
Endogenous Expression of G-CSF in Rat Dorsal Root Ganglion Neurons after Nerve Injury


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. Animals
2.3. Establishment of the Neuropathic Pain Animal Model
2.4. Behavioral Testing
2.5. G-CSF Administration
2.6. Tissue Preparation and Sectioning
2.7. Immunohistochemistry
2.8. Double Immunofluorescence Staining
2.9. Image Analysis
2.10. Blood Sample Collection and Analysis
2.11. Statistical Analysis
3. Results
3.1. G-CSFR Is Expressed in Small- and Medium-Sized DRG Neurons
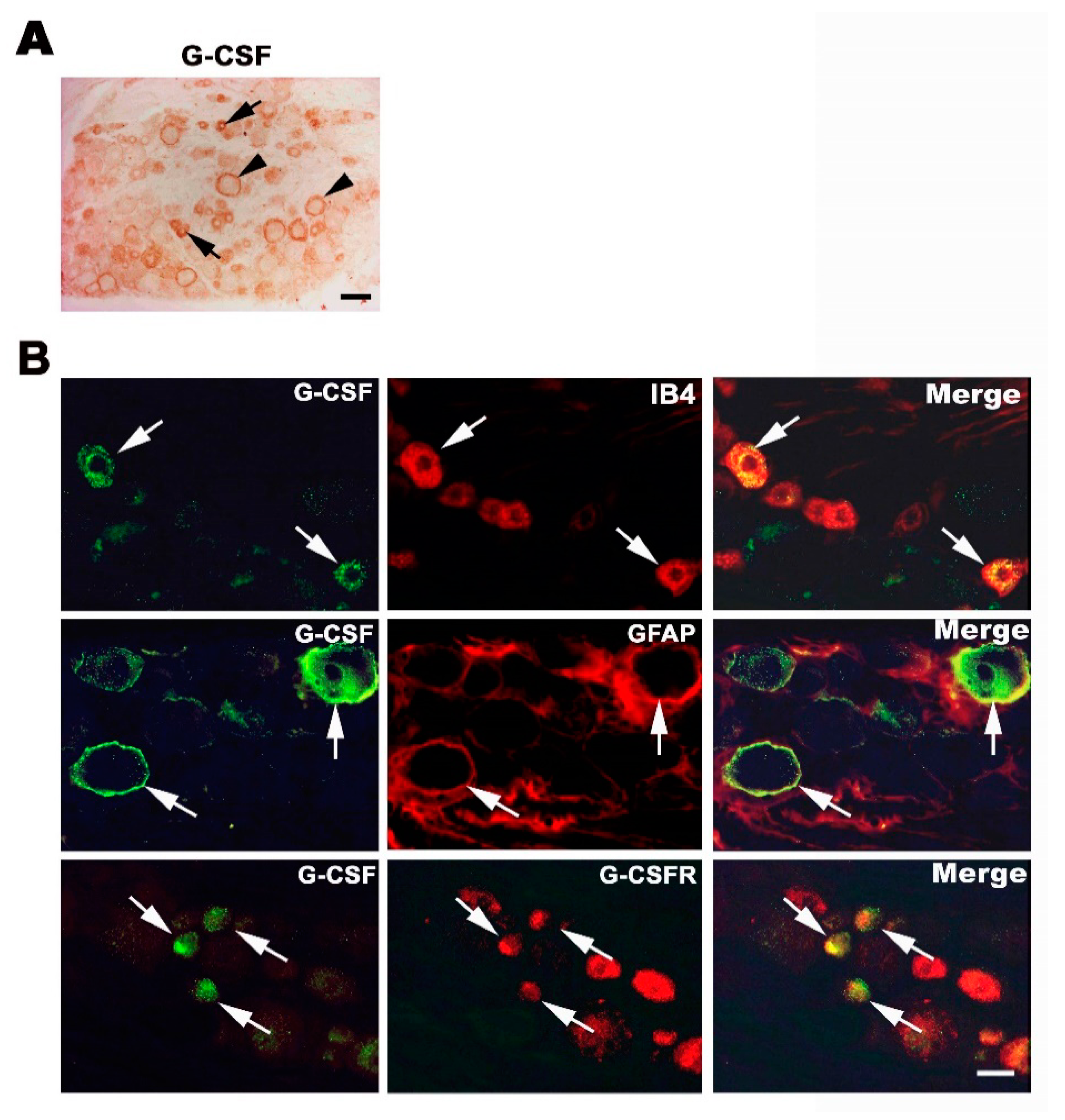
3.2. Endogenous G-CSF Is Expressed in Small and Medium-Size DRG Neurons and Satellite Cells
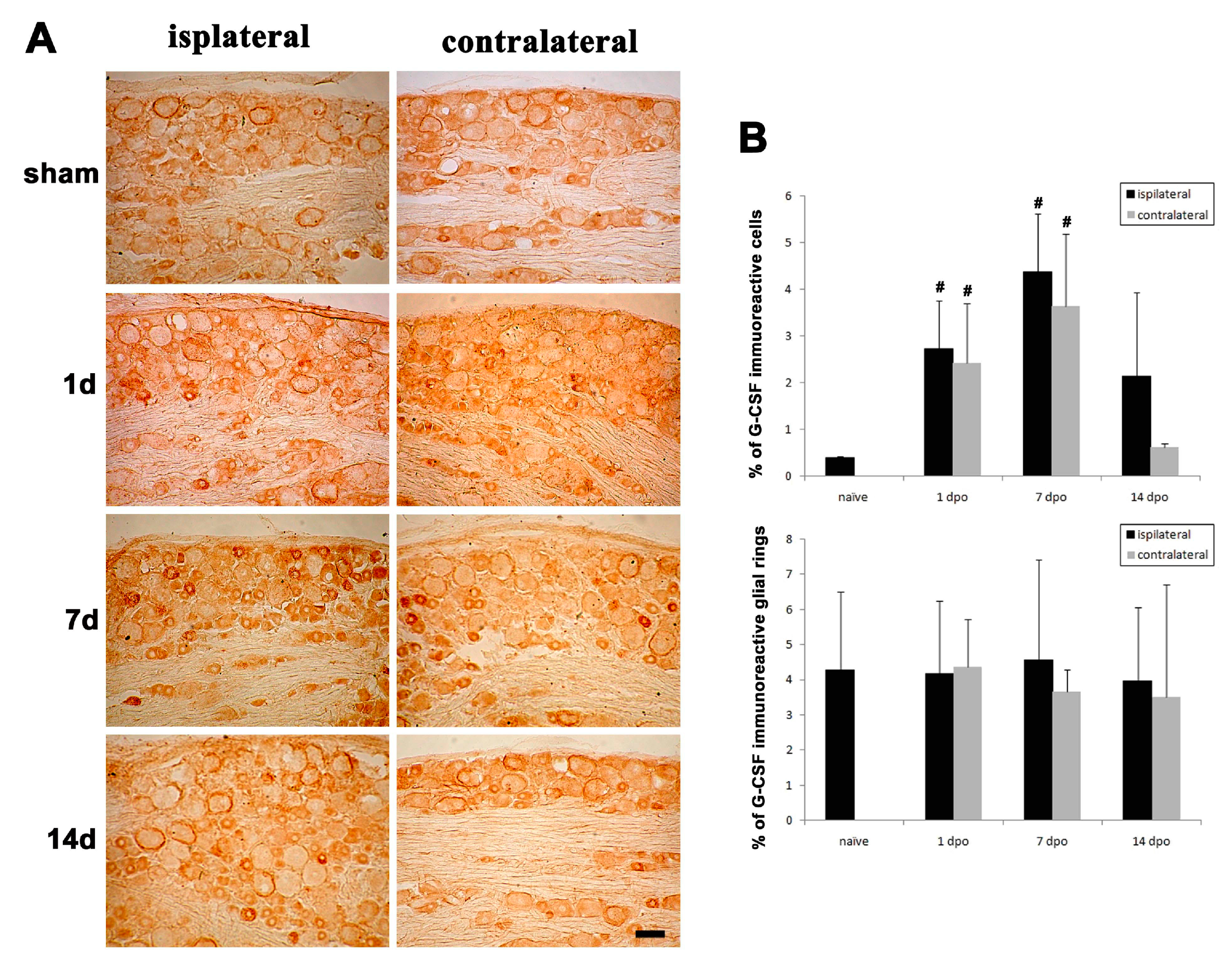
3.3. Changes in G-CSF Expression in the DRG in Rats with Partial Sciatic Nerve Transection (PST)
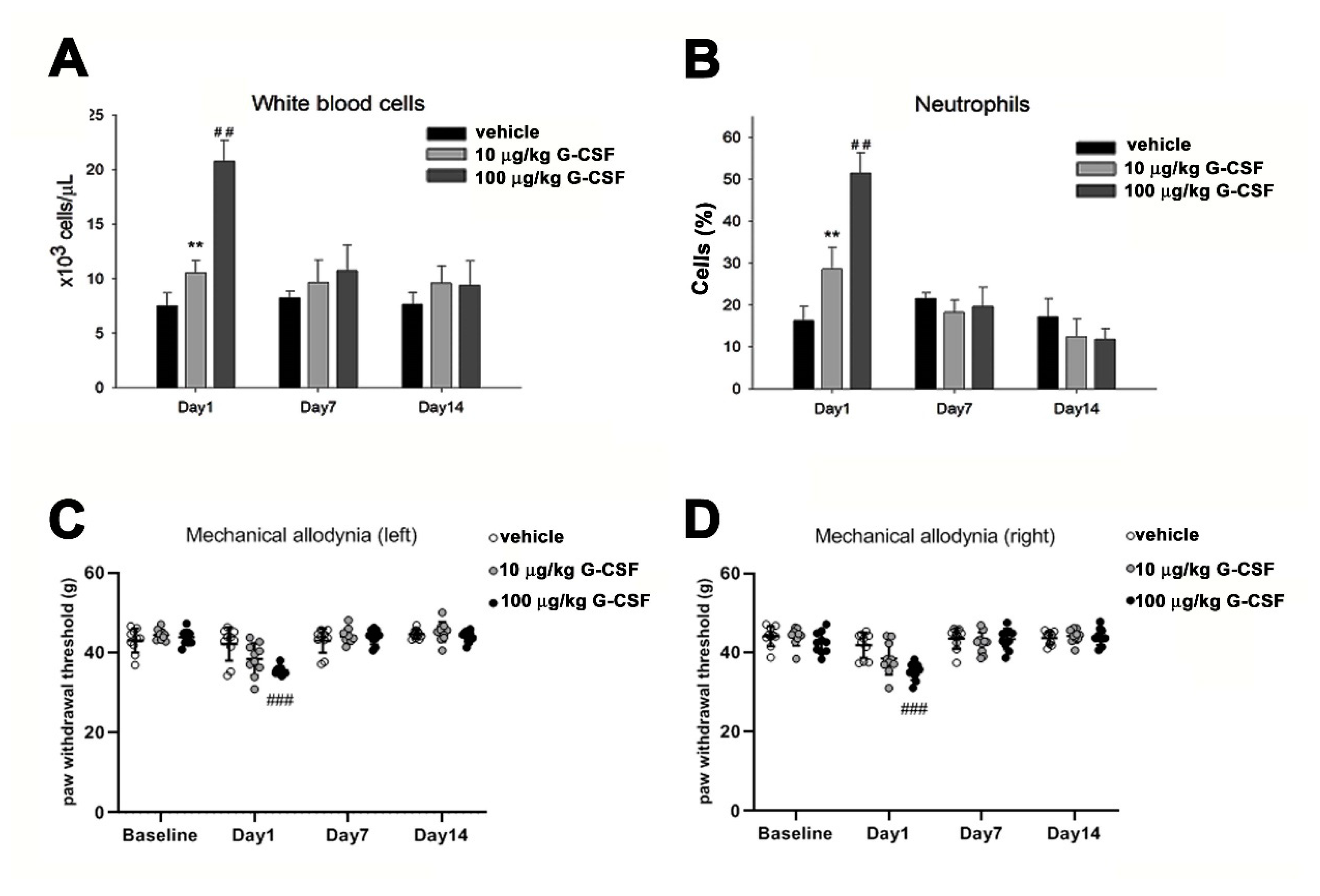
3.4. G-CSF Induces Pain Behavior
3.5. G-CSF Enhances Mechanical Allodynia Following PST
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grace, P.M.; Tawfik, V.L.; Svensson, C.I.; Burton, M.D.; Loggia, M.L.; Hutchinson, M.R. The Neuroimmunology of Chronic Pain: From Rodents to Humans. J. Neurosci. Off. J. Soc. Neurosci. 2021, 41, 855–865. [Google Scholar] [CrossRef]
- Yeh, T.Y.; Luo, I.W.; Hsieh, Y.L.; Tseng, T.J.; Chiang, H.; Hsieh, S.T. Peripheral Neuropathic Pain: From Experimental Models to Potential Therapeutic Targets in Dorsal Root Ganglion Neurons. Cells 2020, 9, 2725. [Google Scholar] [CrossRef] [PubMed]
- Szok, D.; Tajti, J.; Nyari, A.; Vecsei, L. Therapeutic Approaches for Peripheral and Central Neuropathic Pain. Behav. Neurol. 2019, 2019, 8685954. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, A.; Marsch, L.A.; Joseph, H.; Portenoy, R.K. Opioids and the treatment of chronic pain: Controversies, current status, and future directions. Exp. Clin. Psychopharmacol. 2008, 16, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Oelhaf, R.C.; Azadfard, M. Opioid Toxicity. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- O'Brien, J.B.; Roman, D.L. Novel treatments for chronic pain: Moving beyond opioids. Transl. Res. J. Lab. Clin. Med. 2021, 234, 1–19. [Google Scholar]
