
Pentraxin-3 in the Spinal Dorsal Horn Upregulates Nectin-1 Expression in Neuropathic Pain after Spinal Nerve Damage in Male Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Surgical Procedure
2.3. Reagents and Administration
2.4. Behavioral Tests
2.5. Immunoblot
2.6. Quantitative Real-Time PCR (qRT-PCR)
2.7. Statistical Analysis
3. Results
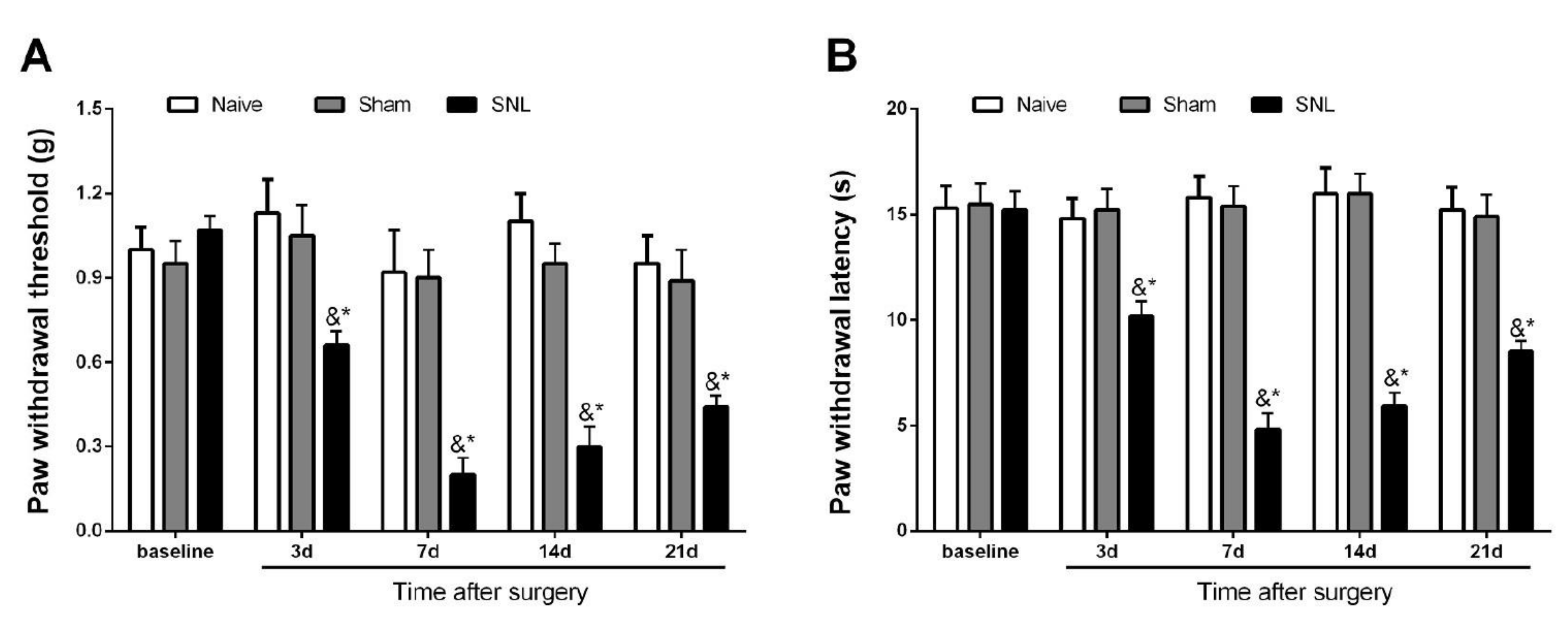
3.1. Initiation and Persistence of Mechanical Allodynia and Thermal Hyperalgesia Following Spinal Nerve Trauma in Male Mice
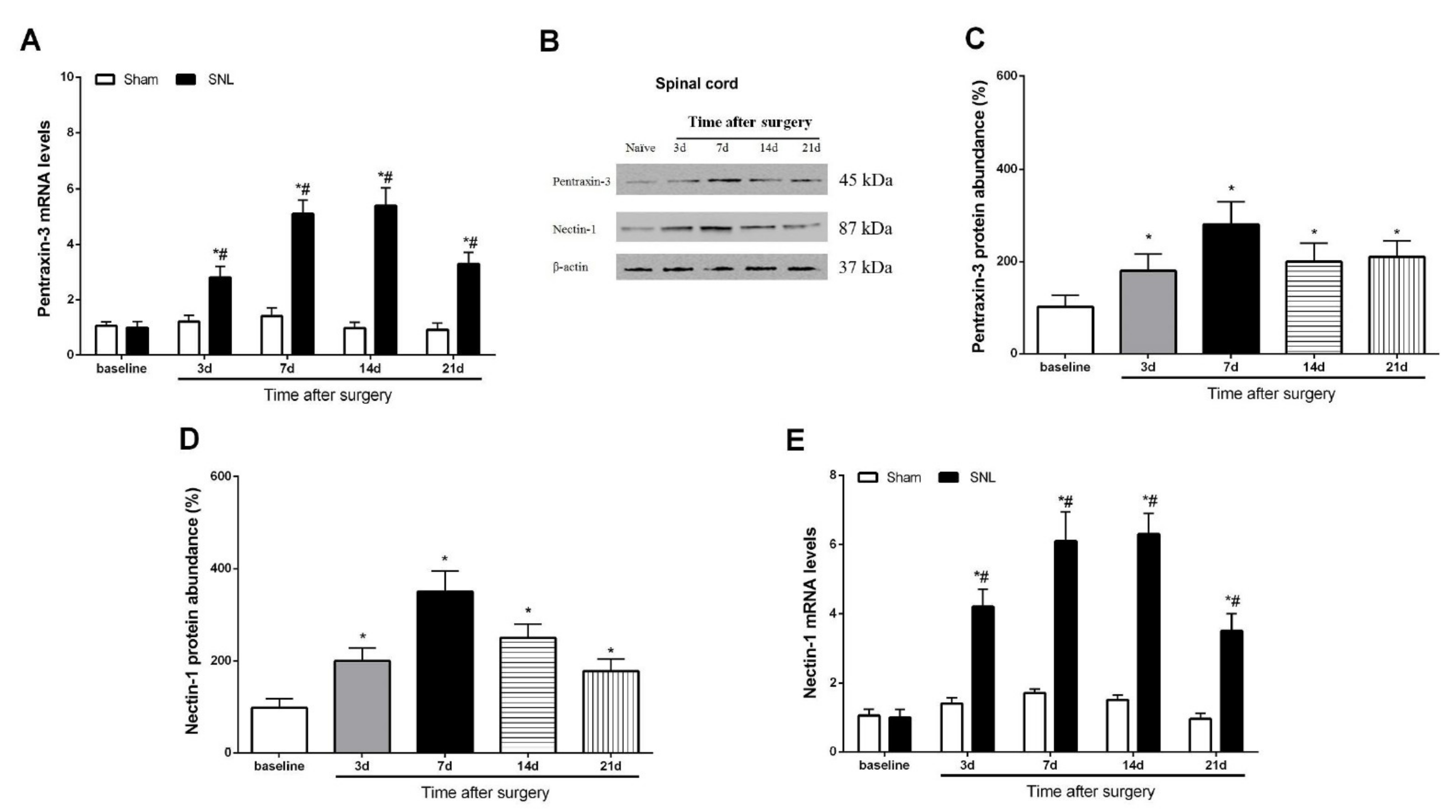
3.2. Pentraxin-3 and Nectin-1 Are Upregulated in the Spinal Cord Dorsal Horn upon Spinal Nerve Injury in Male Mice
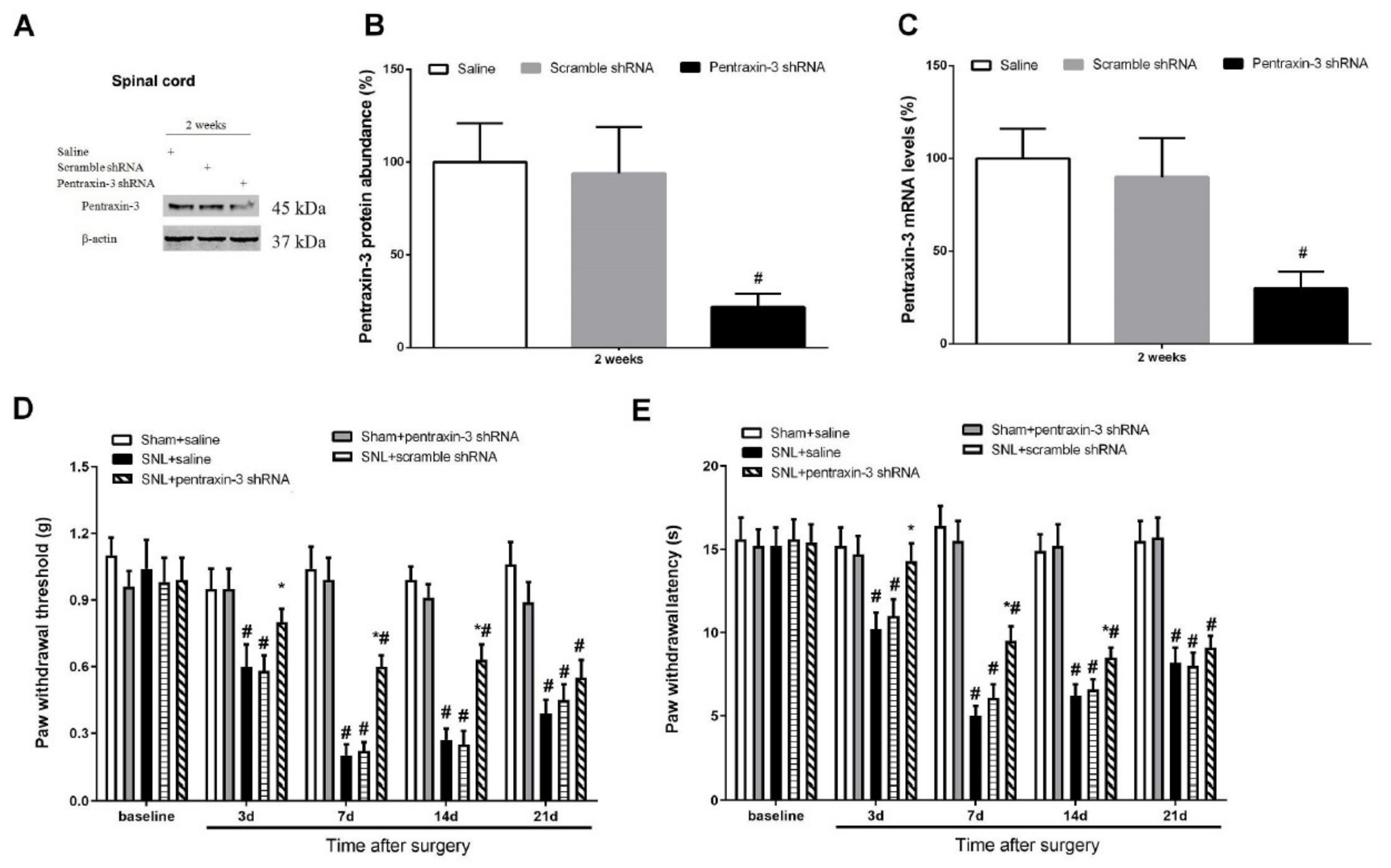
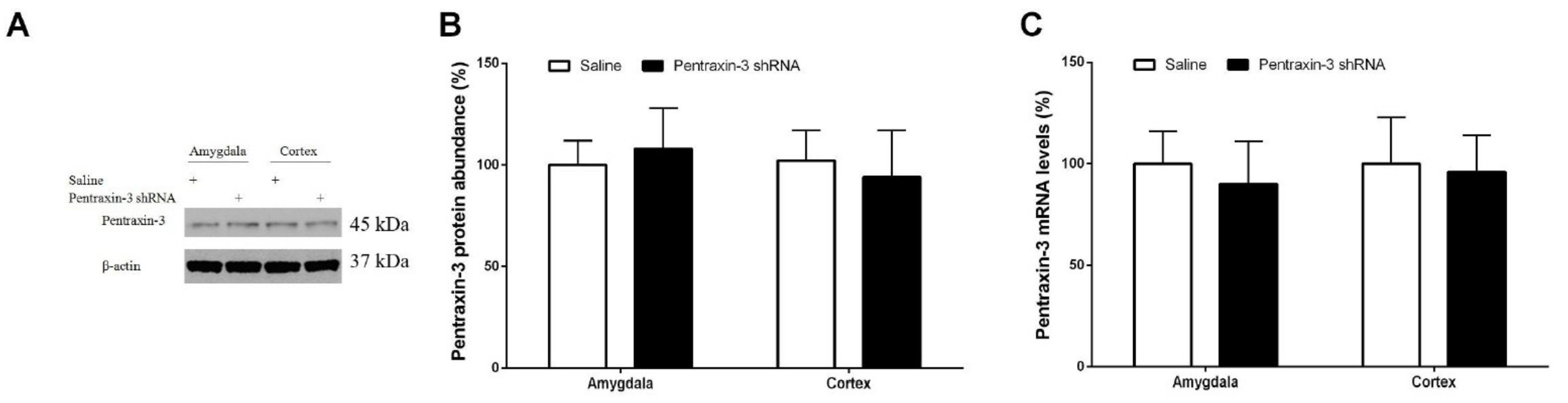
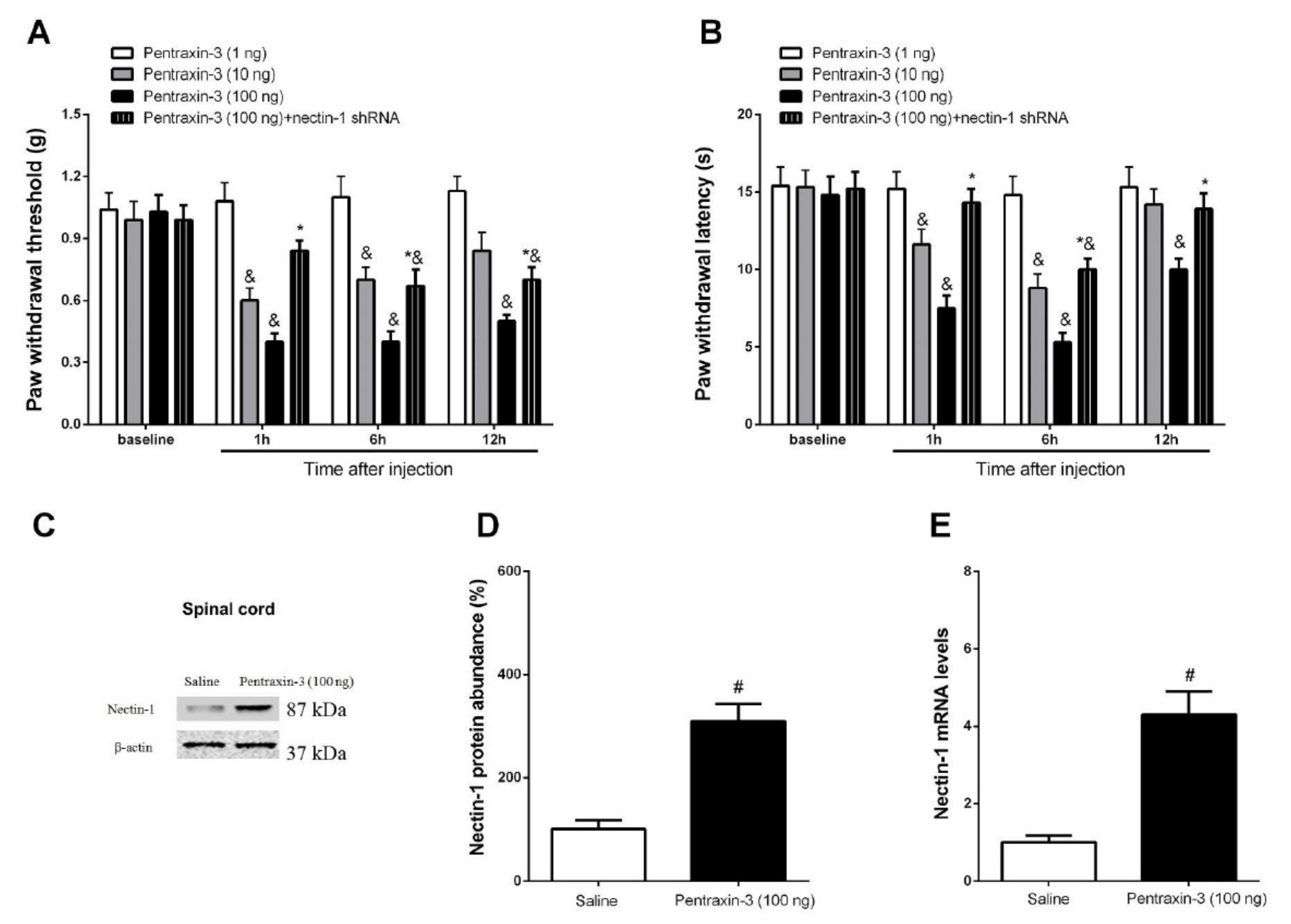
3.3. Pentraxin-3 Knockdown Reduces Neuropathic Pain Behaviors and Spinal Nectin-1 Expression after Peripheral Nerve Damage in Male Mice
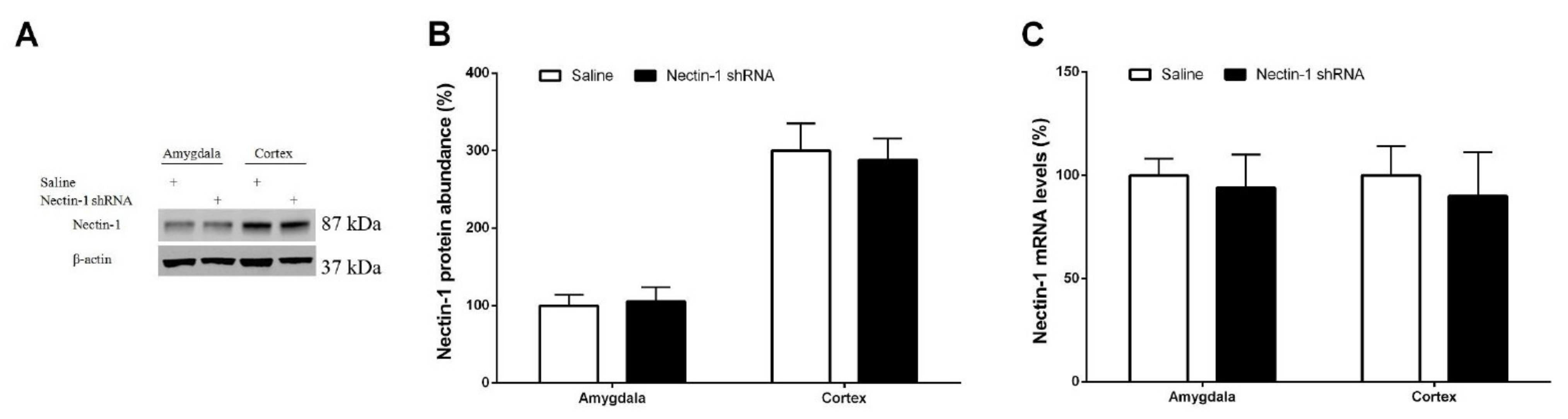
3.4. Spinal Nectin-1 Deficiency Impairs the Development of Neuropathic Pain after Peripheral Nerve Damage in Male Mice
3.5. Inhibition of Recombinant Pentraxin-3 Evokes Acute Pain Behaviors by Spinal Nectin-1 Knockdown in Male Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Johannes, C.B.; Le, T.K.; Zhou, X.; Johnston, J.A.; Dworkin, R.H. The