
Enhanced Expectation of External Sensations of the Chest Regulates the Emotional Perception of Fearful Faces

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Participants
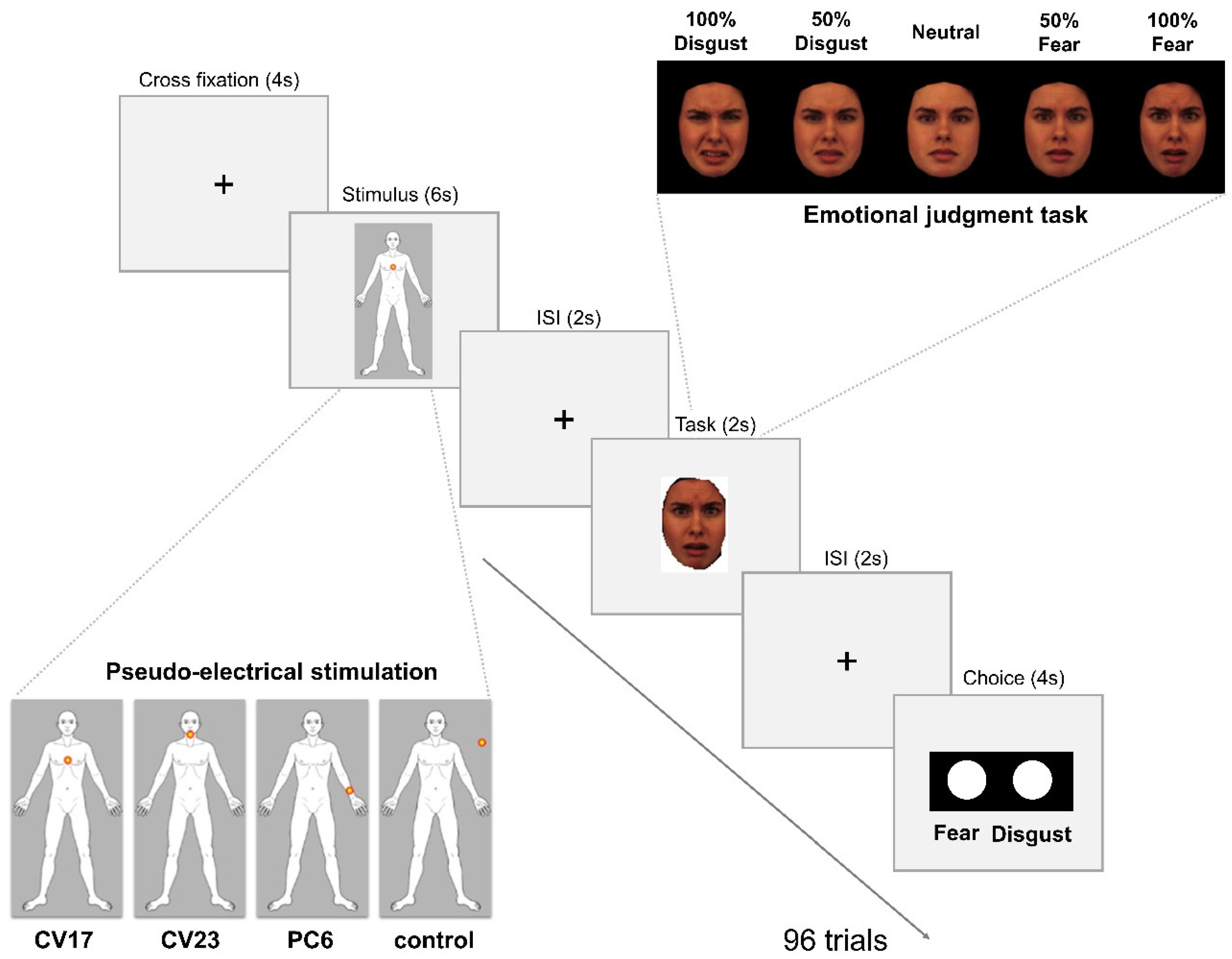
2.2. Pseudo-Electrical Acupuncture Stimulation
2.3. Emotional Face Stimuli and Emotion Judgment Task
2.4. Behavioral Analysis
2.5. fMRI Acquisition and Analysis
3. Results
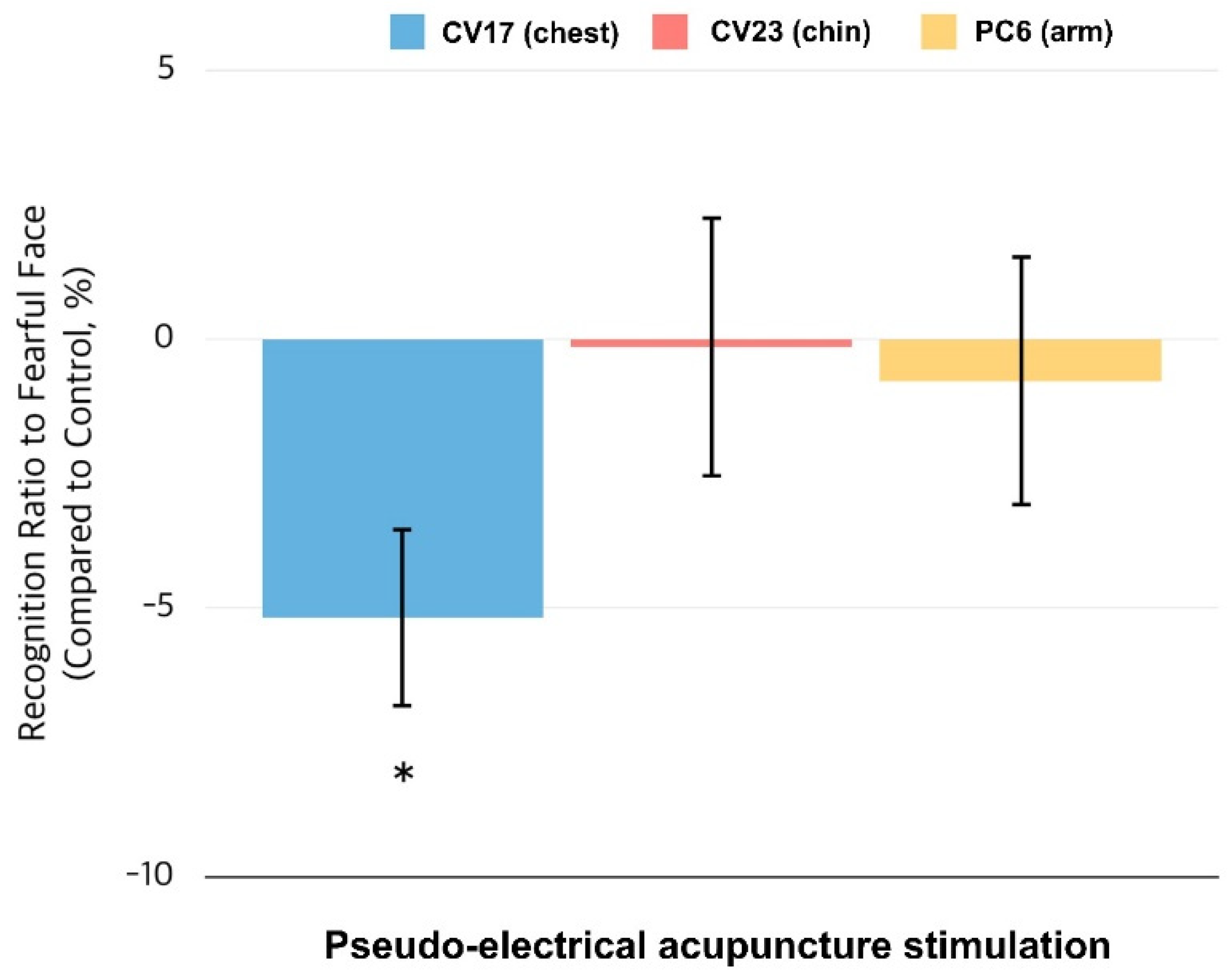
3.1. Behavioral Responses to the Emotion Judgment Task
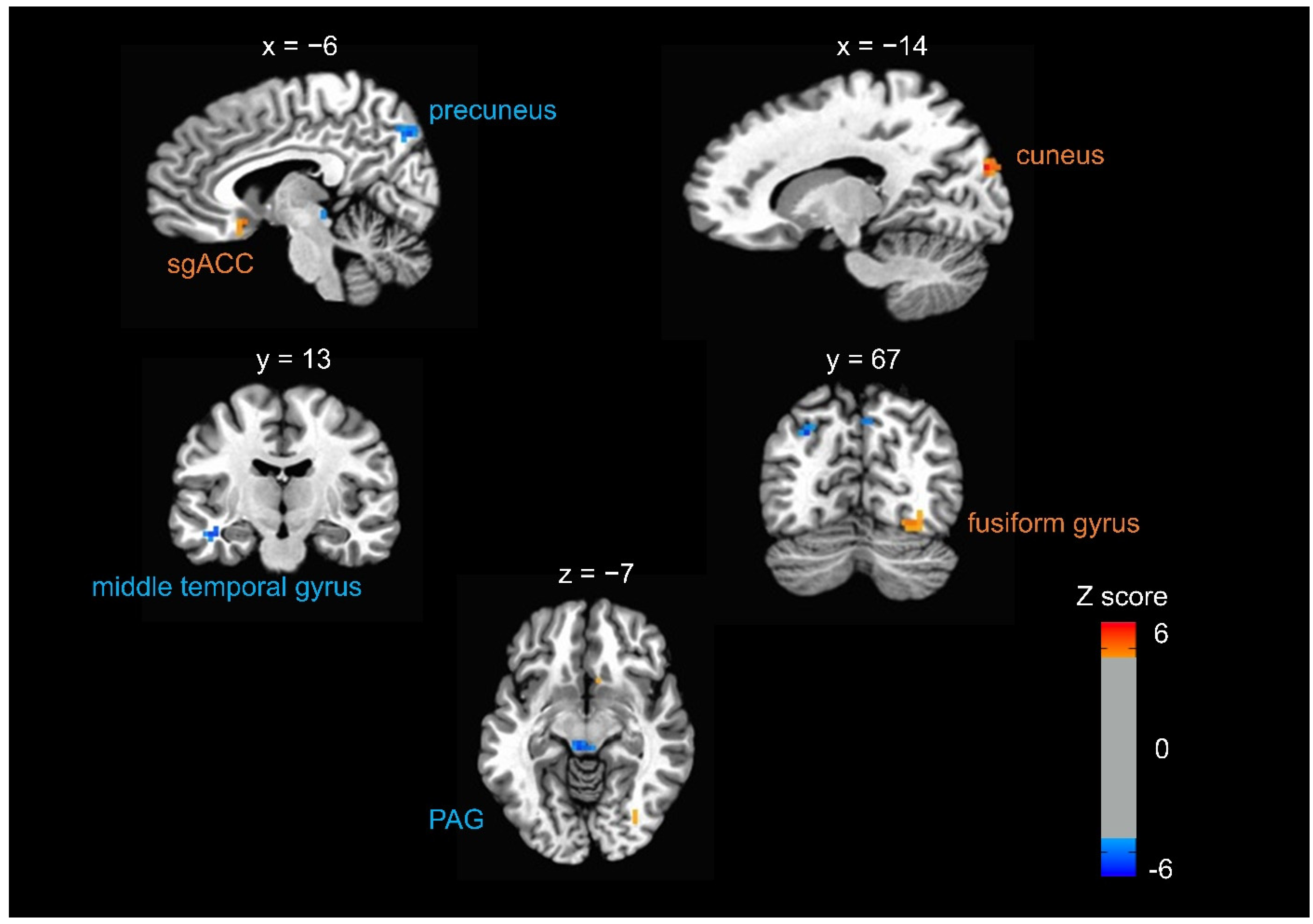
3.2. Brain Responses to Fearful Faces When Expecting Somatosensory Stimulation at CV17
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bechara, A.; Damasio, H.; Damasio, A.R. Emotion, decision making and the orbitofrontal cortex. Cereb. Cortex. 2000, 10, 295–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Critchley, H.D. Psychophysiology of neural, cognitive and affective integration: fMRI and autonomic indicants. Int. J. Psychophysiol. 2009, 73, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Critchley, H.D.; Nagai, Y.; Gray, M.A.; Mathias, C.J. Dissecting axes of autonomic control in humans: Insights from neuroimaging. Auton. Neurosci. 2011, 161, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Critchley, H.D.; Wiens, S.; Rotshtein, P.; Öhman, V.; Dolan, R.J. Neural systems supporting interoceptive awareness. Nat. Neurosci. 2004, 7, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, M.A.; Rylander, K.; Harrison, N.A.; Wallin, B.G.; Critchley, H.D. Following one’s heart: Cardiac rhythms gate central initiation of sympathetic reflexes. J. Neurosci. 2009, 29, 1817–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, N.A.; Brydon, L.; Walker, C.; Gray, M.A.; Steptoe, A.; Critchley, H.D. Inflammation causes mood changes through alterations in subgenual cingulate activity and mesolimbic connectivity. Biol. Psychiatry 2009, 66, 407–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, N.A.; Brydon, L.; Walker, C.; Gray, M.A.; Steptoe, A.; Dolan, R.J.; Critchley, H.D. Neural origins of human sickness in interoceptive responses to inflammation. Biol. Psychiatry 2009, 66, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, N.A.; Gray, M.A.; Gianaros, P.J.; Critchley, H.D. The embodiment of emotional feelings in the brain. J. Neurosci. 