- Jahandideh, B.; Derakhshani, M.; Abbaszadeh, H.; Akbar Movassaghpour, A.; Mehdizadeh, A.; Talebi, M.; Yousefi, M. The pro-Inflammatory cytokines effects on mobilization, self-renewal and differentiation of hematopoietic stem cells. Hum. Immunol. 2020, 81, 206–217. [Google Scholar] [CrossRef]
- Dwivedi, P.; Greis, K.D. Granulocyte colony-stimulating factor receptor signaling in severe congenital neutropenia, chronic neutrophilic leukemia, and related malignancies. Exp. Hematol. 2017, 46, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.H.; Wang, T.F.; Yang, K.L. Hematopoietic stem cell donation. Int. J. Hematol. 2013, 97, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.C.; Hoffmann, B.W.; Csar, X.F.; Hamilton, J.A. Granulocyte colony-stimulating factor-stimulated proliferation of myeloid cells: Mode of cell cycle control by a range of inhibitors. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 1996, 16, 869–877. [Google Scholar] [CrossRef]
- Lejnieks, D.V.; Han, S.W.; Ramesh, N.; Lau, S.; Osborne, W.R. Granulocyte colony-stimulating factor expression from transduced vascular smooth muscle cells provides sustained neutrophil increases in rats. Hum. Gene Ther. 1996, 7, 1431–1436. [Google Scholar] [CrossRef]
- Leizer, T.; Cebon, J.; Layton, J.E.; Hamilton, J.A. Cytokine regulation of colony-stimulating factor production in cultured human synovial fibroblasts: I. Induction of GM-CSF and G-CSF production by interleukin-1 and tumor necrosis factor. Blood 1990, 76, 1989–1996. [Google Scholar] [CrossRef] [Green Version]
- Sieff, C.A.; Niemeyer, C.M.; Mentzer, S.J.; Faller, D.V. Interleukin-1, tumor necrosis factor, and the production of colony-stimulating factors by cultured mesenchymal cells. Blood 1988, 72, 1316–1323. [Google Scholar] [CrossRef]