prevalence of chronic pain in United States adults: Results of an Internet-based survey. J. Pain 2010, 11, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Woolf, C.J.; Mannion, R.J. Neuropathic pain: Aetiology, symptoms, mechanisms, and management. Lancet 1999, 353, 1959–1964. [Google Scholar] [CrossRef]
- Ji, R.R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.R.; Donnelly, C.R.; Nedergaard, M. Astrocytes in chronic pain and itch. Nat. Rev. Neurosci. 2019, 20, 667–685. [Google Scholar] [CrossRef]
- St John Smith, E. Advances in understanding nociception and neuropathic pain. J. Neurol. 2018, 265, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Kuner, T.; Kuner, R. Synaptic plasticity in pathological pain. Trends Neurosci. 2014, 37, 343–355. [Google Scholar] [CrossRef]
- Liu, P.; Song, C.; Wang, C.; Li, Y.; Su, L.; Li, J.; Zhao, Q.; Wang, Z.; Shen, M.; Wang, G.; et al. Spinal SNAP-25 regulates membrane trafficking of GluA1- containing AMPA receptors in spinal injury-induced neuropathic pain in rats. Neurosci. Lett. 2020, 715, 134616. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Xie, X.; Zhao, H.; Gao, Y.; Li, Y.; Xu, X.; Zhang, X.; Ke, C.; Liu, J. Promotion of bone cancer pain development by decorin is accompanied by modification of excitatory synaptic molecules in the spinal cord. Mol. Pain 2019, 15, 1744806919864253. [Google Scholar] [CrossRef]