2010, 30, 12878–12884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Critchley, H.D.; Garfinkel, S.N. Interoception and emotion. Curr. Opin. Psychol. 2017, 17, 7–14. [Google Scholar] [CrossRef]
- James, W. The physical basis of emotion. Psychol. Rev. 1994, 101, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Aminoff, E.M.; Kveraga, K.; Bar, M. The role of the parahippocampal cortex in cognition. Trends Cogn. Sci. 2013, 17, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Barrett, L.F.; Quigley, K.S.; Bliss-Moreau, E. Interoceptive sensitivity and self-reports of emotional experience. J. Pers. Soc. Psychol. 2004, 87, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasawa, Y.; Moriguchi, Y.; Tochizawa, S.; Umeda, S. Interoceptive sensitivity predicts sensitivity to the emotions of others. Cogn. Emot. 2014, 28, 1435–1448. [Google Scholar] [CrossRef] [PubMed]
- Wiens, S. Interoception in emotional experience. Curr. Opin. Neurol. 2005, 18, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Muller, L.E.; Schulz, A.; Andermann, M. Cortical Representation of Afferent Bodily Signals in Borderline Personality Disorder: Neural Correlates and Relationship to Emotional Dysregulation. JAMA Psychiatry 2015, 72, 1077–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garfinkel, S.N.; Critchley, H.D. Threat and the Body: How the Heart Supports Fear Processing. Trends Cogn. Sci. 2016, 20, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Garfinkel, S.N.; Minati, L.; Gray, M.A.; Seth, A.K.; Dolan, R.J.; Critchley, H.D. Fear from the heart: Sensitivity to fear stimuli depends on individual heartbeats. J. Neurosci. 2014, 34, 6573–6582. [Google Scholar] [CrossRef] [PubMed]
- Khalsa, S.S.; Rudrauf, D.; Feinstein, J.S. The pathways of interoceptive awareness. Nat. Neurosci. 2009, 12, 1494–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paton, J.F.; Li, Y.W.; Kasparov, S. Reflex response and convergence of pharyngoesophageal and peripheral chemoreceptors in the nucleus of the solitary tract. Neuroscience 1999, 93, 143–154. [Google Scholar] [CrossRef]
