
Brain Abnormalities in Individuals with a Desire for a Healthy Limb Amputation: Somatosensory, Motoric or Both? A Task-Based fMRI Verdict

 , , , , ,
, , , , ,
Abstract
:1. Introduction
1.1. Sensory Functions in BID
1.2. Similarities and Differences between BID and Somatoparaphrenia
1.3. A Neurological Origin for BID?
1.4. Brain Morphometry and Resting State fMRI Studies
1.5. Task Based Functional Imaging Studies
1.6. Summary of Imaging Studies and Outstanding Issues
1.7. Aims of the Present Study
1.8. Predictions
2. Materials and Methods
2.1. Participants
2.2. Clinical Interview
2.3. Neurological and Psychiatric Assessment
2.4. Zurich Xenomelia Scale
2.5. FMRI Experiment
2.5.1. Tactile Stimulation Task
2.5.2. Hand Motor Execution Task
2.5.3. Foot Motor Execution Task
2.6. fMRI Data Acquisition and Analyses
2.6.1. fMRI Data Acquisition
2.6.2. Preprocessing
2.6.3. Statistical Analysis of the fMRI Data
First-Level Fixed-Effect Analysis
Second Level Random-Effect Analysis
Conjunction Analyses
Comparison of Conjunction Maps across Groups
3. Results
3.1. Demographic and Clinical Data
3.2. fMRI Results
3.2.1. Within Group Activations for Somatosensory Stimulation and Motor Tasks
Somatosensory Stimulation Task
Motor Execution Task
3.2.2. Between Group Comparisons
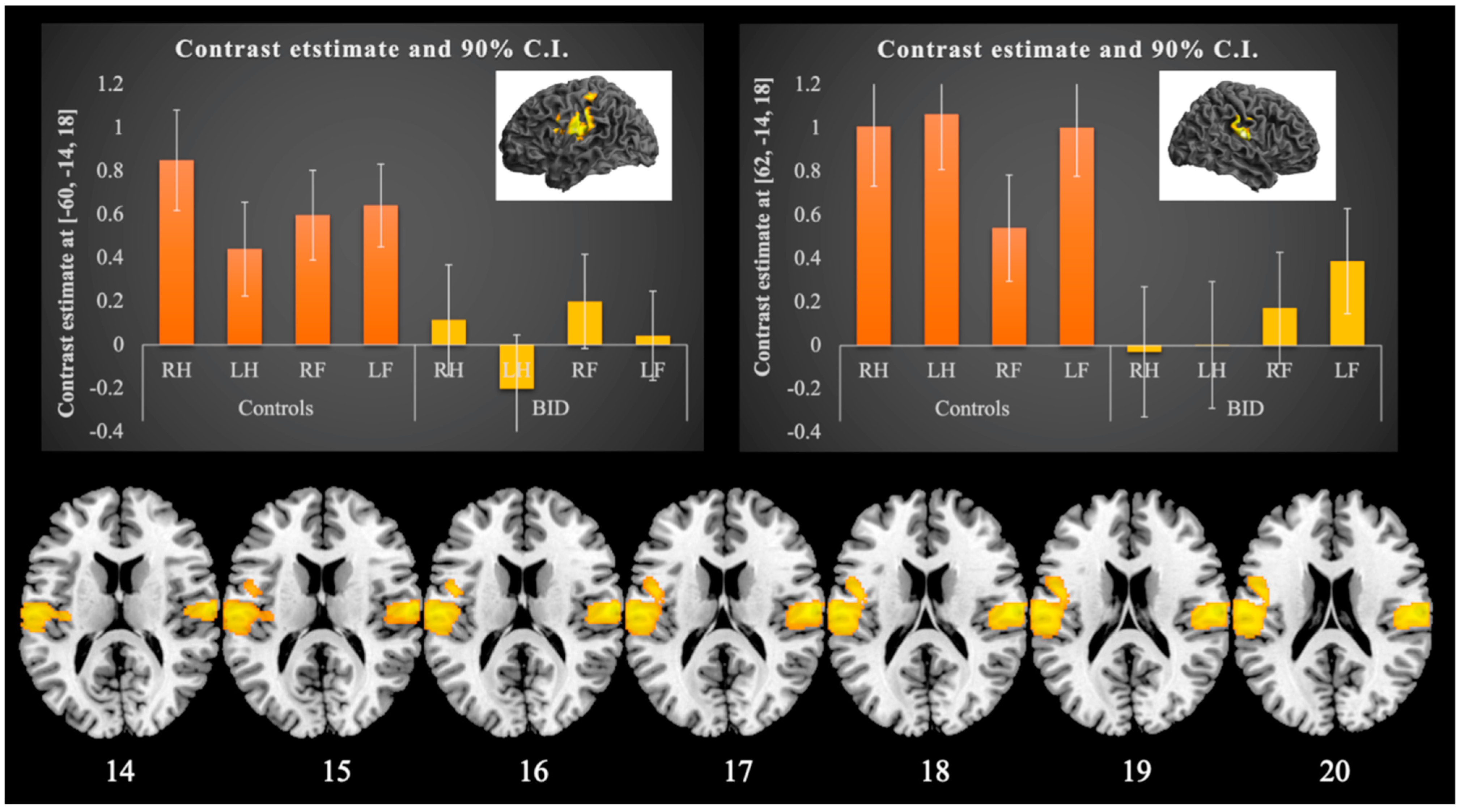
Somatosensory Stimulation Task
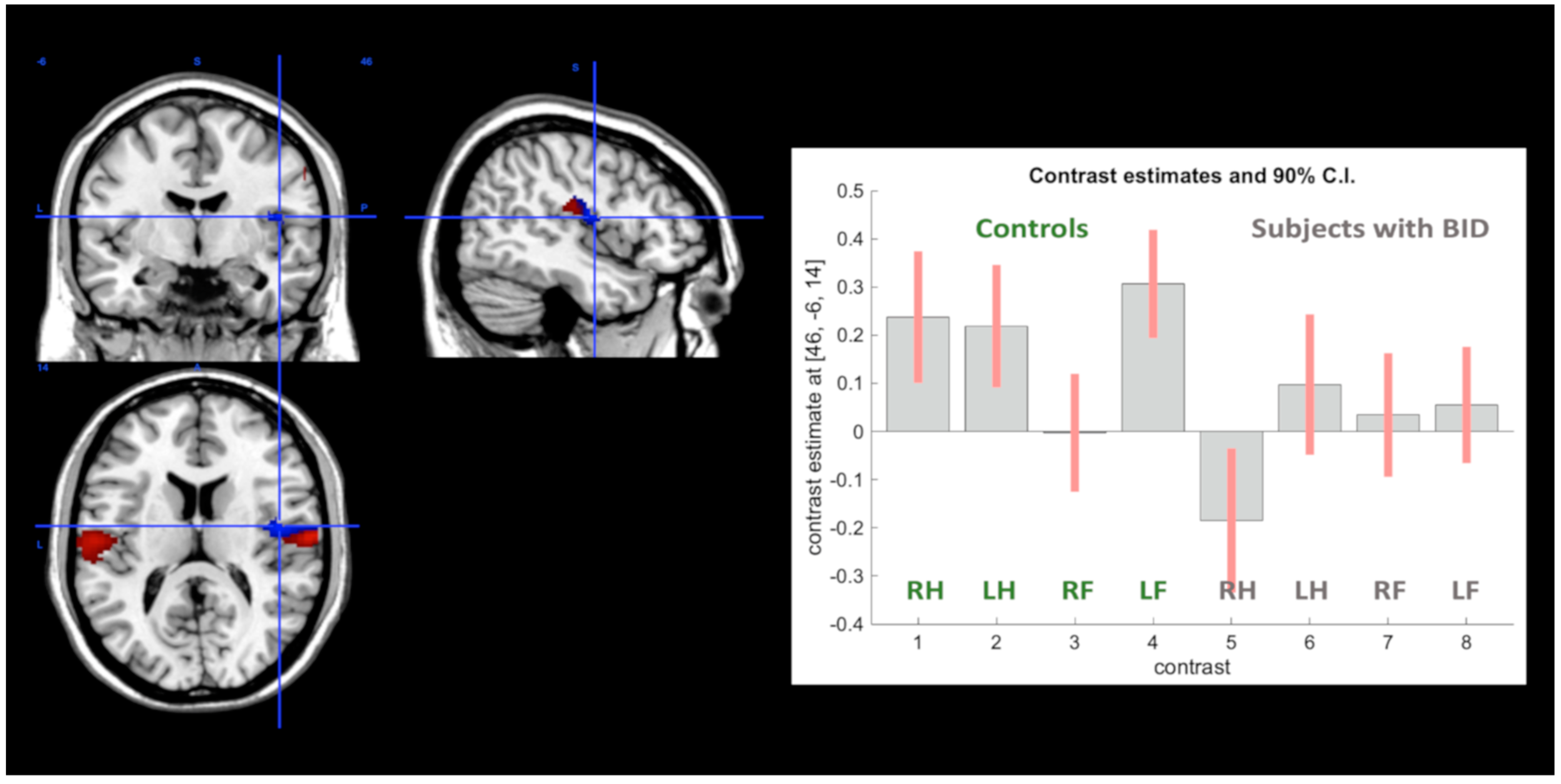
Motor Execution Task
3.2.3. Conjunction Analyses
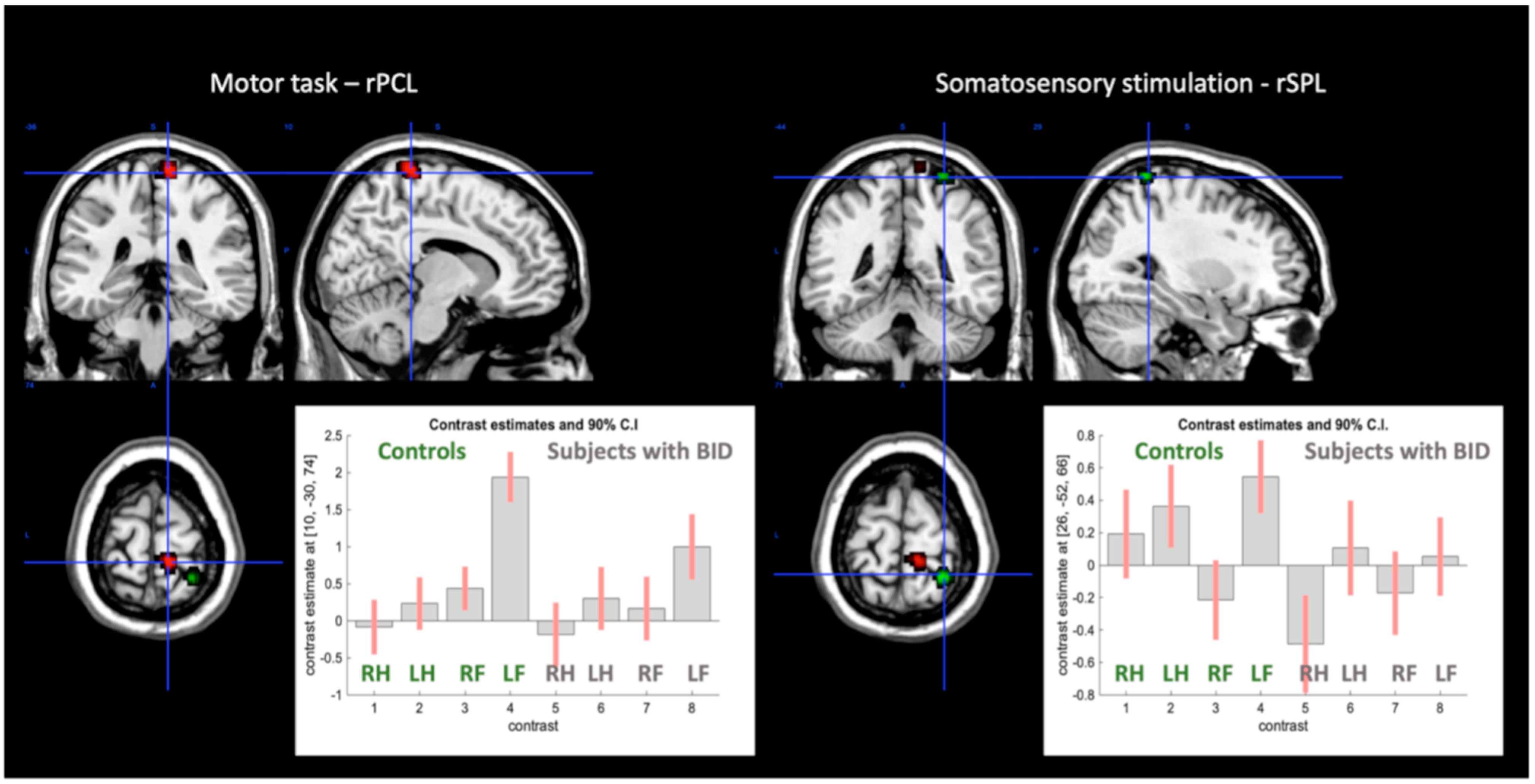
Conjunction of the Effects of the Somatosensory Stimulations
Conjunction of the Effects of Motor Execution
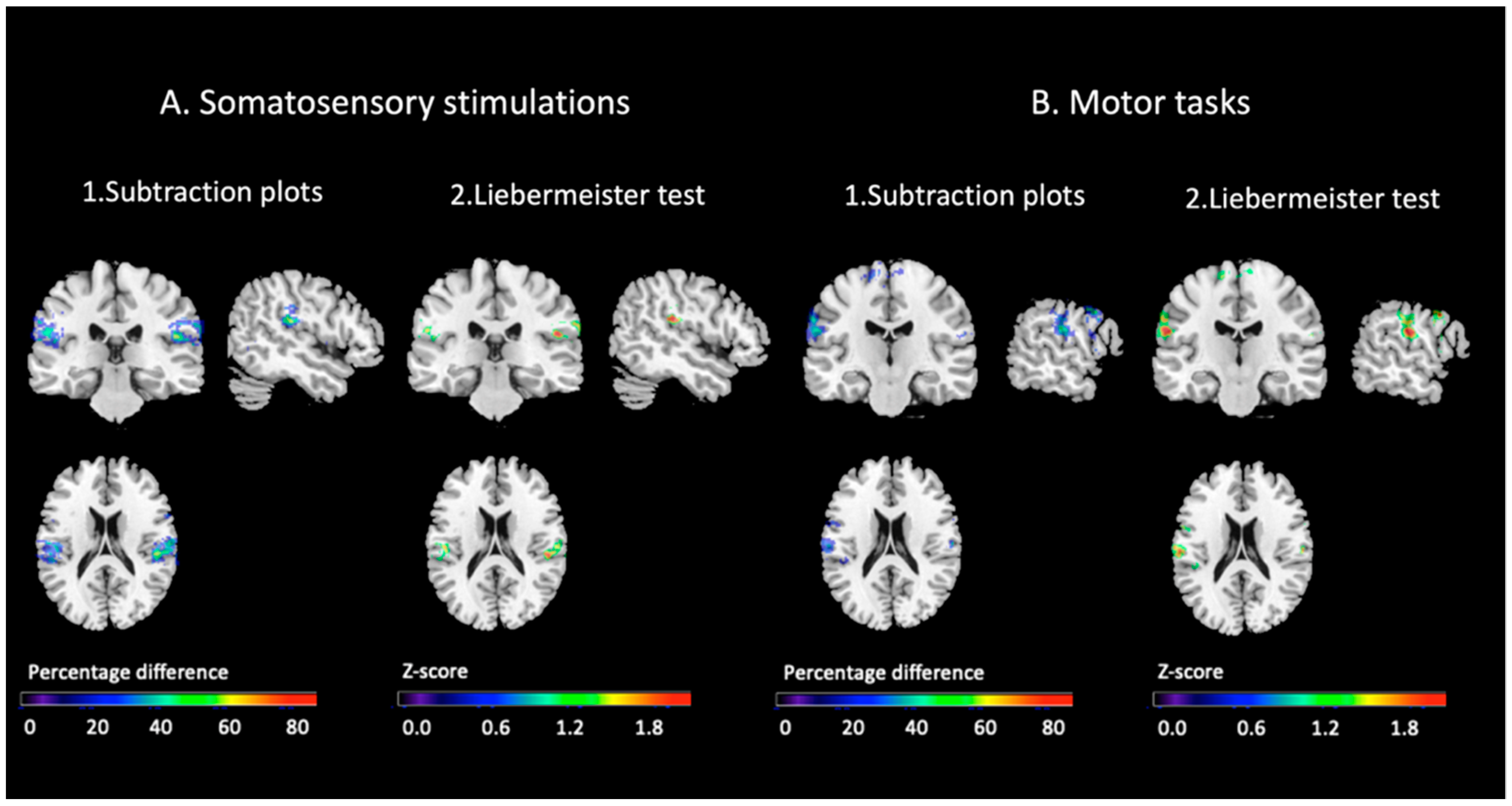
3.2.4. Comparisons of the Topographical Distribution of the Conjunction Effects
4. Discussion
5. Limitations of the Study
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sedda, A. Body integrity identity disorder: From a psychological to a neurological syndrome. Neuropsychol. Rev. 2011, 21, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Sedda, A.; Bottini, G. Apotemnophilia, body integrity identity disorder or xenomelia? Psychiatric and neurologic etiologies face each other. Neuropsychiatr Dis. Treat. 2014, 10, 1255–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrone, D. Disfigured anatomies and imperfect analogies: Body integrity identity disorder and the supposed right to self-demanded amputation of healthy body parts. J. Med. Ethics. 2009, 35, 541–545. [Google Scholar] [CrossRef]
- Blom, R.M.; Hennekam, R.C.; Denys, D. Body integrity identity disorder. PLoS ONE 2012, 7, e34702. [Google Scholar] [CrossRef] [Green Version]
- Bruno, R. Devotees, Pretenders and Wannabes: Two Cases of Factitious Disability Disorder. Sex. Disabil. 1997, 15, 243–260. [Google Scholar] [CrossRef]
- Giummarra, M.J.; Bradshaw, J.L.; Hilti, L.M.; Nicholls, M.E.; Brugger, P. Paralyzed by desire: A new type of body integrity identity disorder. Cogn. Behav. Neurol. Off. J. Soc. Behav. Cogn. Neurol. 2012, 25, 34–41. [Google Scholar] [CrossRef]