- Cui, W.; Li, Y.; Wang, Z.; Song, C.; Yu, Y.; Wang, G.; Li, J.; Wang, C.; Zhang, L. Spinal caspase-6 regulates AMPA receptor trafficking and dendritic spine plasticity through netrin-1 in postoperative pain after orthopedic surgery for tibial fracture in mice. Pain 2020, 162, 124–134. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, S.; Zhao, Q.; Li, Y.; Song, C.; Wang, C.; Yu, Y.; Wang, G. Spinal Protein Kinase Mζ Regulates α-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic Acid Receptor Trafficking and Dendritic Spine Plasticity via Kalirin-7 in the Pathogenesis of Remifentanil-induced Postincisional Hyperalgesia in Rats. Anesthesiology 2018, 129, 173–186. [Google Scholar] [CrossRef]
- Wang, Z.; Tao, Y.; Song, C.; Liu, P.; Wang, C.; Li, Y.; Cui, W.; Xie, K.; Zhang, L.; Wang, G. Spinal hevin mediates membrane trafficking of GluA1-containing AMPA receptors in remifentanil-induced postoperative hyperalgesia in mice. Neurosci. Lett. 2020, 722, 134855. [Google Scholar] [CrossRef]
- Bottazzi, B.; Garlanda, C.; Salvatori, G.; Jeannin, P.; Manfredi, A.; Mantovani, A. Pentraxins as a key component of innate immunity. Curr. Opin. Immunol. 2006, 18, 10–15. [Google Scholar] [CrossRef]
