
Quantum Mechanical Aspects in the Pathophysiology of Neuropathic Pain


 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Mathematical Model
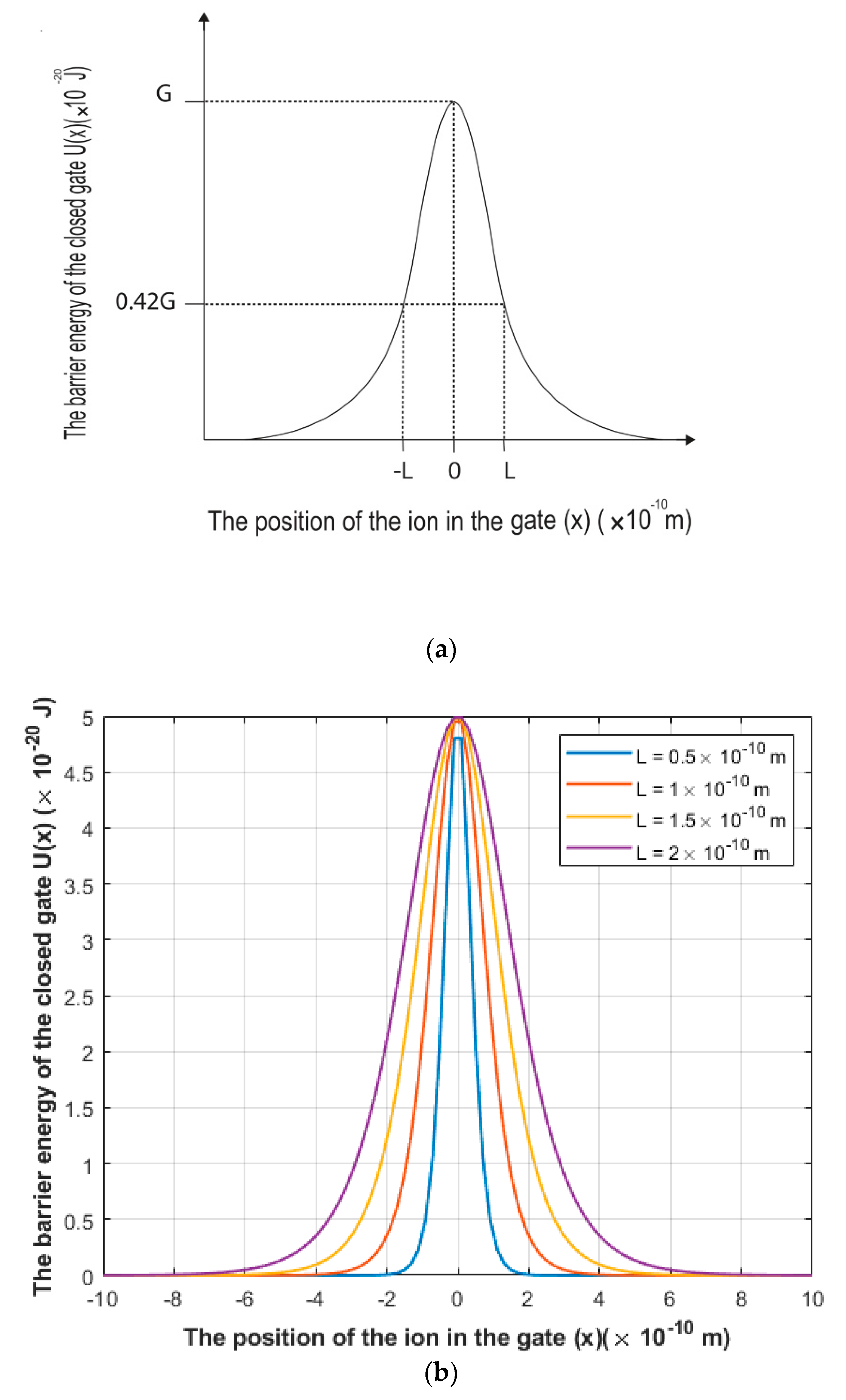
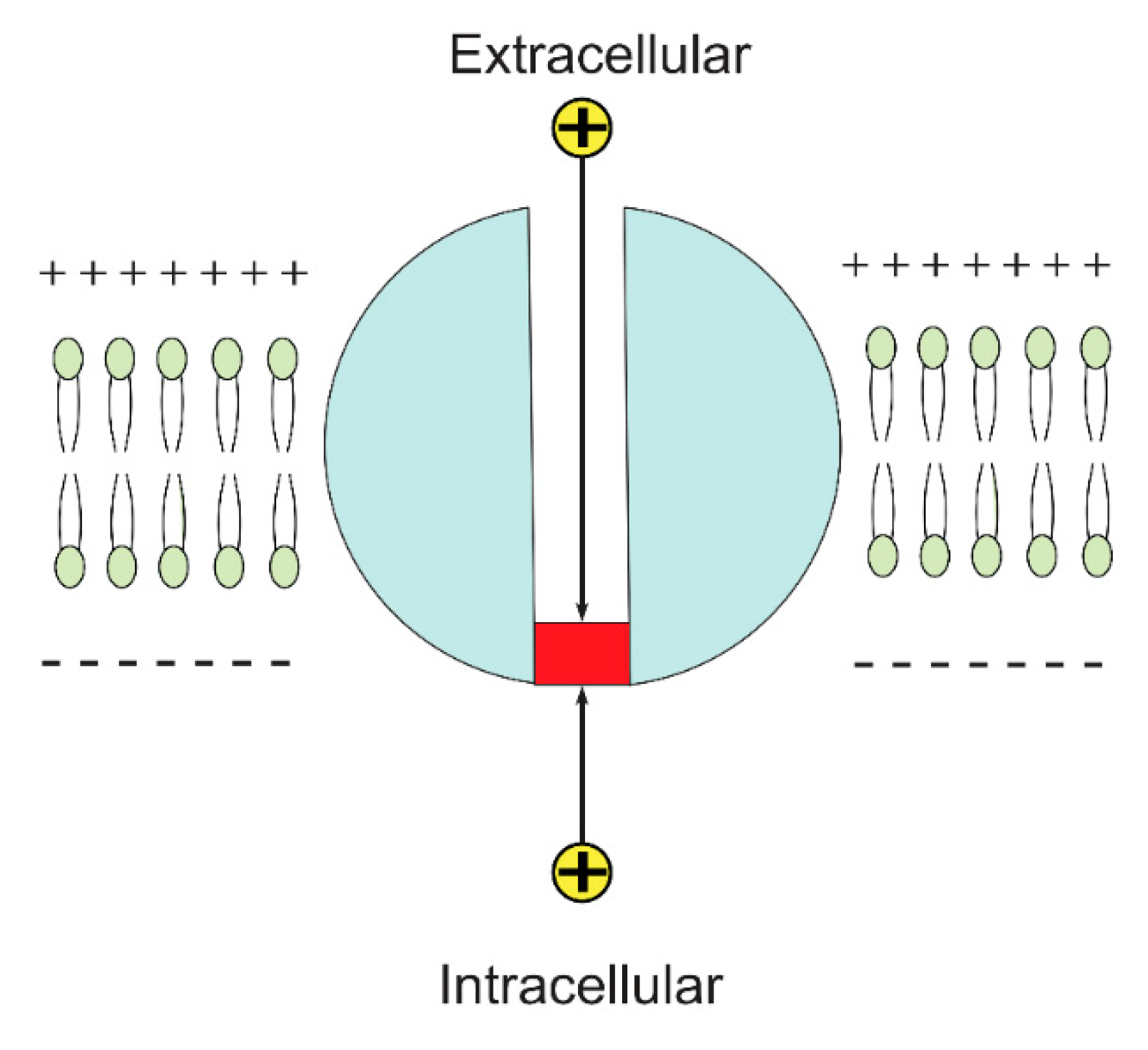
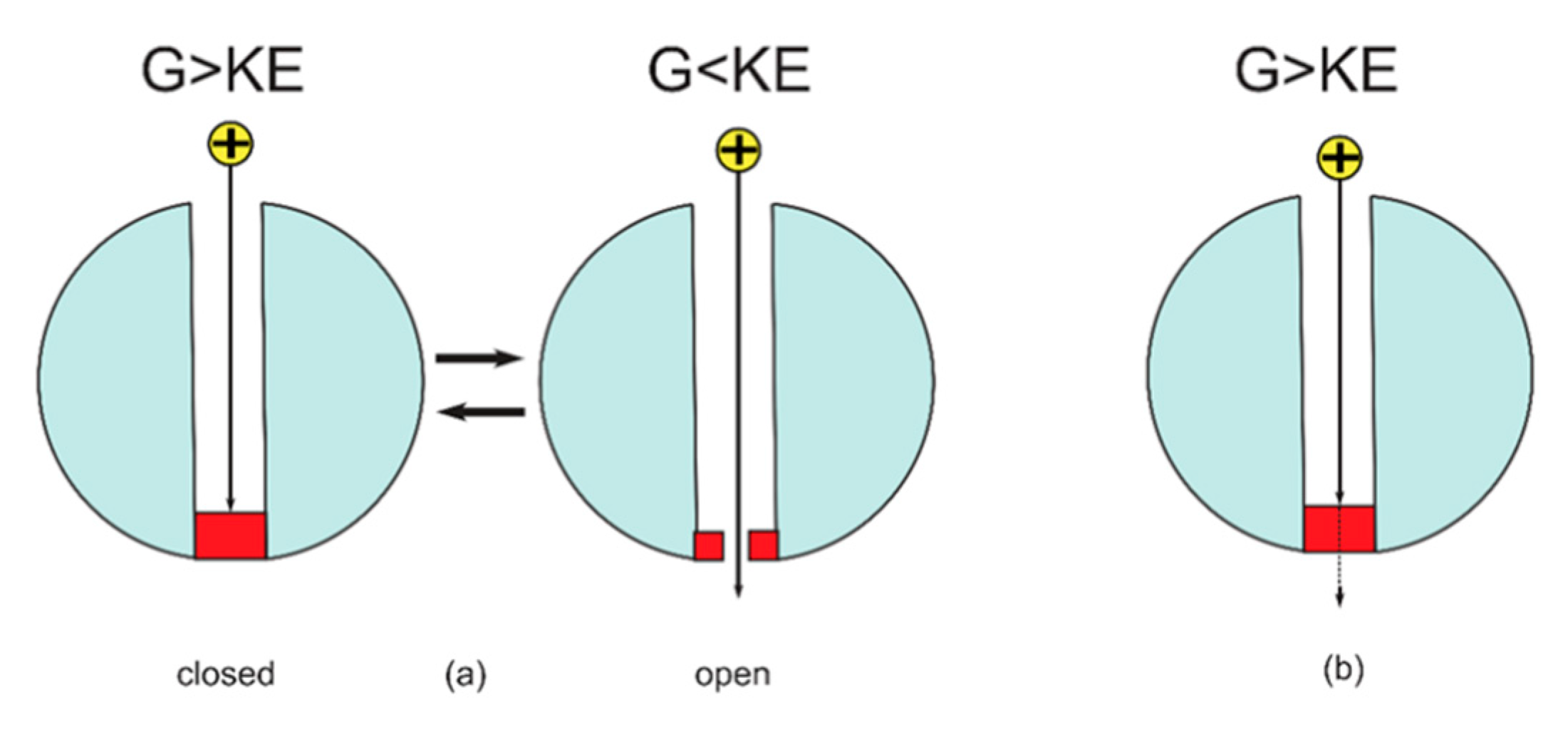
2.1. The Quantum Tunneling of Ions and the Quantum Conductance
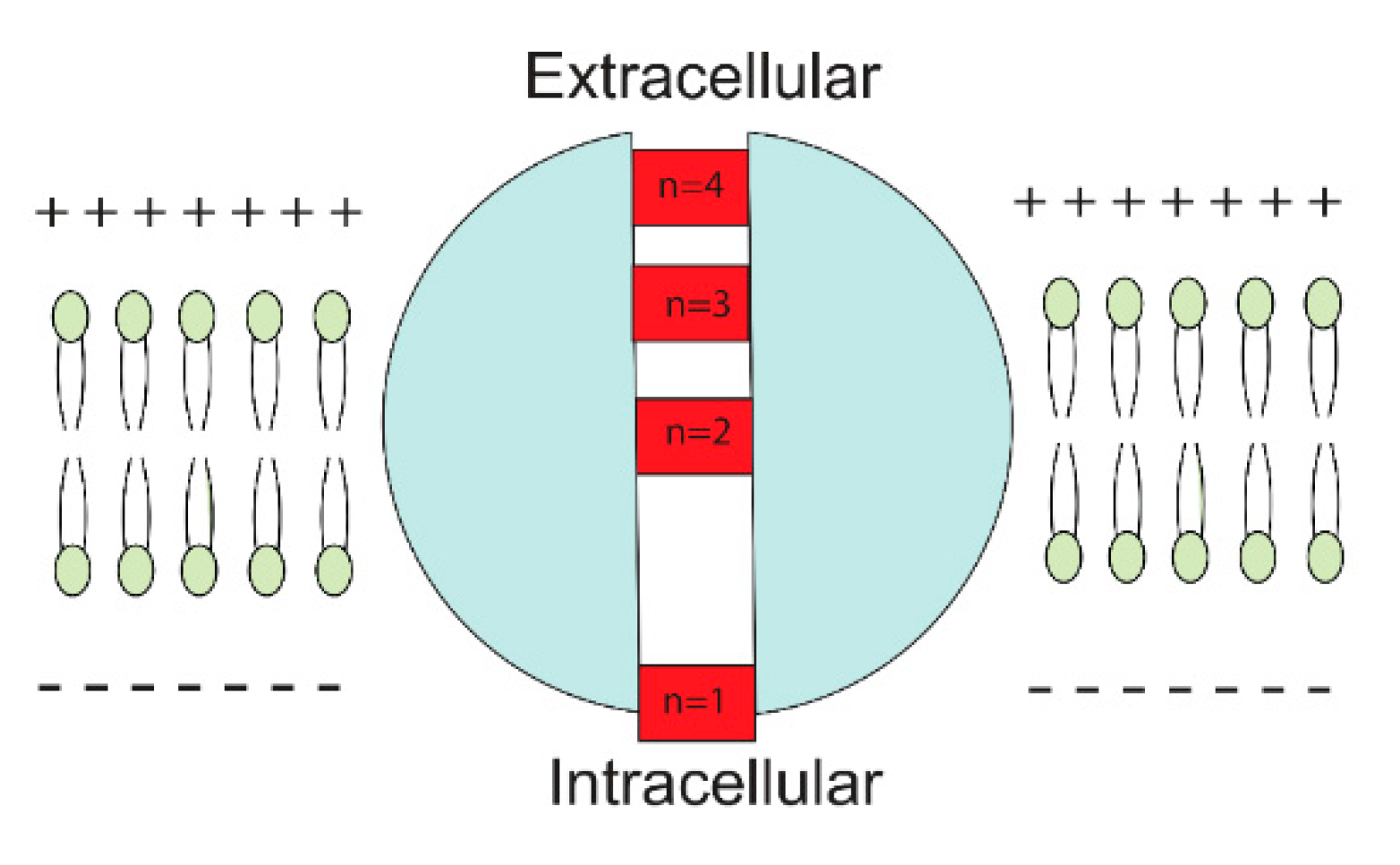
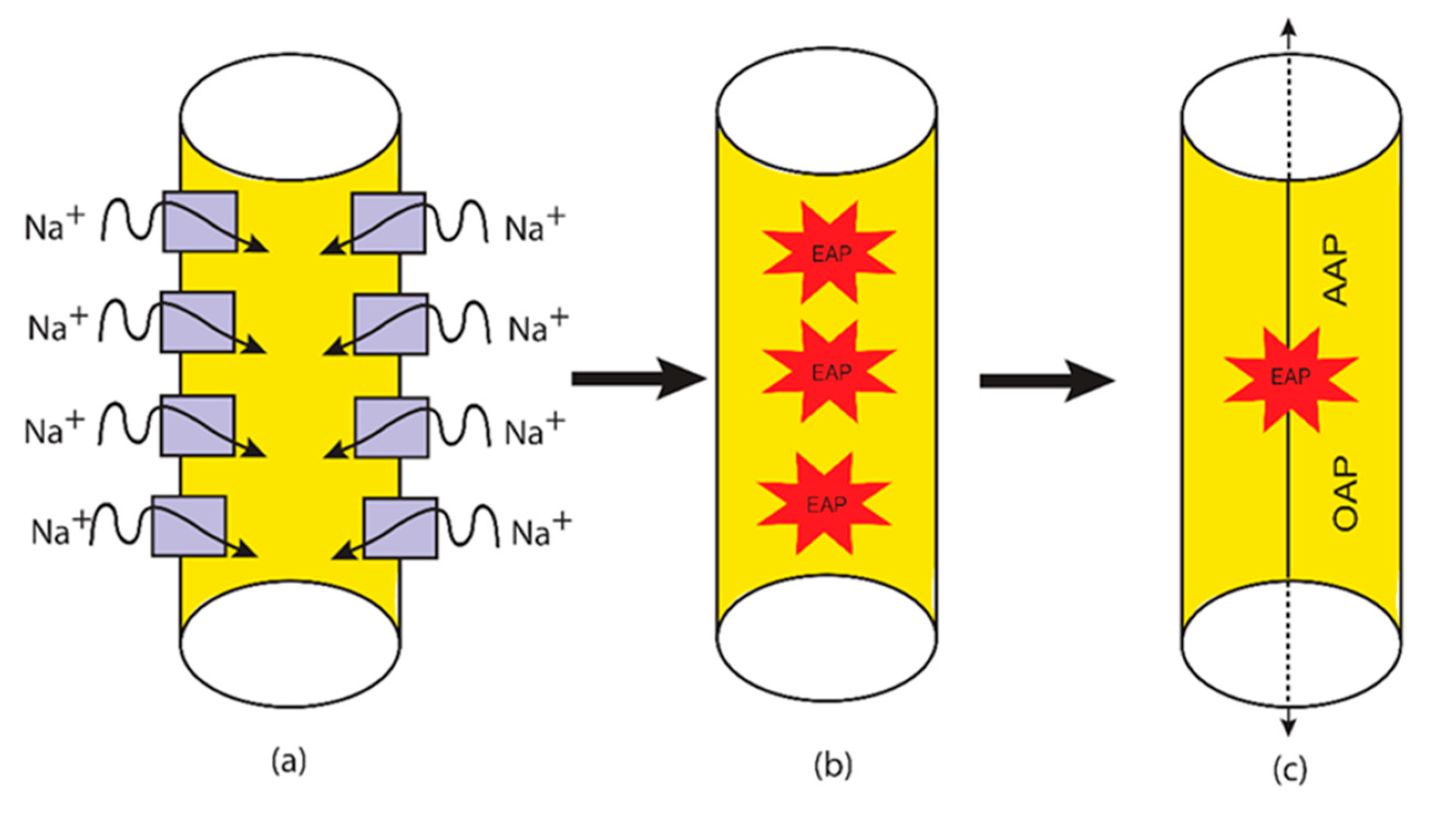
2.2. The Quantum Tunneling-Induced Membrane Depolarization and The Quantum Tunneling Current
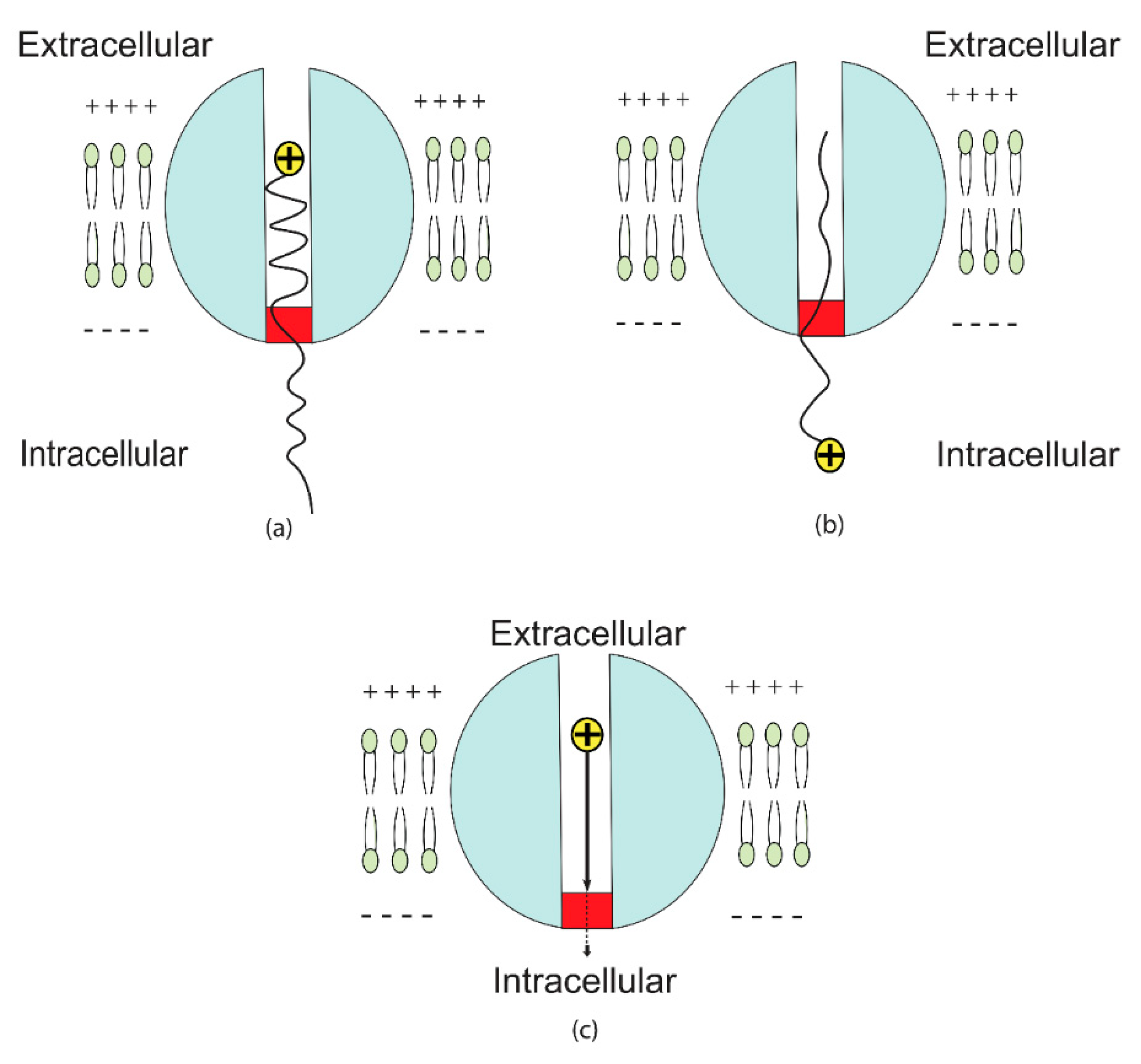
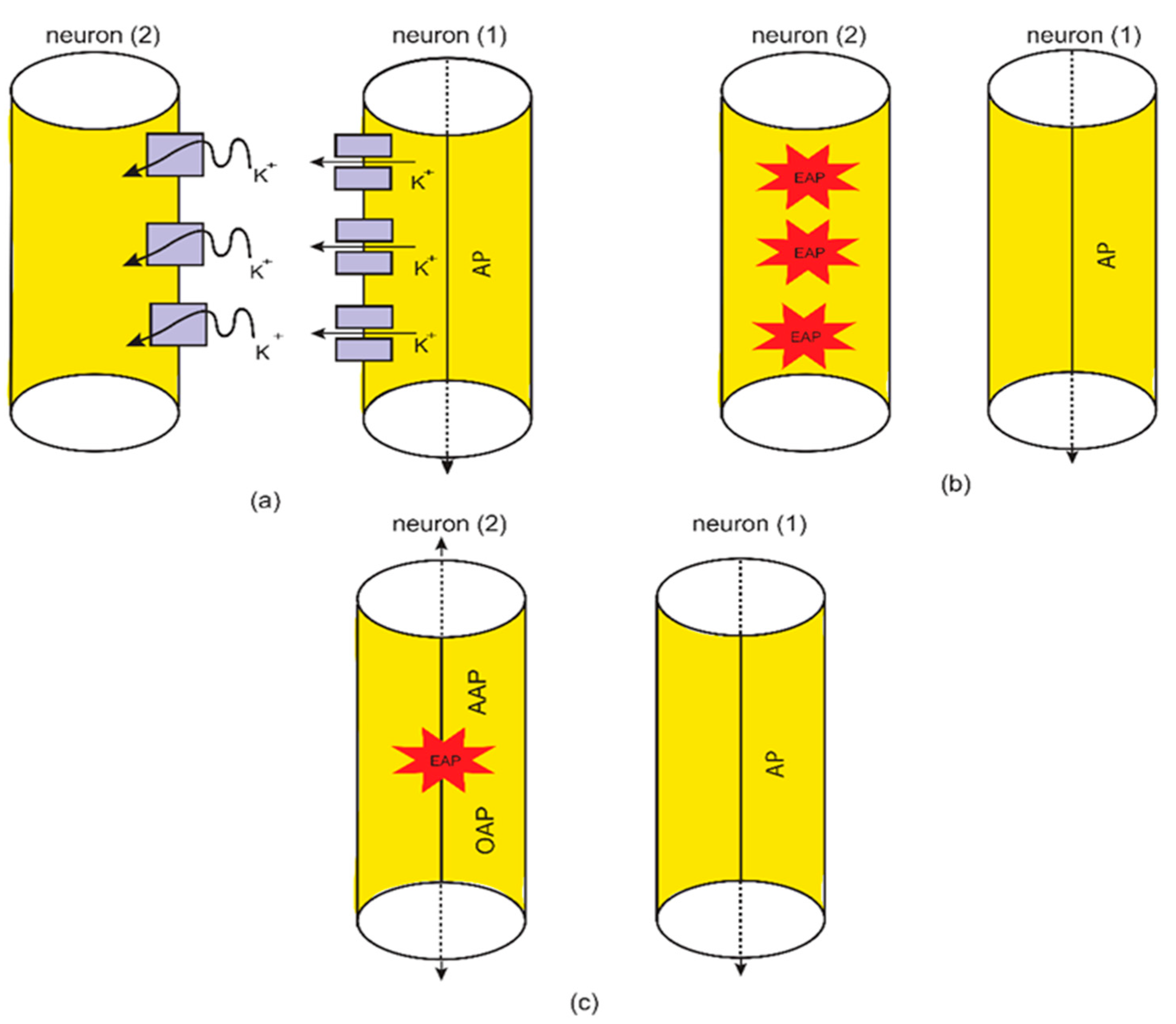
2.3. The Formation of Quantum Synapses between The Axons as a Mechanism for Ephaptic Coupling
2.4. The Quantum Tunneling-Induced Spontaneous Neuronal Firing
3. Results
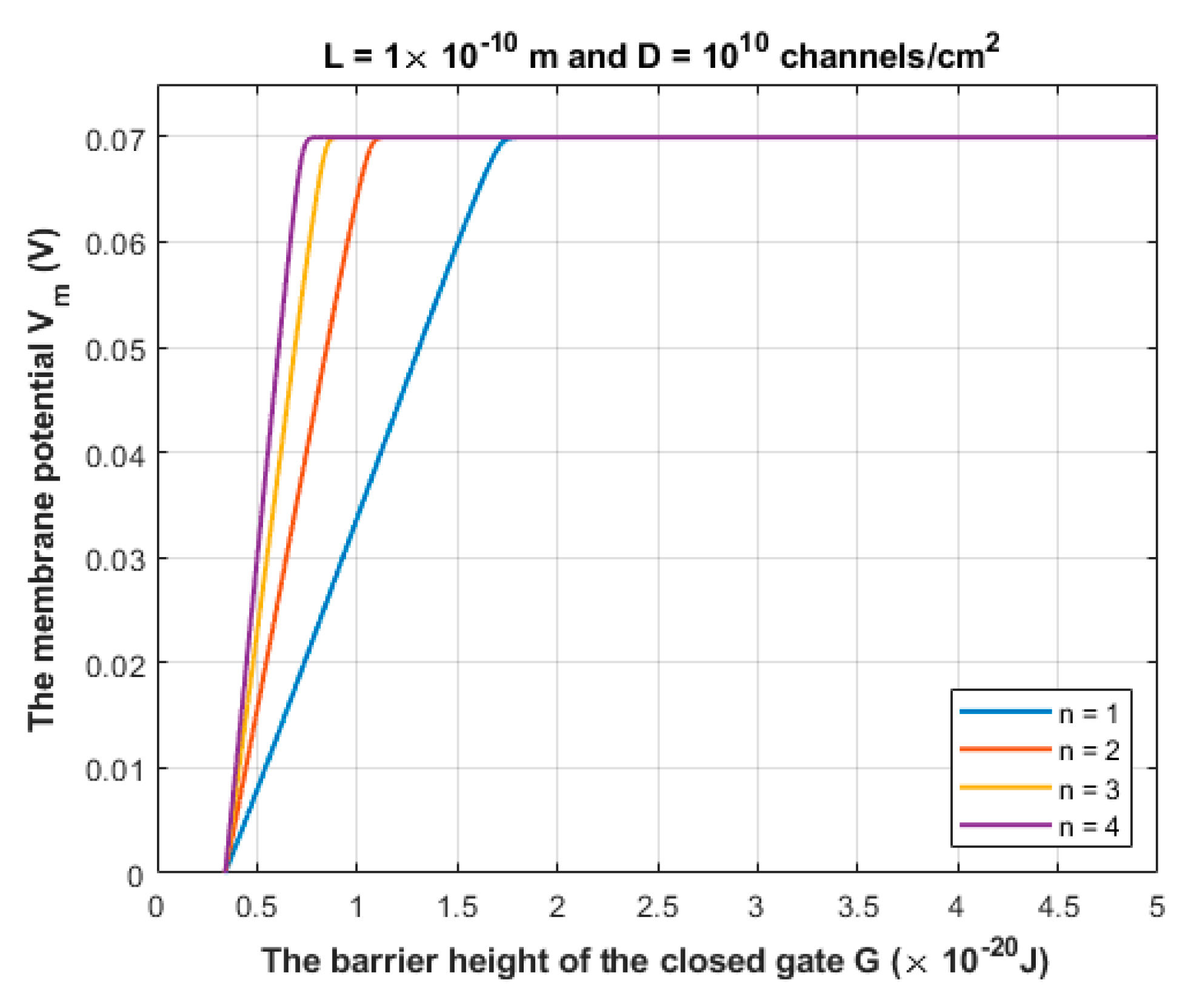
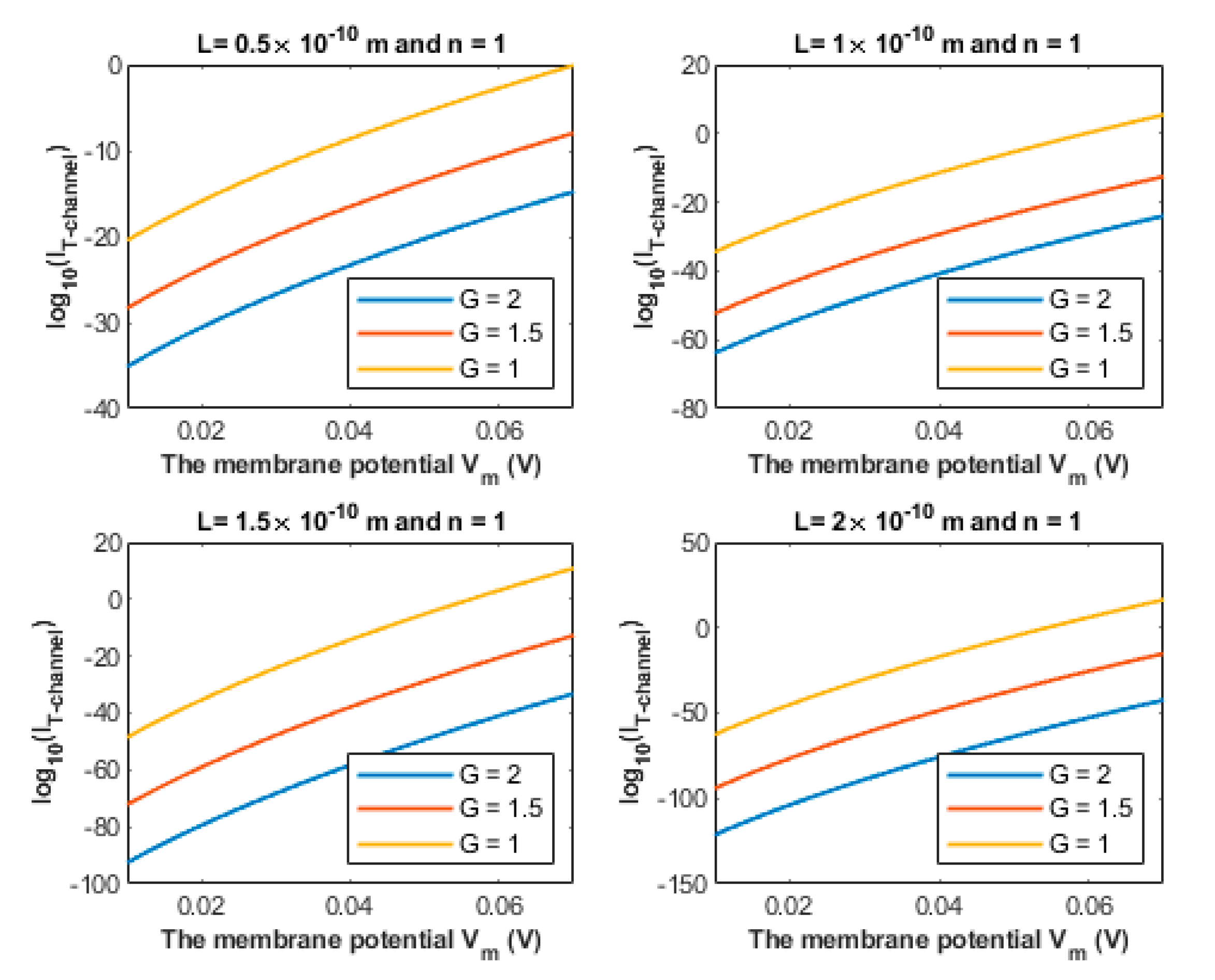
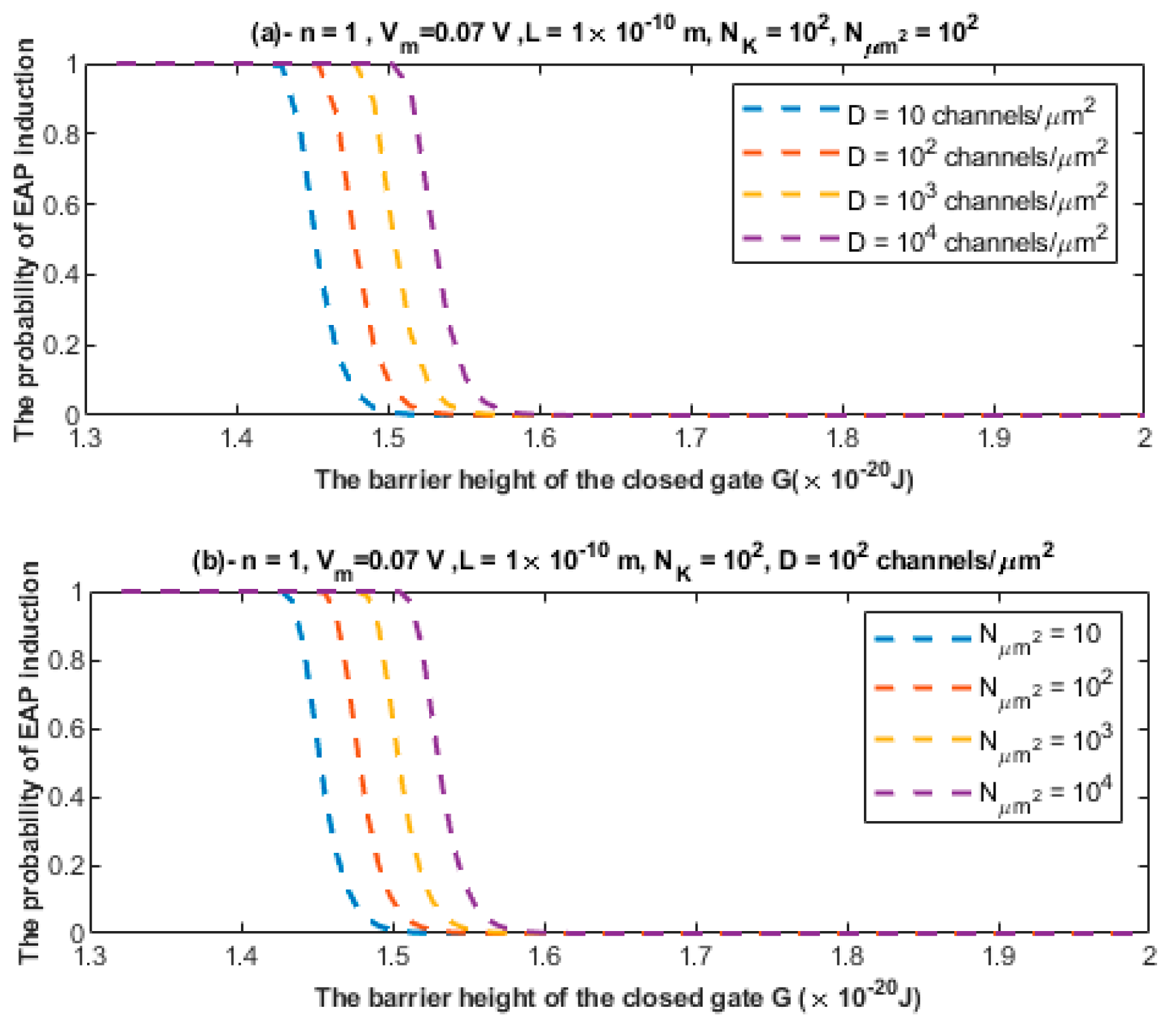
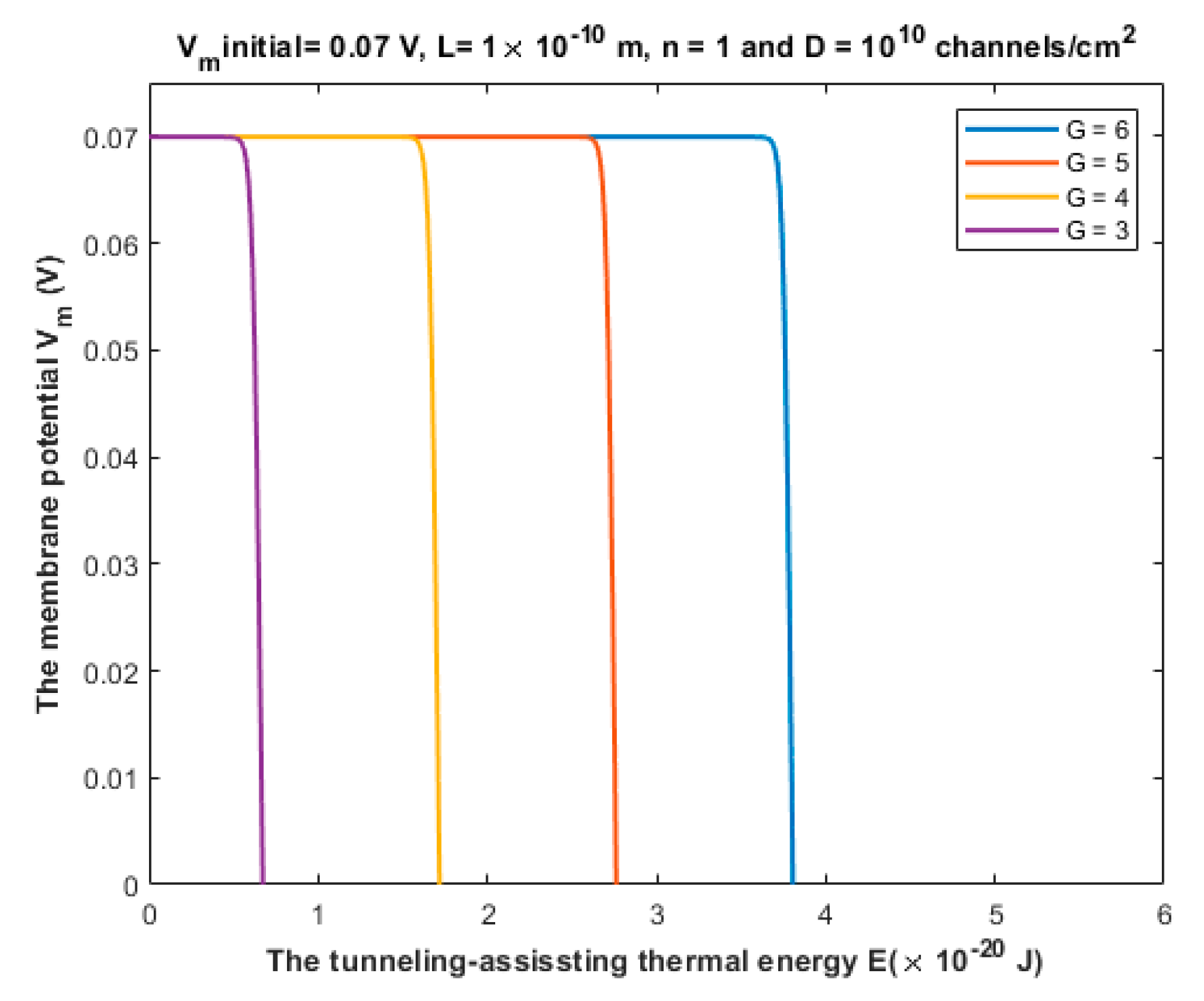
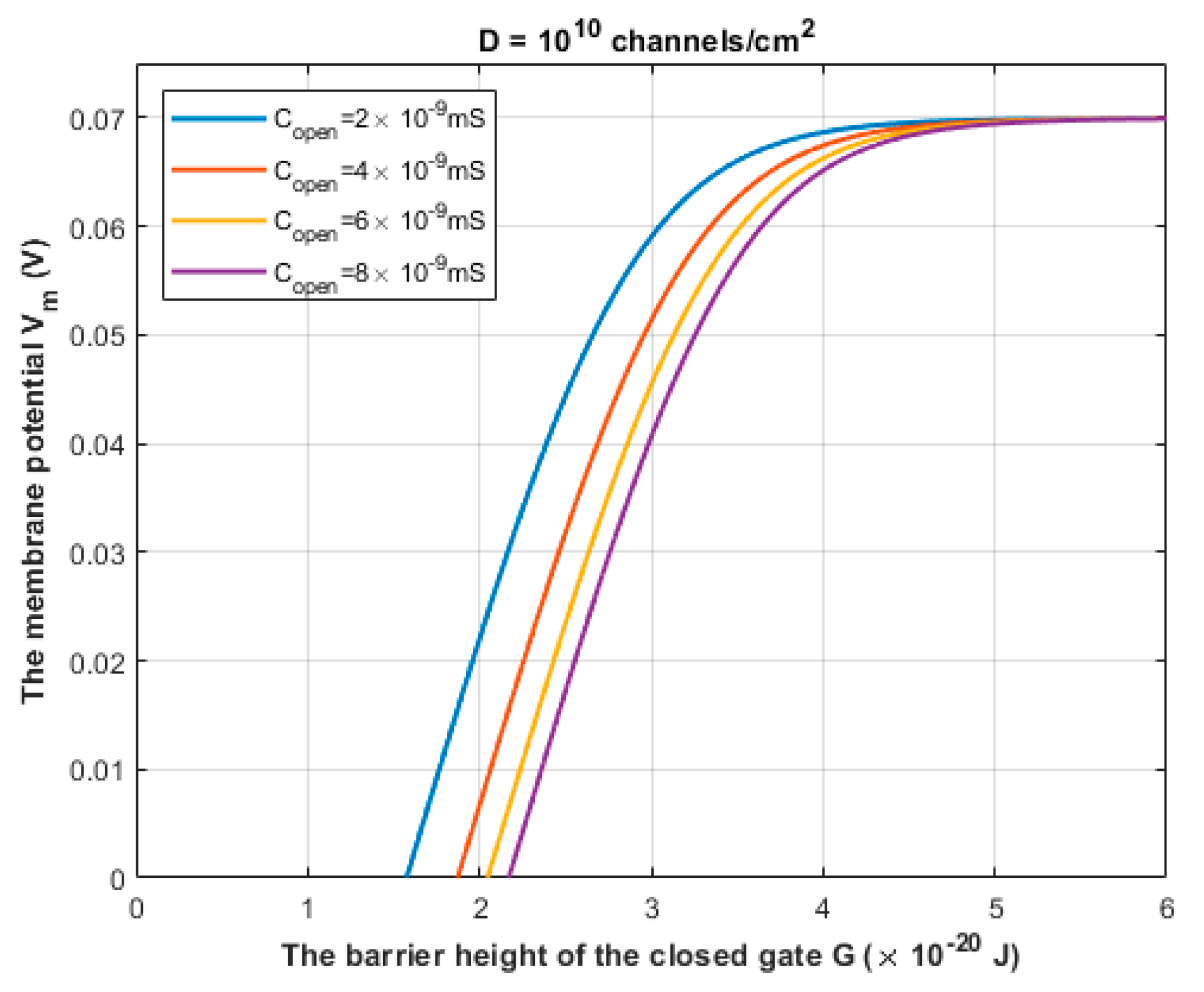