- First, M.B. Desire for amputation of a limb: Paraphilia, psychosis, or a new type of identity disorder. Psychol. Med. 2005, 35, 919–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everaerd, W. A case of apotemnophilia: A handicap as sexual preference. Am. J. Psychother. 1983, 37, 285–293. [Google Scholar] [CrossRef]
- Money, J.; Jobaris, R.; Furth, G. Apotemnophilia: Two cases of self-demand amputation as a paraphilia. J. Sex Res. 1977, 13, 115–125. [Google Scholar] [CrossRef]
- Lawrence, A.A. Clinical and theoretical parallels between desire for limb amputation and gender identity disorder. Arch. Sex. Behav. 2006, 35, 263–278. [Google Scholar] [CrossRef] [Green Version]
- McGeoch, P.D.; Brang, D.; Song, T.; Lee, R.R.; Huang, M.; Ramachandran, V.S. Xenomelia: A new right parietal lobe syndrome. J. Neurol. Neurosurg. Psychiatry 2011, 82, 1314–1319. [Google Scholar] [CrossRef] [Green Version]
- Blanke, O.; Morgenthaler, F.D.; Brugger, P.; Overney, L.S. Preliminary evidence for a fronto-parietal dysfunction in able-bodied participants with a desire for limb amputation. J. Neuropsychol. 2009, 3, 181–200. [Google Scholar] [CrossRef]
- Johnson, A.J.; Liew, S.-L.; Aziz-Zadeh, L. Demographics, learning and imitation, and body schema in body integrity identity disorder. Indiana Univ. Undergrad. J. Cogn. Sci. 2011, 6. [Google Scholar]
- Hilti, L.M.; Hanggi, J.; Vitacco, D.A.; Kraemer, B.; Palla, A.; Luechinger, R.; Jancke, L.; Brugger, P. The desire for healthy limb amputation: Structural brain correlates and clinical features of xenomelia. Brain 2013, 136, 318–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, K.D.; Kornblad, C.A.E.; Engel, M.M.; Dijkerman, H.C.; Blom, R.M.; Keizer, A. An Investigation of Lower Limb Representations Underlying Vision, Touch, and Proprioception in Body Integrity Identity Disorder. Front. Psychiatry 2020, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Stone, K.D.; Dijkerman, H.C.; Bekrater-Bodmann, R.; Keizer, A. Mental rotation of feet in individuals with Body Integrity Identity Disorder, lower-limb amputees, and normally-limbed controls. PLoS ONE 2019, 14, e0221105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerstmann, J. Problem of imperception of disease and of impaired body territories with organic lesions. Relation to body schema and its disorders. Arch. Neurol. Psychiatry 1942, 48, 890–913. [Google Scholar] [CrossRef]
- Ramachandran, V.S.; McGeoch, P. Can vestibular caloric stimulation be used to treat apotemnophilia? Med. Hypotheses 2007, 69, 250–252. [Google Scholar] [CrossRef]
- Gandola, M.; Invernizzi, P.; Sedda, A.; Ferre, E.R.; Sterzi, R.; Sberna, M.; Paulesu, E.; Bottini, G. An anatomical account of somatoparaphrenia. Cortex 2012, 48, 1165–1178. [Google Scholar] [CrossRef]
- Vallar, G.; Ronchi, R. Somatoparaphrenia: A body delusion. A review of the neuropsychological literature. Exp. Brain Res. 2009, 192, 533–551. [Google Scholar] [CrossRef]
- Brang, D.; McGeoch, P.D.; Ramachandran, V.S. Apotemnophilia: A neurological disorder. Neuroreport 2008, 19, 1305–1306. [Google Scholar] [CrossRef] [PubMed]