- Zanier, E.R.; Brandi, G.; Peri, G.; Longhi, L.; Zoerle, T.; Tettamanti, M.; Garlanda, C.; Sigurta, A.; Valaperta, S.; Mantovani, A.; et al. Cerebrospinal fluid pentraxin 3 early after subarachnoid hemorrhage is associated with vasospasm. Intensive Care Med. 2011, 37, 302–309. [Google Scholar] [CrossRef]
- Rodriguez-Grande, B.; Swana, M.; Nguyen, L.; Englezou, P.; Maysami, S.; Allan, S.M.; Rothwell, N.J.; Garlanda, C.; Denes, A.; Pinteaux, E. The acute-phase protein PTX3 is an essential mediator of glial scar formation and resolution of brain edema after ischemic injury. J. Cereb. Blood Flow Metab. 2014, 34, 480–488. [Google Scholar] [CrossRef]
- Zhou, C.; Chen, H.; Zheng, J.F.; Guo, Z.D.; Huang, Z.J.; Wu, Y.; Zhong, J.J.; Sun, X.C.; Cheng, C.J. Pentraxin 3 contributes to neurogenesis after traumatic brain injury in mice. Neural Regen. Res. 2020, 15, 2318–2326. [Google Scholar]
- Fossati, G.; Pozzi, D.; Canzi, A.; Mirabella, F.; Valentino, S.; Morini, R.; Ghirardini, E.; Filipello, F.; Moretti, M.; Gotti, C.; et al. Pentraxin 3 regulates synaptic function by inducing AMPA receptor clustering via ECM remodeling and β1-integrin. EMBO J. 2019, 38, e99529. [Google Scholar] [CrossRef]
- Brügger-Andersen, T.; Pönitz, V.; Kontny, F.; Staines, H.; Grundt, H.; Sagara, M.; Nilsen, D.W. The long pentraxin 3 (PTX3): A novel prognostic inflammatory marker for mortality in acute chest pain. Thromb. Haemost. 2009, 102, 555–563. [Google Scholar] [CrossRef]
