE 2011, 6, e27863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassett, D.S.; Bullmore, E.T.; Meyer-Lindenberg, A.; Apud, J.A.; Weinberger, D.R.; Coppola, R. Cognitive fitness of cost-efficient brain functional networks. Proc. Natl. Acad. Sci. USA 2009, 106, 11747–11752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drakesmith, M.; Caeyenberghs, K.; Dutt, A.; Lewis, G.; David, A.; Jones, D.K. Overcoming the effects of false positives and threshold bias in graph theoretical analyses of neuroimaging data. Neuroimage 2015, 118, 313–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Khalilian, M.; Kazemi, K.; Fouladivanda, M.; Makki, M.; Helfroush, M.S.; Aarabi, A. Effect of Multishell Diffusion MRI Acquisition Strategy and Parcellation Scale on Rich-Club Organization of Human Brain Structural Networks. Diagnostics 2021, 11, 970. [Google Scholar] [CrossRef] [PubMed]
- Harriger, L.; Van Den Heuvel, M.P.; Sporns, O. Rich club organization of macaque cerebral cortex and its role in network communication. PLoS ONE 2012, 7, e46497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouladivanda, M.; Kazemi, K.; Makki, M.; Khalilian, M.; Danyali, H.; Gervain, J.; Aarabi, A. Multi-scale structural rich-club organization of the brain in full-term newborns: A combined DWI and fMRI study. J. Neural Eng. 2021, 18, 046065. [Google Scholar] [CrossRef] [PubMed]
- Van Den Heuvel, M.P.; Sporns, O.; Collin, G.; Scheewe, T.; Mandl, R.C.; Cahn, W.; Goñi, J.; Pol, H.E.H.; Kahn, R.S. Abnormal rich club organization and functional brain dynamics in schizophrenia. JAMA Psychiatry 2013, 70, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, B.C.; Chen, Z.; He, Y.; Evans, A.C.; Bernasconi, N. Graph-theoretical analysis reveals disrupted small-world organization of cortical thickness correlation networks in temporal lobe epilepsy. Cereb. Cortex 2011, 21, 2147–2157. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A network visualization tool for human brain connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.R.; Barry, R.J.; McCarthy, R.; Selikowitz, M. EEG analysis in attention-deficit/hyperactivity disorder: A comparative study of two subtypes. Psychiatry Res. 1998, 81, 19–29. [Google Scholar] [CrossRef]
- Clarke, A.R.; Barry, R.J.; McCarthy, R.; Selikowitz, M. Age and sex effects in the EEG: Differences in two subtypes of attention-deficit/hyperactivity disorder. Clin. Neurophysiol. 2001, 112, 815–826. [Google Scholar] [CrossRef]
- Clarke, A.R.; Barry, R.J.; McCarthy, R.; Selikowitz, M. Age and sex effects in the EEG: Development of the normal child. Clin. Neurophysiol. 2001, 112, 806–814. [Google Scholar] [CrossRef]
- Bresnahan, S.M.; Anderson, J.W.; Barry, R.J. Age-related changes in quantitative EEG in attention-deficit/hyperactivity disorder. Biol. Psychiatry 1999, 46, 1690–1697. [Google Scholar] [CrossRef]
- Bresnahan, S.M.; Barry, R.J. Specificity of quantitative EEG analysis in adults with attention deficit hyperactivity disorder. Psychiatry Res. 2002, 112, 133–144. [Google Scholar] [CrossRef]
- Koehler, S.; Lauer, P.; Schreppel, T.; Jacob, C.; Heine, M.; Boreatti-Hümmer, A.; Fallgatter, A.; Herrmann, M. Increased EEG power density in alpha and theta bands in adult ADHD patients. J. Neural Transm. 2009, 116, 97–104. [Google Scholar] [CrossRef]
- Kamida, A.; Shimabayashi, K.; Oguri, M.; Takamori, T.; Ueda, N.; Koyanagi, Y.; Sannomiya, N.; Nagira, H.; Ikunishi, S.; Hattori, Y. EEG power spectrum analysis in children with ADHD. Yonago Acta Med. 2016, 59, 169. [Google Scholar]