- Horio, H.; Nomori, H.; Morinaga, S.; Kikuchi, T.; Tomonari, H.; Kuriyama, S.; Suemasu, K. Granulocyte colony-stimulating factor-producing primary pericardial mesothelioma. Hum. Pathol. 1999, 30, 718–720. [Google Scholar] [CrossRef]
- Shukla, A.; MacPherson, M.B.; Hillegass, J.; Ramos-Nino, M.E.; Alexeeva, V.; Vacek, P.M.; Bond, J.P.; Pass, H.I.; Steele, C.; Mossman, B.T. Alterations in gene expression in human mesothelial cells correlate with mineral pathogenicity. Am. J. Respir. Cell Mol. Biol. 2009, 41, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tweardy, D.J.; Mott, P.L.; Glazer, E.W. Monokine modulation of human astroglial cell production of granulocyte colony-stimulating factor and granulocyte-macrophage colony-stimulating factor. I. Effects of IL-1 alpha and IL-beta. J. Immunol. 1990, 144, 2233–2241. [Google Scholar] [PubMed]
- Rahi, V.; Jamwal, S.; Kumar, P. Neuroprotection through G-CSF: Recent advances and future viewpoints. Pharmacol. Rep. PR 2021. [Google Scholar] [CrossRef] [PubMed]
- Hasselblatt, M.; Jeibmann, A.; Riesmeier, B.; Maintz, D.; Schabitz, W.R. Granulocyte-colony stimulating factor (G-CSF) and G-CSF receptor expression in human ischemic stroke. Acta Neuropathol. 2007, 113, 45–51. [Google Scholar] [CrossRef]
- Taichman, R.S.; Emerson, S.G. Human osteosarcoma cell lines MG-63 and SaOS-2 produce G-CSF and GM-CSF: Identification and partial characterization of cell-associated isoforms. Exp. Hematol. 1996, 24, 509–517. [Google Scholar]
- Menzie-Suderam, J.M.; Modi, J.; Xu, H.; Bent, A.; Trujillo, P.; Medley, K.; Jimenez, E.; Shen, J.; Marshall, M.; Tao, R.; et al. Granulocyte-colony stimulating factor gene therapy as a novel therapeutics for stroke in a mouse model. J. Biomed. Sci. 2020, 27, 99. [Google Scholar] [CrossRef]