- Romano, D.; Sedda, A.; Brugger, P.; Bottini, G. Body ownership: When feeling and knowing diverge. Conscious. Cogn. 2015, 34, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Blom, R.M.; van Wingen, G.A.; van der Wal, S.J.; Luigjes, J.; van Dijk, M.T.; Scholte, H.S.; Denys, D. The Desire for Amputation or Paralyzation: Evidence for Structural Brain Anomalies in Body Integrity Identity Disorder (BIID). PLoS ONE 2016, 11, e0165789. [Google Scholar] [CrossRef] [PubMed]
- Hänggi, J.; Bellwald, D.; Brugger, P. Shape alterations of basal ganglia and thalamus in xenomelia. Neuroimage Clin 2016, 11, 760–769. [Google Scholar] [CrossRef] [Green Version]
- Hänggi, J.; Vitacco, D.A.; Hilti, L.M.; Luechinger, R.; Kraemer, B.; Brugger, P. Structural and functional hyperconnectivity within the sensorimotor system in xenomelia. Brain Behav. 2017, 7, e00657. [Google Scholar] [CrossRef] [Green Version]
- Oddo-Sommerfeld, S.; Hanggi, J.; Coletta, L.; Skoruppa, S.; Thiel, A.; Stirn, A.V. Brain activity elicited by viewing pictures of the own virtually amputated body predicts xenomelia. Neuropsychologia 2018, 108, 135–146. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, M.T.; van Wingen, G.A.; van Lammeren, A.; Blom, R.M.; de Kwaasteniet, B.P.; Scholte, H.S.; Denys, D. Neural basis of limb ownership in individuals with body integrity identity disorder. PLoS ONE 2013, 8, e72212. [Google Scholar] [CrossRef] [Green Version]
- Saetta, G.; Hänggi, J.; Gandola, M.; Zapparoli, L.; Salvato, G.; Berlingeri, M.; Sberna, M.; Paulesu, E.; Bottini, G.; Brugger, P. Neural Correlates of Body Integrity Dysphoria. Curr. Biol. 2020, 30, 2191–2195. [Google Scholar] [CrossRef]
- Moseley, G.L.; Gallace, A.; Spence, C. Bodily illusions in health and disease: Physiological and clinical perspectives and the concept of a cortical ‘body matrix’. Neurosci. Biobehav. Rev. 2012, 36, 34–46. [Google Scholar] [CrossRef]
- Berlucchi, G.; Aglioti, S.M. The body in the brain revisited. Exp. Brain Res. 2010, 200, 25–35. [Google Scholar] [CrossRef]
- So, E.L.; Schauble, B.S. Ictal asomatognosia as a cause of epileptic falls: Simultaneous video, EMG, and invasive EEG. Neurology 2004, 63, 2153–2154. [Google Scholar] [CrossRef]
- Arzy, S.; Overney, L.S.; Landis, T.; Blanke, O. Neural mechanisms of embodiment: Asomatognosia due to premotor cortex damage. Arch. Neurol. 2006, 63, 1022–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arzy, S.; Thut, G.; Mohr, C.; Michel, C.M.; Blanke, O. Neural basis of embodiment: Distinct contributions of temporoparietal junction and extrastriate body area. J. Neurosci. 2006, 26, 8074–8081. [Google Scholar] [CrossRef] [PubMed]
- Baier, B.; Karnath, H.O. Tight link between our sense of limb ownership and self-awareness of actions. Stroke 2008, 39, 486–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeller, D.; Gross, C.; Bartsch, A.; Johansen-Berg, H.; Classen, J. Ventral premotor cortex may be required for dynamic changes in the feeling of limb ownership: A lesion study. J. Neurosci. 2011, 31, 4852–4857. [Google Scholar] [CrossRef] [Green Version]