3.1. The Quantum Tunneling-Induced Membrane Depolarization and The Quantum Tunneling Currents
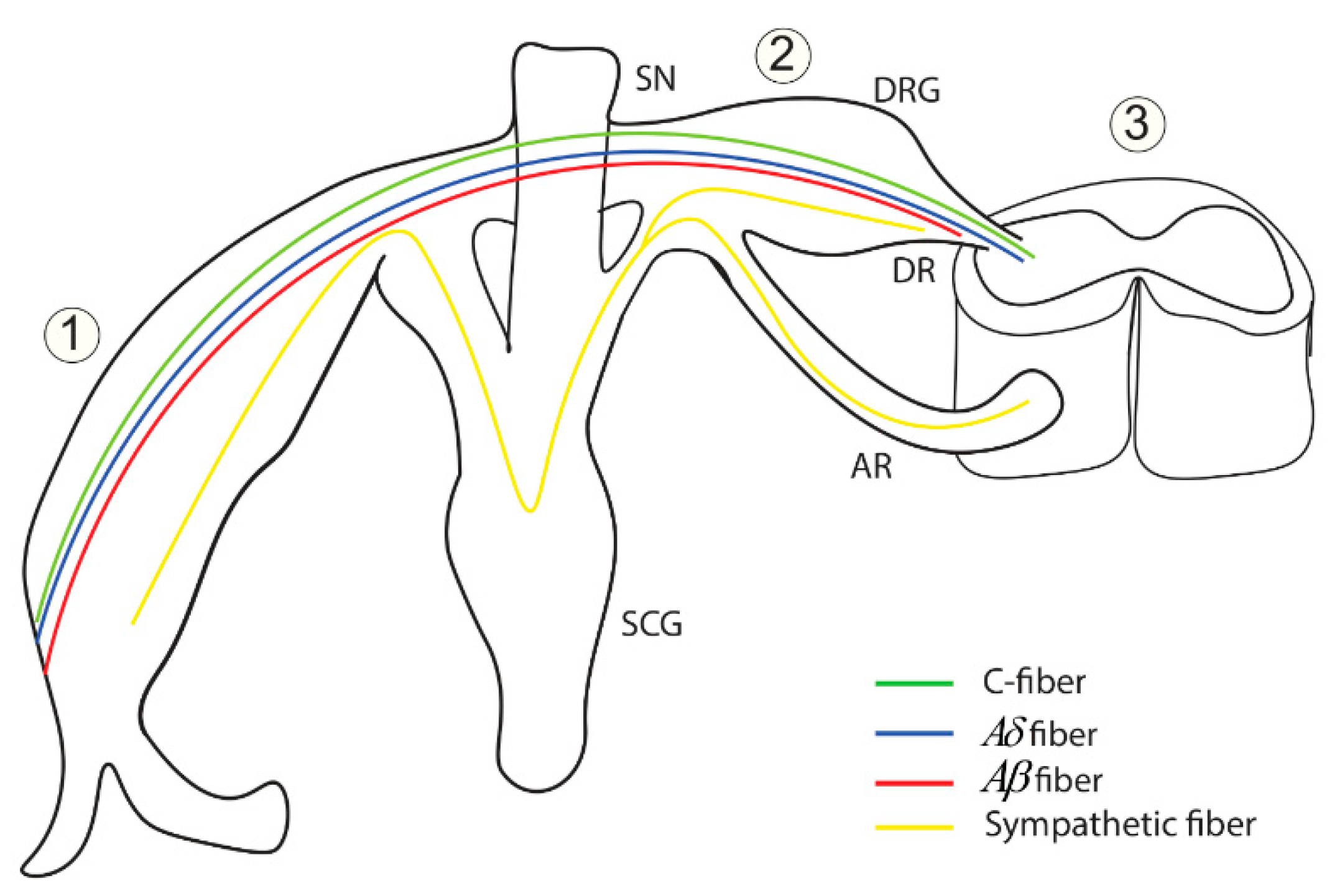
3.2. The Quantum Ephaptic Coupling or ‘Quantum Synapse’
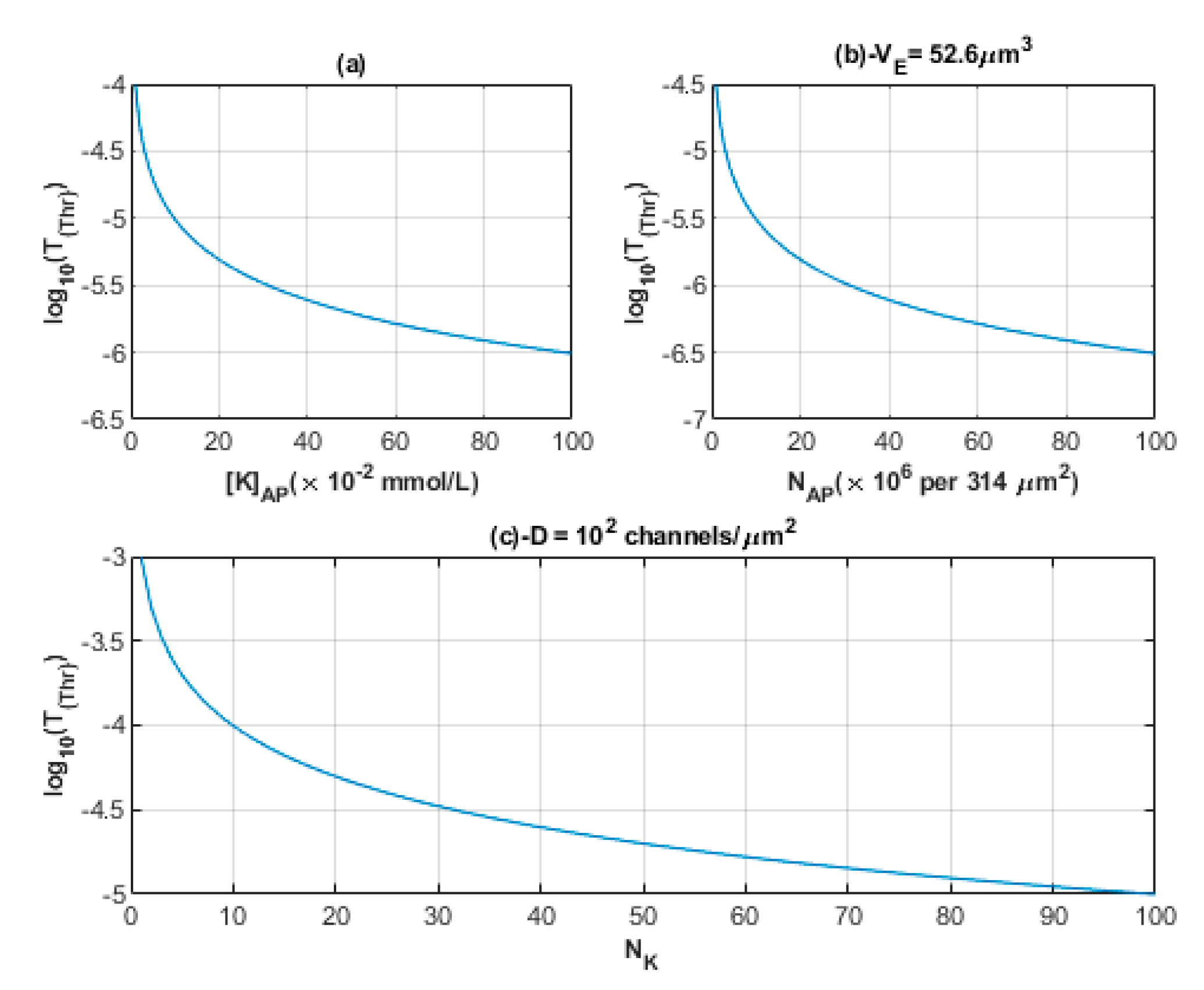
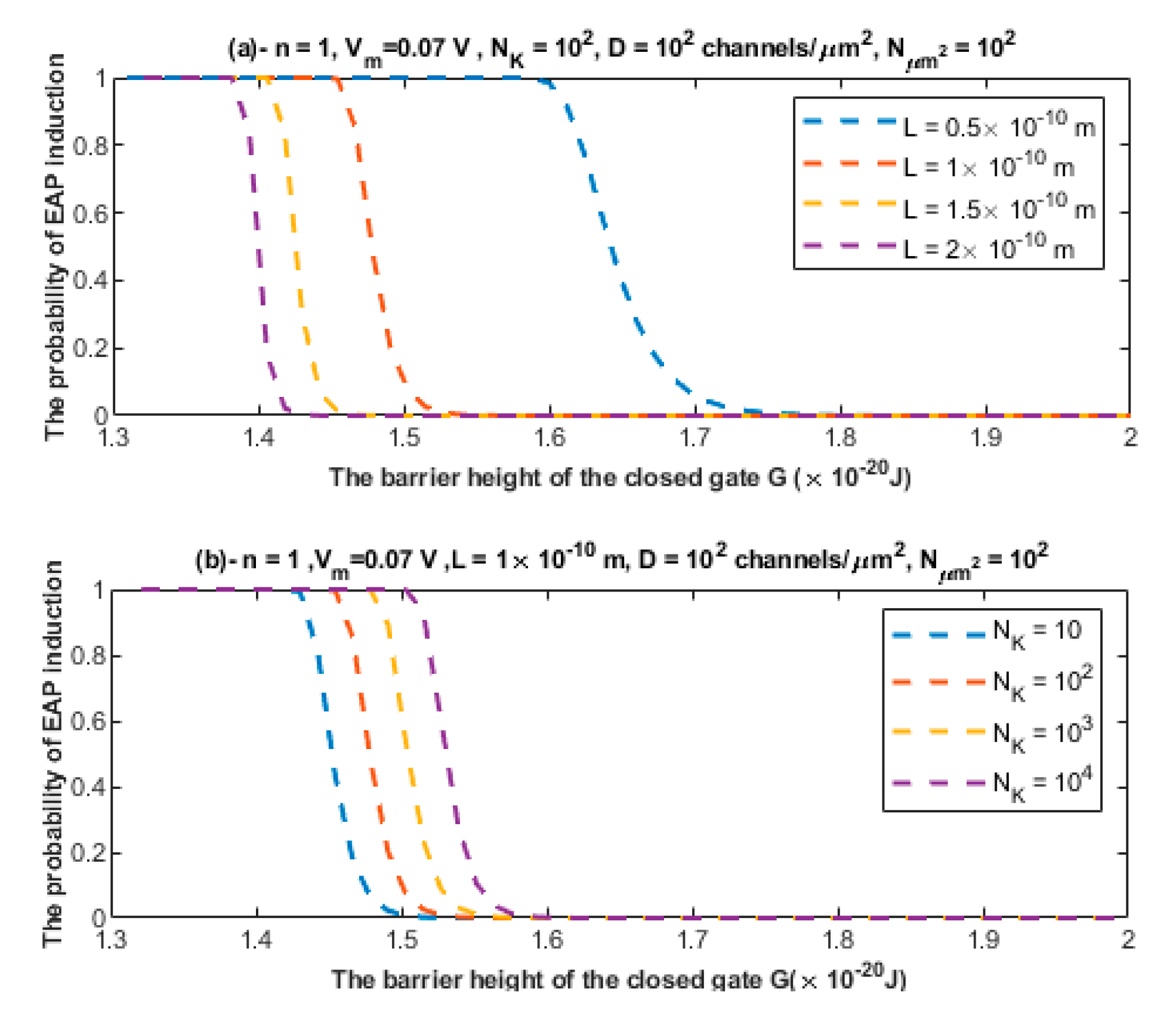
3.3. The Quantum Tunneling-Induced Spontaneous Neuronal Firing
4. Discussion
4.1. Elaboration of The Ion Quantum Tunneling Model in The Context of Neuropathic Pain
4.2. The Potential Applicability of The Quantum Tunneling Model in The Context of Neuropathic Pain
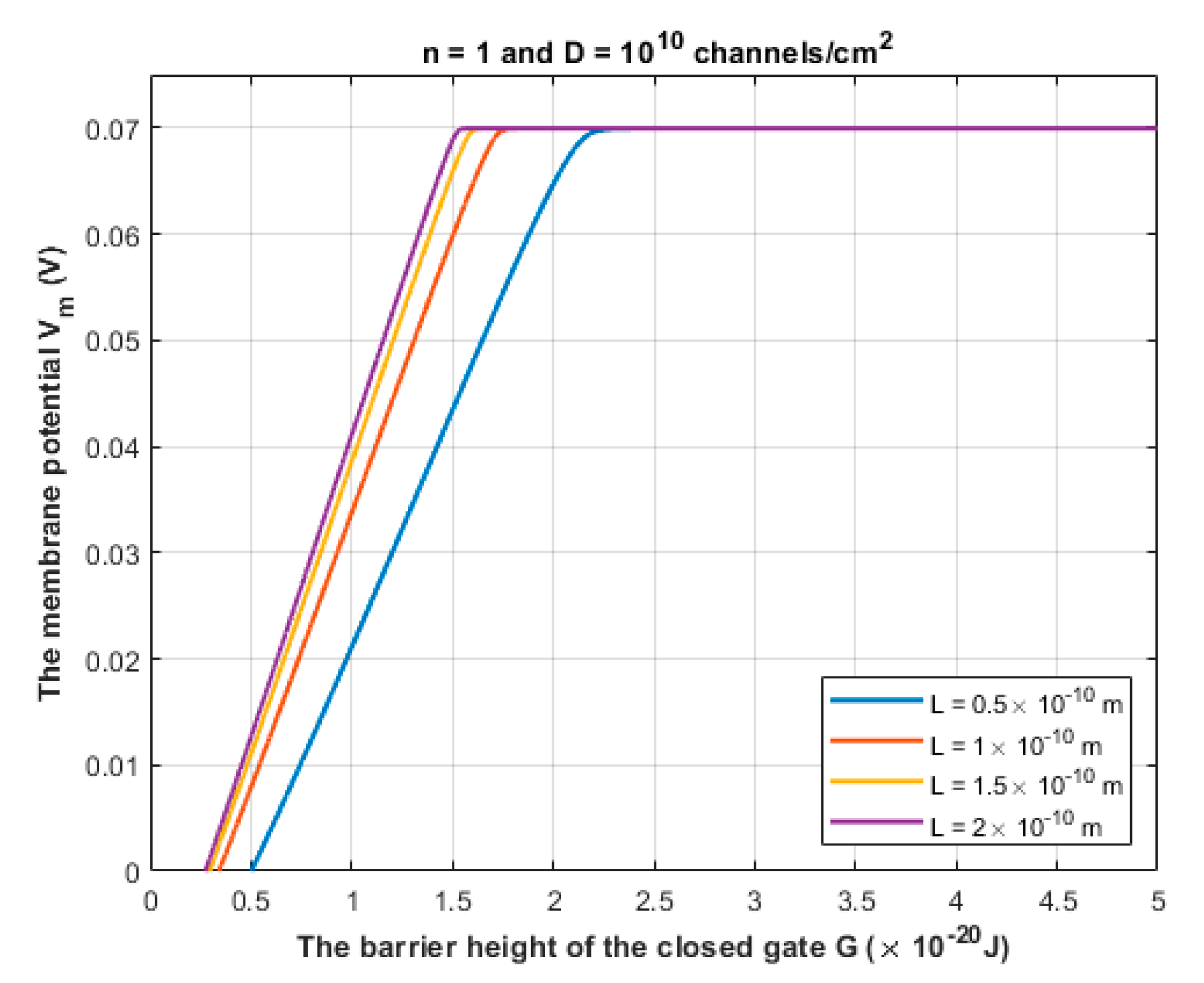
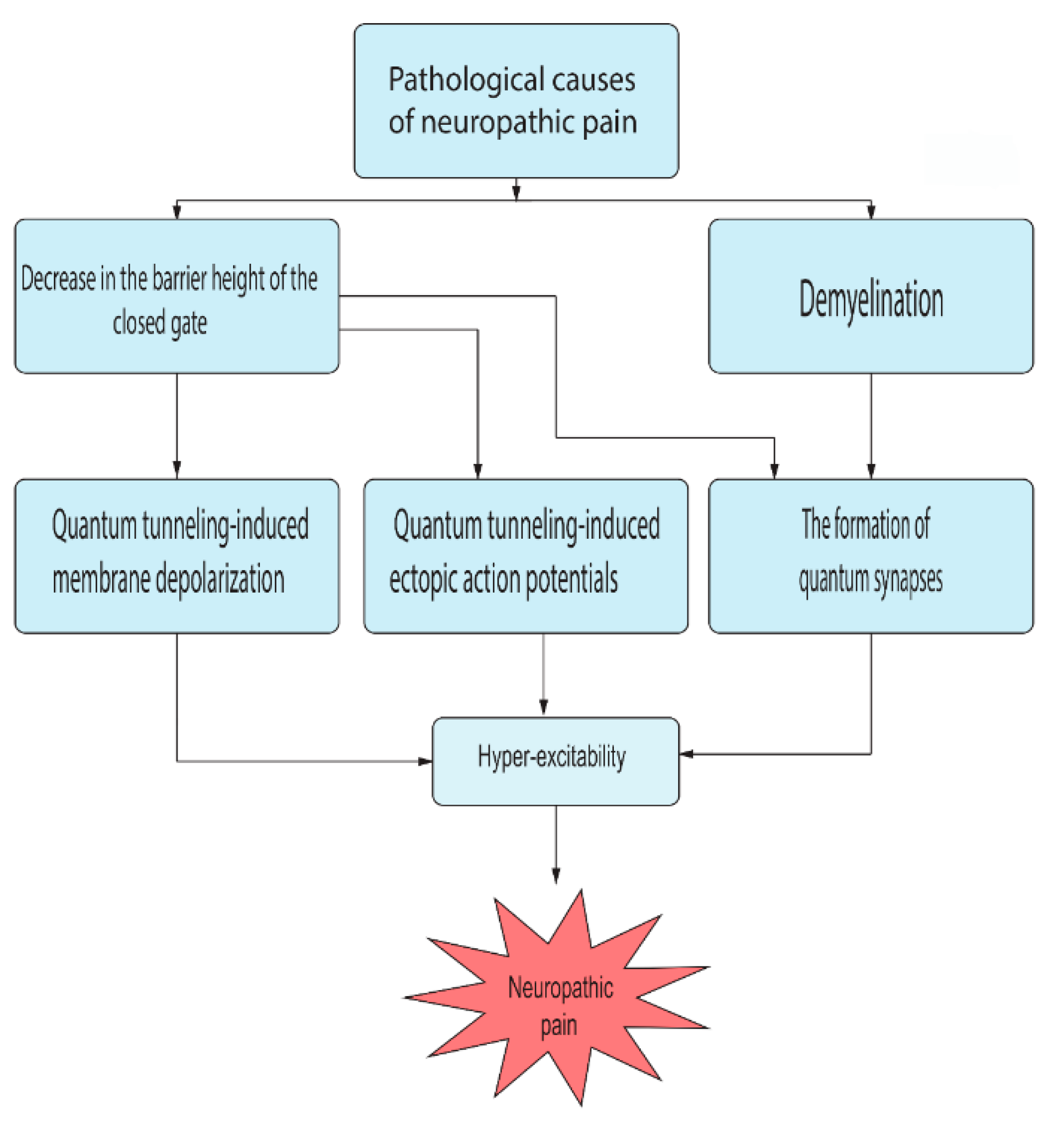
- Pathophysiological implications: There are certain unique features in the quantum tunneling model that make it distinctive from any classical model. These features can be deduced from Equation (2) and include: (1) The exponential dependence on the mass of the ion and the length of the gate to determine the quantum tunneling probability and quantum