- Craig, A.D. Interoception: The sense of the physiological condition of the body. Curr. Opin. Neurobiol. 2003, 13, 500–505. [Google Scholar] [CrossRef]
- Singer, T.; Critchley, H.D.; Preuschoff, K. A common role of insula in feelings, empathy and uncertainty. Trends Cogn. Sci. 2009, 13, 334–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hietanen, J.K.; Glerean, E.; Hari, R.; Nummenmaa, L. Bodily maps of emotions across child development. Dev. Sci. 2016, 19, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.M.; Ryu, Y.; Lee, Y.S.; Wallraven, C.; Chae, Y. Role of interoceptive accuracy in topographical changes in emotion-induced bodily sensations. PLoS ONE 2017, 12, e0183211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nummenmaa, L.; Glerean, E.; Hari, R.; Hietanen, J.K. Bodily maps of emotions. Proc. Natl. Acad. Sci. USA 2014, 111, 646–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, W.M.; Lee, Y.S.; Lee, I.S.; Wallraven, C.; Ryu, Y.; Chae, Y. Enhanced bodily states of fear facilitates bias perception of fearful faces. Mol. Brain 2020, 13, 157. [Google Scholar] [CrossRef]
- Couto, B.; Salles, A.; Sedeno, L.; Peradejordi, M.; Barttfeld, P.; Canales-Johnson, A.; Dos Santos, Y.V.; Huepe, D.; Bekinschtein, T.; Sigman, M.; et al. The man who feels two hearts: The different pathways of interoception. Soc. Cogn. Affect Neurosci. 2014, 9, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Aspell, J.E.; Heydrich, L.; Marillier, G. Turning body and self inside out: Visualized heartbeats alter bodily self-consciousness and tactile perception. Psychol. Sci. 2013, 24, 2445–2453. [Google Scholar] [CrossRef] [PubMed]
- Schauder, K.B.; Mash, L.E.; Bryant, L.K.; Cascio, C.J. Interoceptive ability and body awareness in autism spectrum disorder. J. Exp. Child Psychol. 