- Salcini, C.; Atasever-Arslan, B.; Sunter, G.; Gur, H.; Isik, F.B.; Saylan, C.C.; Yalcin, A.D. High Plasma Pentraxin 3 Levels in Diabetic Polyneuropathy Patients with Nociceptive Pain. Tohoku J. Exp. Med. 2016, 239, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Mizoguchi, A.; Nakanishi, H.; Kimura, K.; Matsubara, K.; Ozaki-Kuroda, K.; Katata, T.; Honda, T.; Kiyohara, Y.; Heo, K.; Higashi, M.; et al. Nectin: An adhesion molecule involved in formation of synapses. J. Cell Biol. 2002, 156, 555–565. [Google Scholar] [CrossRef]
- Gao, Y.; Hong, X.; Wang, H. Role of Nectin-1/c-Src Signaling in the Analgesic Effect of GDNF on a Rat Model of Chronic Constrictive Injury. J. Mol. Neurosci. 2016, 60, 258–266. [Google Scholar] [CrossRef]
- Matsushita, K.; Tozaki-Saitoh, H.; Kojima, C.; Masuda, T.; Tsuda, M.; Inoue, K.; Hoka, S. Chemokine (C-C motif) receptor 5 is an important pathological regulator in the development and maintenance of neuropathic pain. Anesthesiology 2014, 120, 1491–1503. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Qiu, X.; Deis, J.; Lin, T.; Chen, X. Pentraxin 3 deficiency exacerbates lipopolysaccharide-induced inflammation in adipose tissue. Int. J. Obes. 2020, 44, 525–538. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, L.; Wang, C.; Tang, X.; Chen, Y.; Wang, X.; Su, L.; Hu, N.; Xie, K.; Yu, Y.; et al. Sevoflurane-induced learning deficits and spine loss via nectin-1/corticotrophin- releasing hormone receptor type 1 signaling. Brain Res. 2019, 1710, 188–198. [Google Scholar] [CrossRef]
- Zhang, L.; Terrando, N.; Xu, Z.Z.; Bang, S.; Jordt, S.E.; Maixner, W.; Serhan, C.N.; Ji, R.R. Distinct Analgesic Actions of DHA and DHA-Derived Specialized Pro-Resolving Mediators on Post-operative Pain after Bone Fracture in Mice. Front. Pharmacol. 2018, 9, 412. [Google Scholar] [CrossRef] [Green Version]
- Bang, S.; Xie, Y.K.; Zhang, Z.J.; Wang, Z.; Xu, Z.Z.; Ji, R.R. GPR37 regulates macrophage phagocytosis and resolution of inflammatory pain. J. Clin. Investig. 2018, 128, 3568–3582. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Peirs, C.; Seal, R.P. Neural circuits for pain: Recent advances and current views. Science 2016, 354, 578–584. [Google Scholar] [CrossRef]
- Zhang, L.; Shu, R.; Wang, H.; Yu, Y.; Wang, C.; Yang, M.; Wang, M.; Wang, G. Hydrogen-rich saline prevents remifentanil-induced hyperalgesia and inhibits MnSOD nitration via regulation of NR2B-containing NMDA receptor in rats. Neuroscience 2014, 280, 171–180. [Google Scholar] [CrossRef]