conductance. This would serve to be a promising strategy to test the validity of the role of the quantum behavior of ions. For example, as sodium and potassium ions have different masses, then observing an exponential difference between ions in terms of tunneling probability and quantum conductance is expected. Interestingly, the exponential mass difference can be also applied on other ions such as lithium and hydrogen ions. Similarly, observing an exponential dependence on the length of the gate will add additional supporting evidence. (2) Another implication that indicates strongly to the quantum tunneling behavior of ions is observing a depolarization action by potassium ions especially when there is a decrease in the energy barrier of the closed gate. Classically, when there is a gain-of-function mutation in potassium channels or when these channels open, it is expected that the outward potassium current will occur, which tends to hyperpolarize the membrane potential. However, according to the quantum tunneling model, it is expected that the inward potassium current will occur, which tends to depolarize the membrane potential.
- Pharmacological implications: These implications are crucial to be demonstrated to exhibit the beneficial consequences of the quantum tunneling model. If the pathophysiological implications can be tested experimentally and the quantum coherence of ions can be proven in ion channels, especially in the narrow hydrophobic gate, then we can propose this class of medications, which is ‘quantum decoherence inducers’ or ‘quantum coherence destroyers’ or ‘quantum decoherence agents’. All of these coined terms can be used to describe the ability of these proposed drugs to collapse the quantum wave or to weaken the quantum behavior of ions. If this quantum decoherence happens, then all the proposed pathophysiological mechanisms will be diminished or will be eliminated. Consequently, these