- Tsakiris, M. My body in the brain: A neurocognitive model of body-ownership. Neuropsychologia 2010, 48, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Berti, A.; Bottini, G.; Gandola, M.; Pia, L.; Smania, N.; Stracciari, A.; Castiglioni, I.; Vallar, G.; Paulesu, E. Shared cortical anatomy for motor awareness and motor control. Science 2005, 309, 488–491. [Google Scholar] [CrossRef] [Green Version]
- Gandola, M.; Bottini, G.; Zapparoli, L.; Invernizzi, P.; Verardi, M.; Sterzi, R.; Santilli, I.; Sberna, M.; Paulesu, E. The physiology of motor delusions in anosognosia for hemiplegia: Implications for current models of motor awareness. Conscious. Cogn. 2014, 24, 98–112. [Google Scholar] [CrossRef]
- Zapparoli, L.; Seghezzi, S.; Devoto, F.; Mariano, M.; Banfi, G.; Porta, M.; Paulesu, E. Altered sense of agency in Gilles de la Tourette syndrome: Behavioural, clinical and functional magnetic resonance imaging findings. Brain Commun. 2020, 2, fcaa204. [Google Scholar] [CrossRef]
- Bottini, G.; Paulesu, E.; Gandola, M.; Loffredo, S.; Scarpa, P.; Sterzi, R.; Santilli, I.; Defanti, C.A.; Scialfa, G.; Fazio, F.; et al. Left caloric vestibular stimulation ameliorates right hemianesthesia. Neurology 2005, 65, 1278–1283. [Google Scholar] [CrossRef]
- Pons, T.P.; Garraghty, P.E.; Friedman, D.P.; Mishkin, M. Physiological evidence for serial processing in somatosensory cortex. Science 1987, 237, 417–420. [Google Scholar] [CrossRef]
- Paulesu, E.; Frackowiak, R.; Bottini, G. Maps of somatosensory systems. In Human Brain Function; Frackowiak, R., Friston, K., Frith, C., Dolan, R., Mazziotta, J., Eds.; Academic Press: Cambridge, MA, USA, 1997; pp. 183–242. [Google Scholar]
- Zapparoli, L.; Seghezzi, S.; Zirone, E.; Guidali, G.; Tettamanti, M.; Banfi, G.; Bolognini, N.; Paulesu, E. How the effects of actions become our own. Sci. Adv. 2020, 6. [Google Scholar] [CrossRef]
- Bisiach, E.; Vallar, G.; Perani, D.; Papagno, C.; Berti, A. Unawareness of disease following lesions of the right hemisphere: Anosognosia for hemiplegia and anosognosia for hemianopia. Neuropsychologia 1986, 24, 471–482. [Google Scholar] [CrossRef]
- Aoyama, A.; Krummenacher, P.; Palla, A.; Hilti, L.M.; Brugger, P. Impaired Spatial-Temporal Integration of Touch in Xenomelia (Body Integrity Identity Disorder). Spat. Cogn. Comput. 2012, 12, 96–110. [Google Scholar] [CrossRef]
- Friston, K.J.; Ashburner, J.; Poline, J.B.; Frith, C.D.; Heather, J.D.; Frackowiak, R.S.J. Spatial registration and normalisation of images. Hum. Brain Mapp. 1995, 2, 165–189. [Google Scholar] [CrossRef]
- Ashburner, J.; Friston, K.J. Nonlinear spatial normalization using basis functions. Hum. Brain Mapp. 1999, 7, 254–266. [Google Scholar] [CrossRef]
- Worsley, K.J.; Friston, K.J. Analysis of fMRI time-series revisited--again. Neuroimage 1995, 2, 173–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, A.P.; Friston, K.J. Generalisability, Random Effects and Population inference. Neuroimage 1988, 7, S754. [Google Scholar] [CrossRef]