2015, 131, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Garfinkel, S.N.; Critchley, H.D.; Seth, A.K. Multisensory integration across exteroceptive and interoceptive domains modulates self-experience in the rubber-hand illusion. Neuropsychologia 2013, 51, 2909–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsakiris, M.; Tajadura-Jimenez, A.; Costantini, M. Just a heartbeat away from one’s body: Interoceptive sensitivity predicts malleability of body-representations. Proc. Biol. Sci. 2011, 278, 2470–2476. [Google Scholar] [CrossRef] [Green Version]
- Seth, A.K. Interoceptive inference, emotion, and the embodied self. Trends Cogn. Sci. 2013, 17, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Barrett, L.F. The theory of constructed emotion: An active inference account of interoception and categorization. Soc. Cogn. Affect. Neurosci. 2017, 12, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.; Kilner, J.; Harrison, L. A free energy principle for the brain. J. Physiol. Paris 2006, 100, 70–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.S.; Mumford, D. Hierarchical Bayesian inference in the visual cortex. J. Opt. Soc. Am. A Opt. Image. Sci. Vis. 2003, 20, 1434–1448. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.M.; Ryu, Y.; Park, H.J.; Lee, H.; Chae, Y. Brain activation during the expectations of sensory experience for cutaneous electrical stimulation. Neuroimage Clin. 2018, 19, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Isoyama, D.; Cordts, E.B.; van Niewegen, A.M.B.d.S. Effect of acupuncture on symptoms of anxiety in women undergoing in vitro fertilisation: A prospective randomised controlled study. Acupunct. Med. 2012, 30, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Kurono, Y.; Minagawa, M.; Ishigami, T. Acupuncture to Danzhong but not to Zhongting increases the cardiac vagal component of heart rate variability. Auton. Neurosci. 2011, 161, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Park, H.J.; Han, S.M.; Hahm, D.H.; Lee, H.J.; Kim, K.S.; Shim, I. The effects of acupuncture stimulation at PC6 (Neiguan) on chronic mild stress-induced biochemical and behavioral responses. Neurosci. Lett. 2009, 460, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Park, H.J.; Shim, H.S.; Han, S.M.; Hahm, D.H.; Lee, H.; Shim, I. The effects of acupuncture (PC6) on chronic mild stress-induced memory loss. Neurosci. Lett. 2011, 488, 225–288. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.L.; Zhao, G.W.; Li, H.Z.; Yang, X.D.; Wu, Y.Y.; Lin, F.; Guan, L.X.; Zhai, F.G.; Liu, J.Q.; Yang, C.H.; et al. Acupuncture Attenuates Anxiety-Like Behavior by Normalizing Amygdaloid Catecholamines during Ethanol Withdrawal in Rats. Evid. Based Complement. Altern. Med. 2011, 2011, 429843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beissner, F. Therapeutic Sensations: A New Unifying Concept. Evid. Based Complement. Altern. Med. 2020, 2020, 7630190. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.M.; Shim, W.; Lee, T.; Park, H.J.; Ryu, Y.; Beissner, F.; Chae, Y. More than DeQi: Spatial Patterns of Acupuncture-Induced Bodily Sensations. Front. Neurosci. 2016, 10, 462. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.W. AFNI: Software for analysis and visualization of functional magnetic resonance neuroimages. Comput. Biomed. Res. 1996, 29, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Paus, T.; Otaky, N.; Caramanos, Z. In vivo morphometry of the intrasulcal gray matter in the human cingulate, paracingulate, and superior-rostral sulci: Hemispheric asymmetries, gender differences and probability maps. J. Comp. Neurol. 1996, 376, 664–673. [Google Scholar] [CrossRef]
- Forman, S.D.; Cohen, J.D.; Fitzgerald, M.; Eddy, W.F.; Mintun, M.A.; Noll, D.C. Improved assessment of significant activation in functional magnetic resonance imaging (fMRI): Use of a cluster-size threshold. Magn. Reson. Med. 1995, 33, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Eccles, J.A.; Garfinkel, S.N.; Harrison, N.A.; Ward, J.; Taylor, R.E.; Bewley, A.P.; Critchley, H.D. Sensations of skin infestation linked to abnormal frontolimbic brain reactivity and differences in self-representation. Neuropsychologia 2015, 77, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Behbehani, M.M. Functional characteristics of the midbrain periaqueductal gray. Prog. Neurobiol. 1995, 46, 575–605. [Google Scholar] [CrossRef]