proposed drugs can contribute significantly to pain relief and achieve satisfactory clinical outcomes. Our proposal for these drugs requires further investigation and an interdisciplinary cooperation to test the potential applicability of this proposal. We propose these drugs to attract the attention of all researchers across the different related disciplines to the possible applicability of the quantum tunneling model to act actively in the treatment of the neuropathic pain or even in the prevention of neuropathic pain by implementing our understanding of the pathophysiological mechanisms from the quantum mechanical perspective.
4.3. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B.; et al. Neuropathic pain. Nat. Rev. Dis. Primers 2017, 3, 17002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapunar, D.; Ljubkovic, M.; Lirk, P.; McCallum, J.B.; Hogan, Q.H. Distinct Membrane Effects of Spinal Nerve Ligation on Injured and Adjacent Dorsal Root Ganglion Neurons in Rats. Anesthesiology 2005, 103, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lee, C.H.; Oh, U. Painful channels in sensory neurons. Mol. Cells 2005, 20, 315–324. [Google Scholar]
- Wu, B.; Su, X.; Zhang, W.; Zhang, Y.-H.; Feng, X.; Ji, Y.-H.; Tan, Z.-Y. Oxaliplatin Depolarizes the IB4– Dorsal Root Ganglion Neurons to Drive the Development of Neuropathic Pain through TRPM8 in Mice. Front. Mol. Neurosci. 2021, 14, 690858. [Google Scholar] [CrossRef]
- Coggan, J.S.; Ocker, G.K.; Sejnowski, T.J.; Prescott, S.A. Explaining pathological changes in axonal excitability through dynamical analysis of conductance-based models. J. Neural Eng. 2011, 8, 065002. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Strong, J.; Meij, J.; Zhang, J.-M.; Yu, L. Neuropathic pain: Early spontaneous afferent activity is the trigger. Pain 2005, 116, 243–256. [Google Scholar] [CrossRef]