- Penny, W.D.; Holmes, A.P. Random-effects analysis. In Human Brain Function; Frackowiak, R.S.J., Ashburner, J.T., Penny, W.D., Zeki, S., Friston, K.J., Frith, C.D., Dolan, R.J., Price, C.J., Eds.; Elsevier: San Diego, CA, USA, 2004; pp. 843–850. [Google Scholar]
- Flandin, G.; Friston, K.J. Analysis of family-wise error rates in statistical parametric mapping using random field theory. Hum. Brain Mapp. 2017. [Google Scholar] [CrossRef] [PubMed]
- Eklund, A.; Nichols, T.E.; Knutsson, H. Cluster failure: Why fMRI inferences for spatial extent have inflated false-positive rates. Proc. Natl. Acad. Sci. USA 2016, 113, 7900–7905. [Google Scholar] [CrossRef] [Green Version]
- Rorden, C.; Karnath, H.O.; Bonilha, L. Improving lesion-symptom mapping. J. Cogn. Neurosci. 2007, 19, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Gandola, M.; Bruno, M.; Zapparoli, L.; Saetta, G.; Rolandi, E.; De Santis, A.; Banfi, G.; Zerbi, A.; Sansone, V.; Paulesu, E. Functional brain effects of hand disuse in patients with trapeziometacarpal joint osteoarthritis: Executed and imagined movements. Exp. Brain Res. 2017, 235, 3227–3241. [Google Scholar] [CrossRef]
- Gandola, M.; Zapparoli, L.; Saetta, G.; De Santis, A.; Zerbi, A.; Banfi, G.; Sansone, V.; Bruno, M.; Paulesu, E. Thumbs up: Imagined hand movements counteract the adverse effects of post-surgical hand immobilization. Clinical, behavioral, and fMRI longitudinal observations. Neuroimage Clin. 2019, 23, 101838. [Google Scholar] [CrossRef] [PubMed]
- Ruben, J.; Schwiemann, J.; Deuchert, M.; Meyer, R.; Krause, T.; Curio, G.; Villringer, K.; Kurth, R.; Villringer, A. Somatotopic organization of human secondary somatosensory cortex. Cereb. Cortex. 2001, 11, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Giummarra, M.J.; Bradshaw, J.L.; Nicholls, M.E.; Hilti, L.M.; Brugger, P. Body integrity identity disorder: Deranged body processing, right fronto-parietal dysfunction, and phenomenological experience of body incongruity. Neuropsychol. Rev. 2011, 21, 320–333. [Google Scholar] [CrossRef]
- Invernizzi, P.; Gandola, M.; Romano, D.; Zapparoli, L.; Bottini, G.; Paulesu, E. What is mine? Behavioral and anatomical dissociations between somatoparaphrenia and anosognosia for hemiplegia. Behav. Neurol. 2013, 26, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Saetta, G.; Zindel-Geisseler, O.; Stauffacher, F.; Serra, C.; Vannuscorps, G.; Brugger, P. Asomatognosia: Structured Interview and Assessment of Visuomotor Imagery. Front. Psychol. 2020, 11, 544544. [Google Scholar] [CrossRef]
- Saetta, G.; Michels, L.; Brugger, P. Where in the brain is “the other’s” hand? Mapping dysfunctional neural networks in somatoparaphrenia. Neuroscience 2021. manuscript accepted. [Google Scholar] [CrossRef]
- Critchley, M. The Parietal Lobes; Williams and Wilkins: Oxford, UK, 1953. [Google Scholar]
- Loetscher, T.; Regard, M.; Brugger, P. Misoplegia: A review of the literature and a case without hemiplegia. J. Neurol. Neurosurg. Psychiatry 2006, 77, 1099–1100. [Google Scholar] [CrossRef] [Green Version]