- Carrive, P.; Bandler, R.; Dampney, R.A. Viscerotopic control of regional vascular beds by discrete groups of neurons within the midbrain periaqueductal gray. Brain Res. 1989, 493, 385–390. [Google Scholar] [CrossRef]
- Mobbs, D.; Petrovic, P.; Marchant, J.L.; Hassabis, D.; Weiskopf, N. When fear is near: Threat imminence elicits prefrontal-periaqueductal gray shifts in humans. Science 2007, 317, 1079–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelena, D.; Menant, O.; Andersson, F.; Chaillou, E. Periaqueductal gray and emotions: The complexity of the problem and the light at the end of the tunnel, the magnetic resonance imaging. Endocr. Regul. 2018, 52, 222–238. [Google Scholar] [CrossRef] [Green Version]
- Terpou, B.A.; Harricharan, S.; McKinnon, M.C.; Rakesh, J.P.F.; Lanius, R.A. The effects of trauma on brain and body: A unifying role for the midbrain periaqueductal gray. J. Neurosci. Res. 2019, 97, 1110–1140. [Google Scholar] [CrossRef] [PubMed]
- Harricharan, S.; Rabellino, D.; Frewen, P.A.; Densmore, M.; Theberge, J.; Mckinnon, M.C.; Schore, R.; Lanius, A. fMRI functional connectivity of the periaqueductal gray in PTSD and its dissociative subtype. Brain Behav. 2016, 6, e00579. [Google Scholar] [CrossRef] [PubMed]
- Weng, H.Y.; Feldman, J.L.; Leggio, L.; Napadow, V.; Park, J.; Price, C. Interventions and Manipulations of Interoception. Trends Neurosci. 2021, 44, 52–62. [Google Scholar] [CrossRef]
- Fox, K.C.; Dixon, M.L.; Nijeboer, S.; Girn, M.; Floman, J.L.; Lifshitz, M.; Ellamil, M.; Sedlmeier, P.; Christoff, K. Functional neuroanatomy of meditation: A review and meta-analysis of 78 functional neuroimaging investigations. Neurosci. Biobehav. Rev. 2016, 65, 208–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, A.; Slagter, H.A.; Dunne, J.D.; Davidson, R. Attention regulation and monitoring in meditation. Trends Cogn. Sci. 2008, 12, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleckner, I.R.; Zhang, J.; Touroutoglou, A.; Chanes, L.; Xia, C.; Simmons, W.K.; Quigley, K.S.; Dickerson, B.C.; Barrett, L.F. Evidence for a Large-Scale Brain System Supporting Allostasis and Interoception in Humans. Nat. Hum. Behav. 