- Shephard, E.; Tye, C.; Ashwood, K.L.; Azadi, B.; Asherson, P.; Bolton, P.F.; McLoughlin, G. Resting-state neurophysiological activity patterns in young people with ASD, ADHD, and ASD+ ADHD. J. Autism Dev. Disord. 2018, 48, 110–122. [Google Scholar] [CrossRef]
- Greber, M.; Klein, C.; Leipold, S.; Sele, S.; Jäncke, L. Heterogeneity of EEG resting-state brain networks in absolute pitch. Int. J. Psychophysiol. 2020, 157, 11–22. [Google Scholar] [CrossRef]
- Murias, M.; Swanson, J.M.; Srinivasan, R. Functional connectivity of frontal cortex in healthy and ADHD children reflected in EEG coherence. Cereb. Cortex 2007, 17, 1788–1799. [Google Scholar] [CrossRef] [Green Version]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef]
- Kahana, M.J.; Seelig, D.; Madsen, J.R. Theta returns. Curr. Opin. Neurobiol. 2001, 11, 739–744. [Google Scholar] [CrossRef]
- Michels, L.; Bucher, K.; Lüchinger, R.; Klaver, P.; Martin, E.; Jeanmonod, D.; Brandeis, D. Simultaneous EEG-fMRI during a working memory task: Modulations in low and high frequency bands. PLoS ONE 2010, 5, e10298. [Google Scholar] [CrossRef] [PubMed]
- Başar, E.; Schürmann, M.; Sakowitz, O. The selectively distributed theta system: Functions. Int. J. Psychophysiol. 2001, 39, 197–212. [Google Scholar] [CrossRef]
- Dietl, T.; Dirlich, G.; Vogl, L.; Lechner, C.; Strian, F. Orienting response and frontal midline theta activity: A somatosensory spectral perturbation study. Clin. Neurophysiol. 1999, 110, 1204–1209. [Google Scholar] [CrossRef]
- Clarke, A.R.; Barry, R.J.; Heaven, P.C.; McCarthy, R.; Selikowitz, M.; Byrne, M.K. EEG in adults with attention-deficit/hyperactivity disorder. Int. J. Psychophysiol. 2008, 70, 176–183. [Google Scholar] [CrossRef]
- Kitsune, G.L.; Cheung, C.H.; Brandeis, D.; Banaschewski, T.; Asherson, P.; McLoughlin, G.; Kuntsi, J. A matter of time: The influence of recording context on EEG spectral power in adolescents and young adults with ADHD. Brain Topogr. 2015, 28, 580–590. [Google Scholar] [CrossRef] [Green Version]
- Saha, P.; Mukhopdhyay, P.; Chakraborty, P.; Poria, S.; Mukundan, C.; Sharma, S.; Ghosh, P.; Vijay, M.; Nath, S.; Ghosh, S. Neural oscillations in resting state EEG in ADHD children-A preliminary study. J. Indian Assoc. Child Adolesc. Ment. Health 2017, 13, 180–207. [Google Scholar]
- Montagu, J. The hyperkinetic child: A behavioural, electrodermal and EEG investigation. Dev. Med. Child Neurol. 1975, 17, 299–305. [Google Scholar] [CrossRef]
- Chabot, R.J.; Serfontein, G. Quantitative electroencephalographic profiles of children with attention deficit disorder. Biol. Psychiatry 1996, 40, 951–963. [Google Scholar] [CrossRef]
- Romei, V.; Brodbeck, V.; Michel, C.; Amedi, A.; Pascual-Leone, A.; Thut, G. Spontaneous fluctuations in posterior α-band EEG activity reflect variability in excitability of human visual areas. Cereb. Cortex 2007, 18, 2010–2018. [Google Scholar] [CrossRef]
- Haegens, S.; Händel, B.F.; Jensen, O. Top-down controlled alpha band activity in somatosensory areas determines behavioral performance in a discrimination task. J. Neurosci. 2011, 31, 5197–5204. [Google Scholar] [CrossRef]
- Mathewson, K.E.; Lleras, A.; Beck, D.M.; Fabiani, M.; Ro, T.; Gratton, G. Pulsed out of awareness: EEG alpha oscillations represent a pulsed-inhibition of ongoing cortical processing. Front. Psychol. 2011, 2, 99. [Google Scholar] [CrossRef] [Green Version]
- Klimesch, W.; Sauseng, P.; Hanslmayr, S. EEG alpha oscillations: The inhibition–timing hypothesis. Brain Res. Rev. 2007, 53, 63–88. [Google Scholar] [CrossRef]