- Pia, L.; Garbarini, F.; Fossataro, C.; Fornia, L.; Berti, A. Pain and body awareness: Evidence from brain-damaged patients with delusional body ownership. Front. Hum. Neurosci. 2013, 7, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carruthers, G. Types of body representation and the sense of embodiment. Conscious. Cogn. 2008, 17, 1302–1316. [Google Scholar] [CrossRef] [PubMed]
- Tsakiris, M.; Fotopoulou, A. Is my body the sum of online and offline body-representations? Conscious. Cogn. 2008, 17, 1317–1320. [Google Scholar] [CrossRef]
- Robinson, C.J.; Burton, H. Somatotopographic organization in the second somatosensory area of M. fascicularis. J. Comp. Neurol. 1980, 192, 43–67. [Google Scholar] [CrossRef] [PubMed]
- Hihara, S.; Taoka, M.; Tanaka, M.; Iriki, A. Visual Responsiveness of Neurons in the Secondary Somatosensory Area and its Surrounding Parietal Operculum Regions in Awake Macaque Monkeys. Cereb. Cortex. 2015, 25, 4535–4550. [Google Scholar] [CrossRef] [PubMed]
- Bretas, R.V.; Taoka, M.; Suzuki, H.; Iriki, A. Secondary somatosensory cortex of primates: Beyond body maps, toward conscious self-in-the-world maps. Exp. Brain Res. 2020, 238, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Corradi-Dell’Acqua, C.; Tomasino, B.; Fink, G.R. What is the position of an arm relative to the body? Neural correlates of body schema and body structural description. J. Neurosci. 2009, 29, 4162–4171. [Google Scholar] [CrossRef] [PubMed]
- Graziano, M.S.; Hu, X.T.; Gross, C.G. Visuospatial properties of ventral premotor cortex. J. Neurophysiol. 1997, 77, 2268–2292. [Google Scholar] [CrossRef] [PubMed]
- Brugger, P.; Lenggenhager, B.; Giummarra, M.J. Xenomelia: A social neuroscience view of altered bodily self-consciousness. Front. Psychol. 2013, 4, 204. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Saetta, G.; Simon, C.; Lenggenhager, B.; Ruddy, K.L. Could Brain-Computer Interface be a new therapeutic approach for Body Integrity Dysphoria? Front. Hum. Neurosci. 2021, 15, 699830. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author(s) 1 | Year | Sample Size | Side | Body Part | Technique | Analysis | Structural/Functional Abnormalities | |||
|---|---|---|---|---|---|---|---|---|---|---|
| BID | C | L | R | B | ||||||
| McGeoch et al. [11] | 2011 | 4 | 4 | 1 | 2 | 1 | Leg | MEG | ROIs-based | Reduced activity in the rSPL. |
| Hilti et al. [14] | 2013 | 13 *,^ | 13 | 8 | 2 | 3 | Leg | MRI—surface-based morphometry | ROIs-based | Reduced thickness: RSPL. Increased thickness: rCS, rIPL. Reduced cortical surface area: rIPL, rSI, rSII, r anterior IC. Increased cortical surface area: lIPL, lSII. |
| van Dijk et al. [27] | 2013 | 5 | 10 | 2 | 3 | 0 | Leg | Task-based fMRI | Whole brain | Tactile task: reduced activation in the PMCd and PMCv for the affected leg; Heightened responsivity in a large somatosensory network regardless of which leg was stimulated. Motor task: no differences. |
| Blom et al. [23] | 2016 | 8 | 24 | 2 | 3 | 3 # | Leg | MRI (VBM) | Whole brain and SVC in predefined ROIs | Reduced grey matter concentration: ldPMC, lvPMC; Increased grey matter concentration: lCerebellum. |