2017, 1, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbruzzese, L.; Magnani, N.; Robertson, I.H.; Mancuso, M. Age and Gender Differences in Emotion Recognition. Front. Psychol. 2019, 10, 2371. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Zhang, S.; Le, T.M.; Tang, X.; Li, C.S.R. Neural responses to negative facial emotions: Sex differences in the correlates of individual anger and fear traits. Neuroimage 2020, 221, 117–171. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Activation | Location | Z Score | Voxels | Coordinates of Peak Voxel in Talairach Space (RAI) | |||
|---|---|---|---|---|---|---|---|
| X | Y | Z | |||||
| Decreased activation | Precuneus | Left | −5.10 | 26 | 28.5 | 67.5 | 35.5 |
| Periaqueductal grey | Right /Left | −5.09 | 20 | 4.5 | 28.5 | −6.5 | |
| Precuneus | Right | −4.93 | 17 | −4.5 | 73.5 | 38.5 | |
| Precuneus | Left | −5.72 | 16 | 1.5 | 61.5 | 41.5 | |
| Middle temporal gyrus | Left | −5.16 | 14 | 40.5 | 13.5 | −15.5 | |
| Increased activation | Cuneus | Right | 5.57 | 18 | −13.5 | 85.5 | 23.5 |
| Fusiform gyrus | Right | 4.49 | 16 | −25.5 | 67.5 | −12.5 | |
| Subgenual anterior cingulate cortex | Right | 4.5 | 10 | −4.5 | −13.5 | −9.5 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, W.-M.; Lee, I.-S.; Lee, Y.-S.; Ryu, Y.; Park, H.-J.; Chae, Y. Enhanced Expectation of External Sensations of the Chest Regulates the Emotional Perception of Fearful Faces. Brain Sci. 2021, 11, 946. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070946
Jung W-M, Lee I-S, Lee Y-S, Ryu Y, Park H-J, Chae Y. Enhanced Expectation of External Sensations of the Chest Regulates the Emotional Perception of Fearful Faces. Brain Sciences. 2021; 11(7):946. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070946
Chicago/Turabian StyleJung, Won-Mo, In-Seon Lee, Ye-Seul Lee, Yeonhee Ryu, Hi-Joon Park, and Younbyoung Chae. 2021. "Enhanced Expectation of External Sensations of the Chest Regulates the Emotional Perception of Fearful Faces" Brain Sciences 11, no. 7: 946. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070946