- Chen, G.; Luo, X.; Qadri, M.Y.; Berta, T.; Ji, R.R. Sex-Dependent Glial Signaling in Pathological Pain: Distinct Roles of Spinal Microglia and Astrocytes. Neurosci. Bull. 2018, 34, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Sorge, R.E.; Mapplebeck, J.C.; Rosen, S.; Beggs, S.; Taves, S.; Alexander, J.K.; Martin, L.J.; Austin, J.S.; Sotocinal, S.G.; Chen, D.; et al. Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nat. Neurosci. 2015, 18, 1081–1083. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Yu, H.; Sun, Y.; Yu, W. Pentraxin-3 in the Spinal Dorsal Horn Upregulates Nectin-1 Expression in Neuropathic Pain after Spinal Nerve Damage in Male Mice. Brain Sci. 2022, 12, 648. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050648
Zhu M, Yu H, Sun Y, Yu W. Pentraxin-3 in the Spinal Dorsal Horn Upregulates Nectin-1 Expression in Neuropathic Pain after Spinal Nerve Damage in Male Mice. Brain Sciences. 2022; 12(5):648. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050648
Chicago/Turabian StyleZhu, Min, Hongli Yu, Ying Sun, and Wenli Yu. 2022. "Pentraxin-3 in the Spinal Dorsal Horn Upregulates Nectin-1 Expression in Neuropathic Pain after Spinal Nerve Damage in Male Mice" Brain Sciences 12, no. 5: 648. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050648