- Klimesch, W. Alpha-band oscillations, attention, and controlled access to stored information. Trends Cogn. Sci. 2012, 16, 606–617. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.E.; McEvoy, L.K.; Gevins, A. Neurophysiological indices of strategy development and skill acquisition. Cogn. Brain Res. 1999, 7, 389–404. [Google Scholar] [CrossRef]
- Lazzaro, I.; Gordon, E.; Whitmont, S.; Plahn, M.; Li, W.; Clarke, S.; Dosen, A.; Meares, R. Quantified EEG activity in adolescent attention deficit hyperactivity disorder. Clin. Electroencephalogr. 1998, 29, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Loo, S.K.; Hale, T.S.; Macion, J.; Hanada, G.; McGough, J.J.; McCracken, J.T.; Smalley, S.L. Cortical activity patterns in ADHD during arousal, activation and sustained attention. Neuropsychologia 2009, 47, 2114–2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loo, S.K.; Makeig, S. Clinical utility of EEG in attention-deficit/hyperactivity disorder: A research update. Neurotherapeutics 2012, 9, 569–587. [Google Scholar] [CrossRef] [Green Version]
- Woltering, S.; Jung, J.; Liu, Z.; Tannock, R. Resting state EEG oscillatory power differences in ADHD college students and their peers. Behav. Brain Funct. 2012, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Deiber, M.-P.; Hasler, R.; Colin, J.; Dayer, A.; Aubry, J.-M.; Baggio, S.; Perroud, N.; Ros, T. Linking alpha oscillations, attention and inhibitory control in adult ADHD with EEG neurofeedback. Neuroimage Clin. 2019, 25, 102145. [Google Scholar] [CrossRef]
- Loo, S.K.; Hale, T.S.; Hanada, G.; Macion, J.; Shrestha, A.; McGough, J.J.; McCracken, J.T.; Nelson, S.; Smalley, S.L. Familial clustering and DRD4 effects on electroencephalogram measures in multiplex families with attention deficit/hyperactivity disorder. J. Am. Acad. Child Adolesc. Psychiatry 2010, 49, 368–377. [Google Scholar] [PubMed]
- Clarke, A.R.; Barry, R.J.; Dupuy, F.E.; Heckel, L.D.; McCarthy, R.; Selikowitz, M.; Johnstone, S.J. Behavioural differences between EEG-defined subgroups of children with attention-deficit/hyperactivity disorder. Clin. Neurophysiol. 2011, 122, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Poil, S.-S.; Bollmann, S.; Ghisleni, C.; O’Gorman, R.; Klaver, P.; Ball, J.; Eich-Höchli, D.; Brandeis, D.; Michels, L. Age dependent electroencephalographic changes in attention-deficit/hyperactivity disorder (ADHD). Clin. Neurophysiol. 2014, 125, 1626–1638. [Google Scholar] [CrossRef] [PubMed]
- Hale, T.S.; Smalley, S.L.; Dang, J.; Hanada, G.; Macion, J.; McCracken, J.T.; McGough, J.J.; Loo, S.K. ADHD familial loading and abnormal EEG alpha asymmetry in children with ADHD. J. Psychiatr. Res. 2010, 44, 605–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenartowicz, A.; Mazaheri, A.; Jensen, O.; Loo, S.K. Aberrant modulation of brain oscillatory activity and attentional impairment in attention-deficit/hyperactivity disorder. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M.A.; Wallois, F.; Aarabi, A.; Berquin, P. Dynamic changes in quantitative electroencephalogram during continuous performance test in children with attention-deficit/hyperactivity disorder. Int. J. Psychophysiol. 2011, 81, 230–236. [Google Scholar] [CrossRef]
- Ackerman, P.T.; Dykman, R.A.; Oglesby, D.M.; Newton, J.E. EEG power spectra of children with dyslexia, slow learners, and normally reading children with ADD during verbal processing. J. Learn. Disabil. 1994, 27, 619–630. [Google Scholar] [CrossRef]
- Andreassi, J. Psychophysiology: Human Behavior and Physiological Response, 3rd ed.; L. Lawrence Eflbaum Assoc.: Mahwah, NJ, USA, 1995. [Google Scholar]