- Lu, F.; Nakamura, T.; Toyoshima, T.; Liu, Y.; Shinomiya, A.; Hirooka, K.; Okabe, N.; Miyamoto, O.; Tamiya, T.; Keep, R.F.; et al. Neuroprotection of granulocyte colony-stimulating factor during the acute phase of transient forebrain ischemia in gerbils. Brain Res. 2014, 1548, 49–55. [Google Scholar] [CrossRef]
- Schabitz, W.R.; Kollmar, R.; Schwaninger, M.; Juettler, E.; Bardutzky, J.; Scholzke, M.N.; Sommer, C.; Schwab, S. Neuroprotective effect of granulocyte colony-stimulating factor after focal cerebral ischemia. Stroke 2003, 34, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Liu, S.; Wang, P.; Zhang, H.; Wang, F.; Bing, L.; Gao, J.; Yang, J.; Hao, A. Granulocyte colony-stimulating factor improves neuron survival in experimental spinal cord injury by regulating nucleophosmin-1 expression. J. Neurosci. Res. 2014, 92, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Khorasanizadeh, M.; Eskian, M.; Vaccaro, A.R.; Rahimi-Movaghar, V. Granulocyte Colony-Stimulating Factor (G-CSF) for the Treatment of Spinal Cord Injury. CNS Drugs 2017, 31, 911–937. [Google Scholar] [CrossRef]
- Liao, M.F.; Hsu, J.L.; Lu, K.T.; Chao, P.K.; Cheng, M.Y.; Hsu, H.C.; Lo, A.L.; Lee, Y.L.; Hung, Y.H.; Lyu, R.K.; et al. Granulocyte Colony Stimulating Factor (GCSF) Can Attenuate Neuropathic Pain by Suppressing Monocyte Chemoattractant Protein-1 (MCP-1) Expression, through Upregulating the Early MicroRNA-122 Expression in the Dorsal Root Ganglia. Cells 2020, 9, 1669. [Google Scholar] [CrossRef] [PubMed]
- Ro, L.S.; Chen, S.R.; Chao, P.K.; Lee, Y.L.; Lu, K.T. The potential application of granulocyte colony stimulating factor therapy on neuropathic pain. Chang Gung Med. J. 2009, 32, 235–246. [Google Scholar]
