
Investigating the Migraine Cycle over 21 Consecutive Days Using Proton Magnetic Resonance Spectroscopy and Resting-State fMRI: A Pilot Study

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient
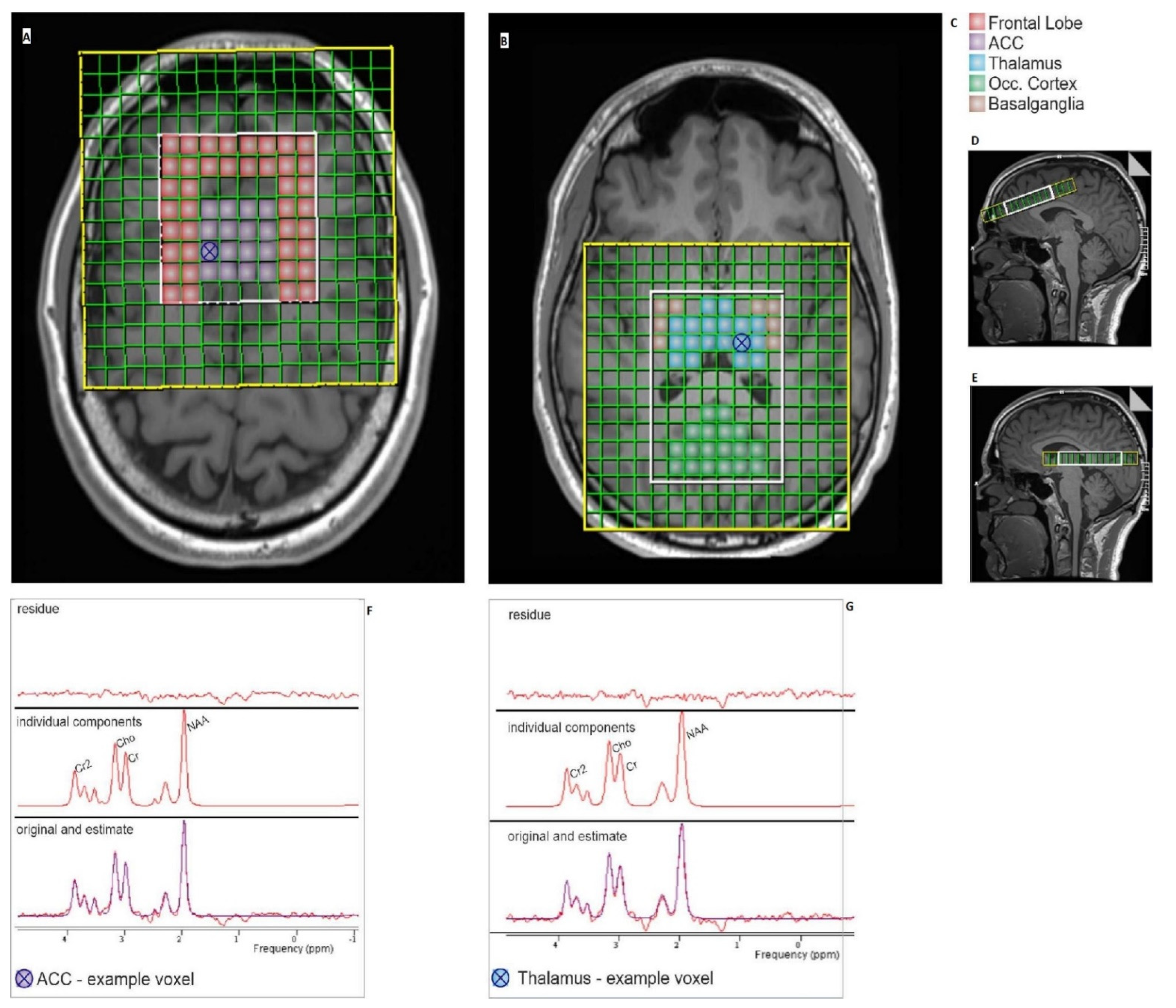
2.2. Image Acquisition
2.3. Data Processing and Analysis
2.3.1. MR—Spectroscopy Analysis
2.3.2. Resting-State Analysis
3. Results
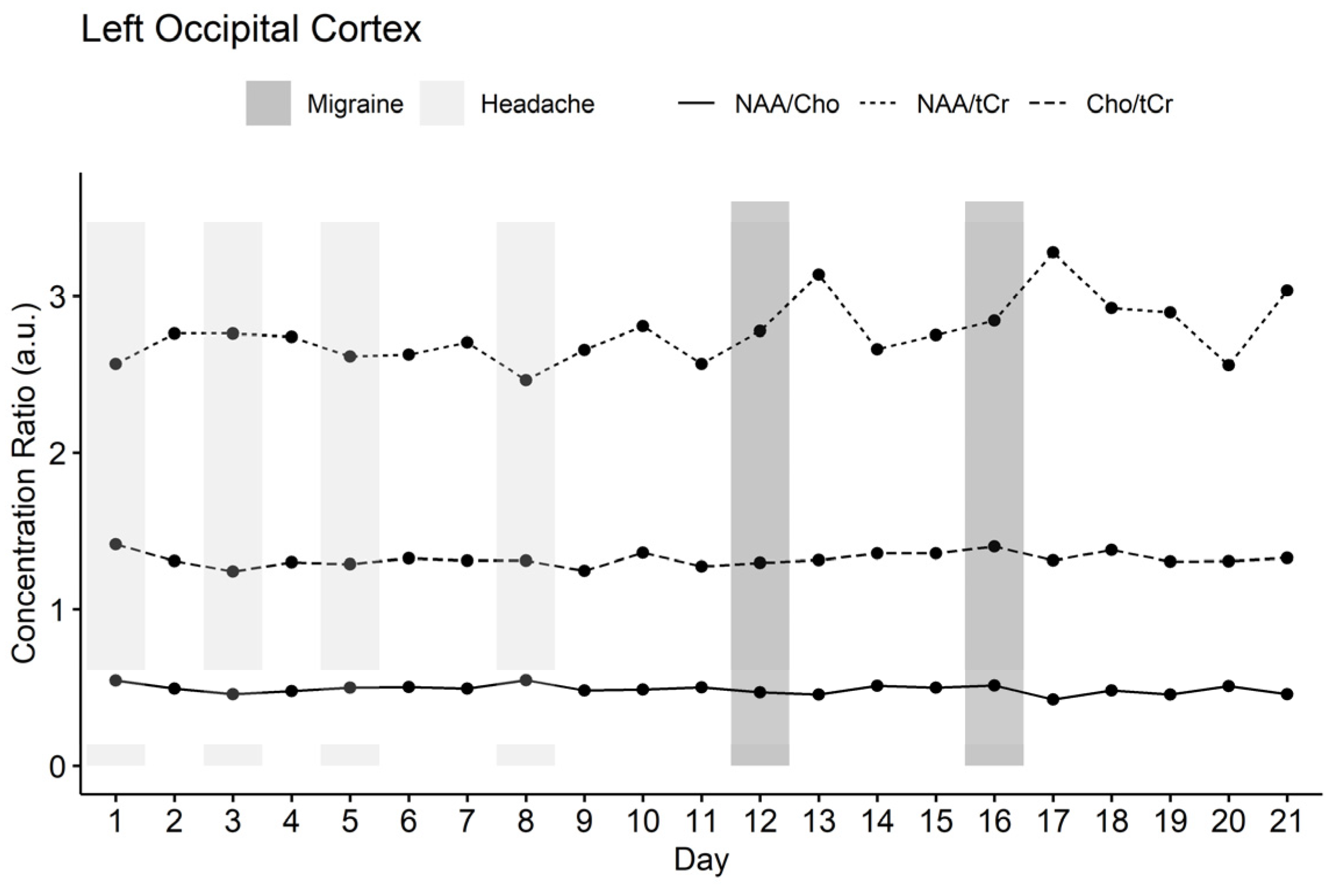
3.1. MR Spectroscopy Results
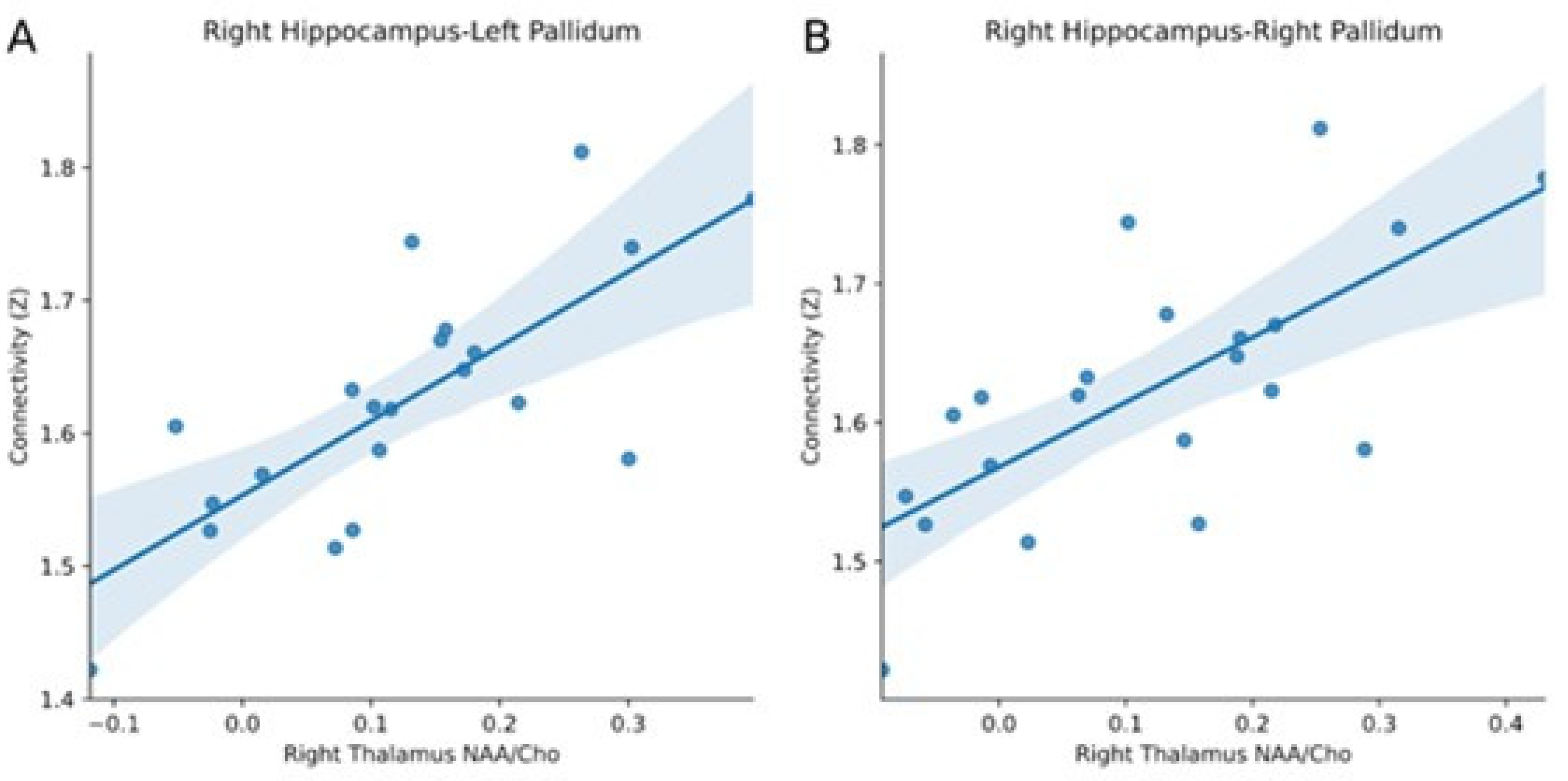
3.2. Comparison of Resting-State fMRI and MR Spectroscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woldeamanuel, Y.W.; Cowan, R.P. Migraine affects 1 in 10 people worldwide featuring recent rise: A systematic review and meta-analysis of community-based studies involving 6 million participants. J. Neurol. Sci. 2017, 372, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Headache Classification Committee of the International Headache Society (IHS). The International Classification of Headache Disorders, 3rd ed.; Cephalalgia 2018, 38, 1–211. [CrossRef] [PubMed]