- Buyck, I.; Wiersema, J.R. Resting electroencephalogram in attention deficit hyperactivity disorder: Developmental course and diagnostic value. Psychiatry Res. 2014, 216, 391–397. [Google Scholar] [CrossRef]
- Tye, C.; McLoughlin, G.; Kuntsi, J.; Asherson, P. Electrophysiological markers of genetic risk for attention deficit hyperactivity disorder. Expert Rev. Mol. Med. 2011, 13, e9. [Google Scholar] [CrossRef]
- Clarke, A.R.; Barry, R.J.; McCarthy, R.; Selikowitz, M. Excess beta activity in children with attention-deficit/hyperactivity disorder: An atypical electrophysiological group. Psychiatry Res. 2001, 103, 205–218. [Google Scholar] [CrossRef]
- Clarke, A.R.; Barry, R.J.; McCarthy, R.; Selikowitz, M. EEG-defined subtypes of children with attention-deficit/hyperactivity disorder. Clin. Neurophysiol. 2001, 112, 2098–2105. [Google Scholar] [CrossRef]
- Sidlauskaite, J.; Sonuga-Barke, E.; Roeyers, H.; Wiersema, J.R. Altered intrinsic organisation of brain networks implicated in attentional processes in adult attention-deficit/hyperactivity disorder: A resting-state study of attention, default mode and salience network connectivity. Eur. Arch. Psychiatry Clin. Neurosci. 2016, 266, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Michelini, G.; Jurgiel, J.; Bakolis, I.; Cheung, C.H.; Asherson, P.; Loo, S.K.; Kuntsi, J.; Mohammad-Rezazadeh, I. Atypical functional connectivity in adolescents and adults with persistent and remitted ADHD. bioRxiv 2018, 1, 201772. [Google Scholar] [CrossRef] [Green Version]
- Stam, C.J.; Nolte, G.; Daffertshofer, A. Phase lag index: Assessment of functional connectivity from multi channel EEG and MEG with diminished bias from common sources. Hum. Brain Mapp. 2007, 28, 1178–1193. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Marqui, R.D.; Michel, C.M.; Lehmann, D. Low resolution electromagnetic tomography: A new method for localizing electrical activity in the brain. Int. J. Psychophysiol. 1994, 18, 49–65. [Google Scholar] [CrossRef]
- Schoffelen, J.M.; Gross, J. Source connectivity analysis with MEG and EEG. Hum. Brain Mapp. 2009, 30, 1857–1865. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Marqui, R.D.; Lehmann, D.; Koukkou, M.; Kochi, K.; Anderer, P.; Saletu, B.; Tanaka, H.; Hirata, K.; John, E.R.; Prichep, L. Assessing interactions in the brain with exact low-resolution electromagnetic tomography. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 3768–3784. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmadi, M.; Kazemi, K.; Kuc, K.; Cybulska-Klosowicz, A.; Helfroush, M.S.; Aarabi, A. Disrupted Functional Rich-Club Organization of the Brain Networks in Children with Attention-Deficit/Hyperactivity Disorder, a Resting-State EEG Study. Brain Sci. 2021, 11, 938. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070938
Ahmadi M, Kazemi K, Kuc K, Cybulska-Klosowicz A, Helfroush MS, Aarabi A. Disrupted Functional Rich-Club Organization of the Brain Networks in Children with Attention-Deficit/Hyperactivity Disorder, a Resting-State EEG Study. Brain Sciences. 2021; 11(7):938. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070938
Chicago/Turabian StyleAhmadi, Maliheh, Kamran Kazemi, Katarzyna Kuc, Anita Cybulska-Klosowicz, Mohammad Sadegh Helfroush, and Ardalan Aarabi. 2021. "Disrupted Functional Rich-Club Organization of the Brain Networks in Children with Attention-Deficit/Hyperactivity Disorder, a Resting-State EEG Study" Brain Sciences 11, no. 7: 938. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070938