| Hänggi et al. [24] | 2016 | 13 * | 13 | 8 | 2 | 3 | Leg | MRI—FIRST | Subcortical regions | Thinning (hypotrophy): bilateral dorsomedial putamen, left ventromedial caudate, left medial pallidum. Thickening (hypertrophy): bilateral lateral pallidum; left frontolateral thalamus. |
| Hänggi et al. [25] | 2017 | 13 * | 13 | 8 | 2 | 3 | Leg | MRI; DTI; rsfMRI | Whole brain connectome; network analyses in predefined nodes | Hyperconnected nodes: Connectome analysis: Paracentral lobule, SMA, Postcentral gyrus, BG, Cerebellum. Network analyses: SPL, SI, SII, PMC, BG, Thalamus, IC. |
| Oddo et al. [26] | 2018 | 12 | 11 | 9 | 1 | 2 | Leg | fMRI | Machine-learning—whole brain | Brain activity evocated by viewing images of the own virtually amputated body predicts BID. bSPL, IPL, caudate nucleus and occipital regions are among the highest predictive regions. |
| Saetta et al. [28] | 2020 | 16 ** | 16 | 16 | 0 | 0 | Leg | RsfMRI—MRI | Whole brain rsfMRI and ROIs-based MRI (VBM) | rPCL; rSPL, lSTG, and lIFGOrb, were less functionally connected to the rest of the brain. lPMC and rSPL, and lIFGOrb were atrophic. |
| Participants with BID | Demographic Features | BID Features | Mean Scores on Zurich Xenomelia Scale * | |||||
|---|---|---|---|---|---|---|---|---|
| Age | Education | Limb | Side | Pure Amputation Desire | Erotic Attraction | Pretending Behaviour | Total Scale Scores | |
| P1 | 42 | 13 | leg | left | 4.50 | 4.25 | 5.00 | 4.58 |
| P2 | 36 | 13 | leg | left | 4.50 | 4.25 | 5.00 | 4.58 |
| P3 | 36 | 13 | leg | left | 5.25 | 3.25 | 3.00 | 3.83 |
| P4 | 48 | 18 | leg | left | 6.00 | 3.50 | 4.00 | 4.50 |
| P5 | 34 | 18 | leg | left | 5.25 | 2.75 | 3.25 | 3.75 |
| P6 | 37 | 18 | leg | left | 6.00 | 6.00 | 4.75 | 5.58 |
| P7 | 41 | 13 | leg | left | 5.75 | 4.50 | 5.00 | 5.08 |
| P8 | 39 | 18 | leg | left | 6.00 | 4.25 | 5.50 | 5.25 |
| P9 | 64 | 13 | leg | left | 6.00 | 5.50 | 4.50 | 5.33 |
| P10 | 38 | 14 | leg | left | 6.00 | 3.75 | 3.00 | 4.25 |
| Brain Region (BA) | Cluster | Peak | |||||
|---|---|---|---|---|---|---|---|
| K | PFWER-corr | Puncorrected | Z-Score | MNI Coordinates | |||
| x | y | z | |||||
| Cluster 1 | 765 | 0.005 | 0.002 | ||||
| R Postcentral gyrus | 5.06 # | 62 | −14 | 18 | |||
| R Postcentral gyrus (43) | 3.65 | 62 | −10 | 38 | |||
| R Supramarginal gyrus (2) | 4.52 # | 66 | −22 | 34 | |||
| Cluster 2 | 1625 | <0.001 | <0.001 | ||||
| L Supramarginal gyrus (1) | 4.72 # | −62 | −22 | 36 | |||
| L Postcentral gyrus | 4.66 # | −60 | −14 | 18 | |||
| L Precentral gyrus (6) | 4.18 | −50 | 2 | 22 | |||
| Cluster 3 | 174 | 0.254 | 0.090 | ||||
| R Postcentral gyrus—SPL ** | 3.51 | 26 | −46 | 64 | |||
| R Inf. parietal gyrus (3) | 3.46 | 30 | −42 | 54 | |||
| Brain Regions (BA) | Cluster | Peak | |||||
|---|---|---|---|---|---|---|---|
| K | PFWER-corr | Puncorrected | Z-Score | MNI Coordinates | |||
| x | y | z | |||||
| (1) MOTOR TASKS | |||||||
| (a) Conjunction Controls and BID | |||||||
| Cluster 1 | 468 | 0.021 | 0.006 | ||||
| R SMA (6) | 4.52 # | 6 | −4 | 60 | |||