- Liao, M.F.; Yeh, S.R.; Lo, A.L.; Chao, P.K.; Lee, Y.L.; Hung, Y.H.; Lu, K.T.; Ro, L.S. An early granulocyte colony-stimulating factor treatment attenuates neuropathic pain through activation of mu opioid receptors on the injured nerve. Sci. Rep. 2016, 6, 25490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, K.; Koda, M.; Takahashi, H.; Sakuma, T.; Inada, T.; Kamiya, K.; Ota, M.; Maki, S.; Okawa, A.; Takahashi, K.; et al. Granulocyte colony-stimulating factor attenuates spinal cord injury-induced mechanical allodynia in adult rats. J. Neurol. Sci. 2015, 355, 79–83. [Google Scholar] [CrossRef]
- Liou, J.T.; Lui, P.W.; Liu, F.C.; Lai, Y.S.; Day, Y.J. Exogenous granulocyte colony-stimulating factor exacerbate pain-related behaviors after peripheral nerve injury. J. Neuroimmunol. 2011, 232, 83–93. [Google Scholar] [CrossRef]
- Lindenlaub, T.; Sommer, C. Partial sciatic nerve transection as a model of neuropathic pain: A qualitative and quantitative neuropathological study. Pain 2000, 89, 97–106. [Google Scholar] [CrossRef]
- Kalmar, B.; Greensmith, L.; Malcangio, M.; McMahon, S.B.; Csermely, P.; Burnstock, G. The effect of treatment with BRX-220, a co-inducer of heat shock proteins, on sensory fibers of the rat following peripheral nerve injury. Exp. Neurol. 2003, 184, 636–647. [Google Scholar] [CrossRef]
- Lever, I.; Cunningham, J.; Grist, J.; Yip, P.K.; Malcangio, M. Release of BDNF and GABA in the dorsal horn of neuropathic rats. Eur. J. Neurosci. 2003, 18, 1169–1174. [Google Scholar] [CrossRef]
- Loeser, J.D.; Treede, R.D. The Kyoto protocol of IASP Basic Pain Terminology. Pain 2008, 137, 473–477. [Google Scholar] [CrossRef]
- Huh, Y.; Ji, R.R.; Chen, G. Neuroinflammation, Bone Marrow Stem Cells, and Chronic Pain. Front. Immunol. 2017, 8, 1014. [Google Scholar] [CrossRef] [Green Version]
- Chao, P.K.; Lu, K.T.; Lee, Y.L.; Chen, J.C.; Wang, H.L.; Yang, Y.L.; Cheng, M.Y.; Liao, M.F.; Ro, L.S. Early systemic granulocyte-colony stimulating factor treatment attenuates neuropathic pain after peripheral nerve injury. PLoS ONE 2012, 7, e43680. [Google Scholar] [CrossRef]