- Ashina, S.; Bentivegna, E.; Martelletti, P.; Eikermann-Haerter, K. Structural and Functional Brain Changes in Migraine. Pain Ther. 2021, 10, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Schulte, L.H.; May, A. The migraine generator revisited: Continuous scanning of the migraine cycle over 30 days and three spontaneous attacks. Brain 2016, 139, 1987–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, F.M.; Hougaard, A.; Magon, S.; Sprenger, T.; Wolfram, F.; Rostrup, E.; Ashina, M. Altered thalamic connectivity during spontaneous attacks of migraine without aura: A resting-state fMRI study. Cephalalgia 2018, 38, 1237–1244. [Google Scholar] [CrossRef]
- Schulte, L.H.; Menz, M.M.; Haaker, J.; May, A. The migraineur’s brain networks: Continuous resting-state fMRI over 30 days. Cephalalgia 2020, 40, 1614–1621. [Google Scholar] [CrossRef]
- Reyngoudt, H.; Achten, E.; Paemeleire, K. Magnetic resonance spectroscopy in migraine: What have we learned so far? Cephalalgia 2012, 32, 845–859. [Google Scholar] [CrossRef] [Green Version]
- Bathel, A.; Schweizer, L.; Stude, P.; Glaubitz, B.; Wulms, N.; Delice, S.; Schmidt-Wilcke, T. Increased thalamic glutamate/glutamine levels in migraineurs. J. Headache Pain 2018, 19, 55. [Google Scholar] [CrossRef]
- Zielman, R.; Wijnen, J.; Webb, A.; Onderwater, G.; Ronen, I.; Ferrari, M.D.; Kan, H.; Terwindt, G.M.; Kruit, M. Cortical glutamate in migraine. Brain 2017, 140, 1859–1871. [Google Scholar] [CrossRef]
- Bigal, M.E.; Hetherington, H.; Pan, J.; Tsang, A.; Grosberg, B.; Avdievich, N.; Friedman, B.; Lipton, R.B. Occipital levels of GABA are related to severe headaches in migraine. Neurology 2008, 70, 2078–2080. [Google Scholar] [CrossRef] [Green Version]
- de Tommaso, M.; Ceci, E.; Pica, C.; Trojano, M.; Delussi, M.; Franco, G.; Livrea, P.; Ruggieri, M. Serum levels of N-acetyl-aspartate in migraine and tension-type headache. J. Headache Pain 2012, 13, 389–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younis, S.; Hougaard, A.; Vestergaard, M.B.; Larsson, H.B.; Ashina, M. Migraine and magnetic resonance spectroscopy: A systematic review. Curr. Opin. Neurol. 2017, 30, 246–262. [Google Scholar] [CrossRef] [PubMed]
- del Río, M.S.; Linera, J.A. Functional neuroimaging of headaches. Lancet Neurol. 2004, 3, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Noseda, R.; Jakubowski, M.; Kainz, V.; Borsook, D.; Burstein, R. Cortical projections of functionally identified thalamic trigeminovascular neurons: Implications for migraine headache and its associated symptoms. J. Neurosci. 2011, 31, 14204–14217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, K.D.; Taylor, S.J.; Crawley, A.P.; Wood, M.; Mikulis, D. Functional MRI of pain and attention-related activations in the human cingulate cortex. J. Neurophysiol. 1997, 7, 3370–3380. [Google Scholar] [CrossRef]
- Vanhammea, L.; Boogaart, A.V.D.; Van Huffel, S. Improved method for accurate and efficient quantification of MRS data with use of prior knowledge. J. Magn. Reson. 1997, 129, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The MRUI Consortium. jMRUI-Software for the Clinical and Biomedical MRS. 2021. Available online: http://www.jmrui.eu/ (accessed on 24 May 2021).