| L SMA (6) | 3.70 | −8 | −12 | 64 | |||
| (b) Conjunction Controls | |||||||
| Cluster 1 | 1600 | <0.001 | <0.001 | ||||
| R SMA | 7.0 # | 2 | −4 | 58 | |||
| R Mid. Cingulum (24) | 3.29 | 10 | 6 | 36 | |||
| Cluster 2 | 845 | 0.002 | <0.001 | ||||
| L Precentral gyrus (6) | 5.27 # | −60 | 6 | 28 | |||
| L Precentral gyrus (6) | 4.59 # | −46 | −6 | 54 | |||
| L Rolandic opercular gyrus | 4.33 | −48 | 2 | 10 | |||
| Cluster 3 | 432 | 0.027 | 0.007 | ||||
| R Supramarginal gyrus | 4.55 # | 62 | −16 | 22 | |||
| R Supramarginal gyrus | 4.05 | 54 | −26 | 34 | |||
| R Superior temporal gyrus | 3.12 | 58 | −34 | 24 | |||
| Cluster 4 | 297 | 0.079 | 0.022 | ||||
| L Supramarginal gyrus | 4.49 # | −56 | −22 | 24 | |||
| Cluster 5 | 463 | 0.022 | 0.006 | ||||
| R Insula | 3.93 | 44 | 2 | 6 | |||
| R Rolandic opercular gyrus | 3.83 | 54 | 8 | 14 | |||
| R Putamen | 3.62 | 28 | −2 | 0 | |||
| (c) Conjunction BID | |||||||
| Cluster 1 | 470 | 0.021 | 0.005 | ||||
| R SMA (6) | 4.52 # | 6 | −4 | 60 | |||
| L SMA (6) | 3.70 | −8 | −12 | 64 | |||
| Cluster 2 | 461 | 0.022 | 0.006 | ||||
| R Rolandic opercular gyrus | 3.82 | 60 | −18 | 16 | |||
| R Supramarginal gyrus** | 3.73 | 48 | −26 | 28 | |||
| R Supramarginal gyrus | 3.68 | 56 | −24 | 36 | |||
| (2) SOMATOSENSORY TASKS | |||||||
| (a) Conjunctions in Controls and BID | |||||||
| No areas of shared activations | |||||||
| (b) Conjunctions in Controls | |||||||
| Cluster 1 | 956 | 0.002 | 0.001 | ||||
| L Postcentral gyrus | 5.15 # | −54 | −18 | 18 | |||
| L Supramarginal gyrus | 4.97 # | −54 | −26 | 22 | |||
| L Supramarginal gyrus (43) | 4.29 | −60 | −20 | 36 | |||
| Cluster 2 | 735 | 0.006 | 0.002 | ||||
| R Rolandic opercular gyrus | 4.93 # | 50 | −28 | 22 | |||
| (c) Conjunctions in BID | |||||||
| No areas of shared activations | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandola, M.; Zapparoli, L.; Saetta, G.; Reverberi, C.; Salvato, G.; Squarza, S.A.C.; Invernizzi, P.; Sberna, M.; Brugger, P.; Bottini, G.; et al. Brain Abnormalities in Individuals with a Desire for a Healthy Limb Amputation: Somatosensory, Motoric or Both? A Task-Based fMRI Verdict. Brain Sci. 2021, 11, 1248. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11091248
Gandola M, Zapparoli L, Saetta G, Reverberi C, Salvato G, Squarza SAC, Invernizzi P, Sberna M, Brugger P, Bottini G, et al. Brain Abnormalities in Individuals with a Desire for a Healthy Limb Amputation: Somatosensory, Motoric or Both? A Task-Based fMRI Verdict. Brain Sciences. 2021; 11(9):1248. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11091248
Chicago/Turabian StyleGandola, Martina, Laura Zapparoli, Gianluca Saetta, Carlo Reverberi, Gerardo Salvato, Silvia Amaryllis Claudia Squarza, Paola Invernizzi, Maurizio Sberna, Peter Brugger, Gabriella Bottini, and et al. 2021. "Brain Abnormalities in Individuals with a Desire for a Healthy Limb Amputation: Somatosensory, Motoric or Both? A Task-Based fMRI Verdict" Brain Sciences 11, no. 9: 1248. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11091248