- Ha, S.O.; Kim, J.K.; Hong, H.S.; Kim, D.S.; Cho, H.J. Expression of brain-derived neurotrophic factor in rat dorsal root ganglia, spinal cord and gracile nuclei in experimental models of neuropathic pain. Neuroscience 2001, 107, 301–309. [Google Scholar] [CrossRef]
- Terada, Y.; Morita-Takemura, S.; Isonishi, A.; Tanaka, T.; Okuda, H.; Tatsumi, K.; Shinjo, T.; Kawaguchi, M.; Wanaka, A. NGF and BDNF expression in mouse DRG after spared nerve injury. Neurosci. Lett. 2018, 686, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Kleinschnitz, C.; Schroeter, M.; Jander, S.; Stoll, G. Induction of granulocyte colony-stimulating factor mRNA by focal cerebral ischemia and cortical spreading depression. Brain Res. Mol. Brain Res. 2004, 131, 73–78. [Google Scholar] [CrossRef]
- Schneider, A.; Kruger, C.; Steigleder, T.; Weber, D.; Pitzer, C.; Laage, R.; Aronowski, J.; Maurer, M.H.; Gassler, N.; Mier, W.; et al. The hematopoietic factor G-CSF is a neuronal ligand that counteracts programmed cell death and drives neurogenesis. J. Clin. Investig. 2005, 115, 2083–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, A.; Kuhn, H.G.; Schabitz, W.R. A role for G-CSF (granulocyte-colony stimulating factor) in the central nervous system. Cell Cycle 2005, 4, 1753–1757. [Google Scholar] [PubMed] [Green Version]
- Liongue, C.; Wright, C.; Russell, A.P.; Ward, A.C. Granulocyte colony-stimulating factor receptor: Stimulating granulopoiesis and much more. Int. J. Biochem. Cell Biol. 2009, 41, 2372–2375. [Google Scholar] [CrossRef] [PubMed]
- Villa, G.; Fumagalli, M.; Verderio, C.; Abbracchio, M.P.; Ceruti, S. Expression and contribution of satellite glial cells purinoceptors to pain transmission in sensory ganglia: An update. Neuron Glia Biol. 2010, 6, 31–42. [Google Scholar] [CrossRef]