- Kreis, R. Issues of spectral quality in clinical 1H-magnetic resonance spectroscopy and a gallery of artifacts. NMR Biomed. 2004, 17, 361–381. [Google Scholar] [CrossRef]
- Dezortova, M.; Jiru, F.; Petrasek, J.; Malinova, V.; Zeman, J.; Jirsa, M.; Hajek, M. 1H MR spectroscopy as a diagnostic tool for cerebral creatine deficiency. Magn. Reason. Mater. Phys. Biol. Med. 2008, 21, 327–332. [Google Scholar] [CrossRef]
- Scheau, C.; Preda, E.; Popa, G.; Ghergus, A.; Capsa, R.; Lupescu, I. Magnetic resonance spectroscopy—A non-invasive method in evaluating focal and diffuse central nervous system disease. J. Med. Life 2012, 5, 423–427. [Google Scholar]
- Ramadan, S.; Lin, A.; Stanwell, P. Glutamate and glutamine: A review of in vivo MRS in the human brain. NMR Biomed. 2013, 26, 1630–1646. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Clark, J. N-acetyl aspartate: A marker for neuronal loss or mitochondrial dysfunction. Dev. Neurosci. 1998, 20, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Graybiel, A.M. Network-level neuroplasticity in cortico-basal ganglia pathways. Parkinsonism Relat. Disord. 2004, 10, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.A.; Rodriguez, M.C.; DeLong, M.R. Basal ganglia pathophysiology: A critical review. Adv. Neurol. 1997, 74, 3–18. [Google Scholar] [PubMed]
- Chudler, E.H.; Dong, W.K. The role of the basal ganglia in nociception and pain. Pain 1995, 60, 3–38. [Google Scholar] [CrossRef]
- Maleki, N.; Becerra, L.; Nutile, L.; Pendse, G.; Brawn, J.; Bigal, M.; Burstein, R.; Borsook, D. Migraine attacks the Basal Ganglia. Mol. Pain 2011, 21, 71. [Google Scholar] [CrossRef] [Green Version]
- Moulton, E.A.; Becerra, L.; Maleki, N.; Pendse, G.; Tully, S.; Hargreaves, R.; Burstein, R.; Borsook, D. Painful Heat Reveals Hyperexcitability of the Temporal Pole in Interictal and Ictal Migraine States. Cereb. Cortex 2011, 21, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Chabriat, H.; Bousser, M.-G. Neuropsychiatric manifestations in CADASIL. Dialogues Clin. Neurosci. 2007, 9, 199–208. [Google Scholar] [CrossRef]
- Chaudhary, S.; Kumaran, S.S.; Goyal, V.; Kalaivani, M.; Kaloiya, G.S.; Sagar, R.; Mehta, N.; Srivastava, A.K.; Jagannathan, N.R. Frontal lobe metabolic alterations characterizing Parkinson’s disease cognitive impairment. Neurol. Sci. 2021, 42, 1053–1064. [Google Scholar] [CrossRef]
- de Tommaso, M.; Vecchio, E.; Quitadamo, S.; Coppola, G.; Di Renzo, A.; Parisi, V.; Silvestro, M.; Russo, A.; Tedeschi, G. Pain-Related Brain Connectivity Changes in Migraine: A Narrative Review and Proof of Concept about Possible Novel Treatments Interference. Brain Sci. 2021, 11, 234. [Google Scholar] [CrossRef]
- Arika, W.M.; Kibiti, C.M.; Njagi, J.M.; Ngugi, M.P. Effects of DCM Leaf Extract of Gnidia glauca (Fresen) on Locomotor Activity, Anxiety, and Exploration-Like Behaviors in High-Fat Diet-Induced Obese Rats. Behav. Neurol. 2019, 2019, 7359235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Huang, J.; Zhang, Z.; Cao, Z. Altered Metabolites in the Occipital Lobe in Migraine Without Aura During the Attack and the Interictal Period. Front. Neurol. 2021, 12, 656349. [Google Scholar] [CrossRef] [PubMed]
- Bulboacă, A.E.; Stănescu, I.C.; Bolboacă, S.D.; Bulboacă, A.C.; Bodizs, G.I.; Nicula, C.A. Retinal Nerve Fiber Layer Thickness and Oxidative Stress Parameters in Migraine Patients without Aura: A Pilot Study. Antioxidants 2020, 9, 494. [Google Scholar] [CrossRef] [PubMed]
- Vincent, M.B. Vision and migraine. Headache J. Head Face Pain 2015, 55, 595–599. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filippi, V.; Steiger, R.; Beliveau, V.; Frank, F.; Kaltseis, K.; Gizewski, E.R.; Broessner, G. Investigating the Migraine Cycle over 21 Consecutive Days Using Proton Magnetic Resonance Spectroscopy and Resting-State fMRI: A Pilot Study. Brain Sci. 2022, 12, 646. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050646
Filippi V, Steiger R, Beliveau V, Frank F, Kaltseis K, Gizewski ER, Broessner G. Investigating the Migraine Cycle over 21 Consecutive Days Using Proton Magnetic Resonance Spectroscopy and Resting-State fMRI: A Pilot Study. Brain Sciences. 2022; 12(5):646. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050646
Chicago/Turabian StyleFilippi, Vera, Ruth Steiger, Vincent Beliveau, Florian Frank, Katharina Kaltseis, Elke R. Gizewski, and Gregor Broessner. 2022. "Investigating the Migraine Cycle over 21 Consecutive Days Using Proton Magnetic Resonance Spectroscopy and Resting-State fMRI: A Pilot Study" Brain Sciences 12, no. 5: 646. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050646