- Goldschmidt, E.; Fellows-Mayle, W.; Wolfe, R.; Niranjan, A.; Flickinger, J.C.; Lunsford, L.D.; Gerszten, P.C. Radiosurgery to the spinal dorsal root ganglion induces fibrosis and inhibits satellite glial cell activation while preserving axonal neurotransmission. J. Neurosurg. Spine 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Schweizerhof, M.; Stosser, S.; Kurejova, M.; Njoo, C.; Gangadharan, V.; Agarwal, N.; Schmelz, M.; Bali, K.K.; Michalski, C.W.; Brugger, S.; et al. Hematopoietic colony-stimulating factors mediate tumor-nerve interactions and bone cancer pain. Nat. Med. 2009, 15, 802–807. [Google Scholar] [CrossRef]
- Zhang, E.; Lee, S.; Yi, M.H.; Nan, Y.; Xu, Y.; Shin, N.; Ko, Y.; Lee, Y.H.; Lee, W.; Kim, D.W. Expression of granulocyte colony-stimulating factor 3 receptor in the spinal dorsal horn following spinal nerve ligation-induced neuropathic pain. Mol. Med. Rep. 2017, 16, 2009–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diederich, K.; Sevimli, S.; Dorr, H.; Kosters, E.; Hoppen, M.; Lewejohann, L.; Klocke, R.; Minnerup, J.; Knecht, S.; Nikol, S.; et al. The role of granulocyte-colony stimulating factor (G-CSF) in the healthy brain: A characterization of G-CSF-deficient mice. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 11572–11581. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, C.-C.; Yang, C.-P.; Ma, K.-H.; Shih, J.-H.; Tseng, C.-S.; Huang, Y.-S. Endogenous Expression of G-CSF in Rat Dorsal Root Ganglion Neurons after Nerve Injury. Brain Sci. 2021, 11, 956. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070956
Yeh C-C, Yang C-P, Ma K-H, Shih J-H, Tseng C-S, Huang Y-S. Endogenous Expression of G-CSF in Rat Dorsal Root Ganglion Neurons after Nerve Injury. Brain Sciences. 2021; 11(7):956. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070956
Chicago/Turabian StyleYeh, Chun-Chang, Chih-Ping Yang, Kuo-Hsing Ma, Jui-Hu Shih, Ching-San Tseng, and Yuahn-Sieh Huang. 2021. "Endogenous Expression of G-CSF in Rat Dorsal Root Ganglion Neurons after Nerve Injury" Brain Sciences 11, no. 7: 956. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070956