- Djouhri, L.; Koutsikou, S.; Fang, X.; McMullan, S.; Lawson, S.N. Spontaneous Pain, Both Neuropathic and Inflammatory, Is Related to Frequency of Spontaneous Firing in Intact C-Fiber Nociceptors. J. Neurosci. 2006, 26, 1281–1292. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.N.; Meyer, R.A. Mechanisms of neuropathic pain. Neuron 2006, 52, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Nickel, F.T.; Seifert, F.; Lanz, S.; Maihöfner, C. Mechanisms of neuropathic pain. Eur. Neuropsychopharmacol. 2012, 22, 81–91. [Google Scholar] [CrossRef]
- Seltzer, Z.; Devor, M. Ephaptic transmission in chronically damaged peripheral nerves. Neurology 1979, 29, 1061. [Google Scholar] [CrossRef]
- Sorkin, L.S.; Eddinger, K.A.; Woller, S.A.; Yaksh, T.L. Origins of antidromic activity in sensory afferent fibers and neurogenic inflammation. In Seminars in Immunopathology; Springer: Berlin/Heidelberg, Germany, 2018; Volume 40, pp. 237–247. [Google Scholar]
- Asmedi, A.; Wibowo, S.; Meliala, L. Ephaptic crosstalk in Painful Diabetic Neuropathy: An electrodiagnostic study. J. Med Sci. (Berk. Ilmu Kedokt.) 2018, 50, 173–179. [Google Scholar] [CrossRef]
- Gould, H.J.; Soignier, R.D.; Cho, S.R.; Hernandez, C.; Diamond, I.; Taylor, B.K.; Paul, D. Ranolazine attenuates mechanical allodynia associated with demyelination injury. Pain Med. 2014, 15, 1771–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Bertagna, F.; D’Souza, E.M.; Heyes, D.J.; Johannissen, L.O.; Nery, E.T.; Pantelias, A.; Jimenez, A.S.-P.; Slocombe, L.; Spencer, M.G.; et al. Quantum Biology: An Update and Perspective. Quantum Rep. 2021, 3, 80–126. [Google Scholar] [CrossRef]
- Calvillo, L.; Redaelli, V.; Ludwig, N.; Qaswal, A.B.; Ghidoni, A.; Faini, A.; Rosa, D.; Lombardi, C.; Pengo, M.; Bossolasco, P.; et al. Quantum Biology Research Meets Pathophysiology and Therapeutic Mechanisms: A Biomedical Perspective. Quantum Rep. 2022, 4, 148–172. [Google Scholar] [CrossRef]
- Barjas Qaswal, A. Quantum Tunneling of Ions through the Closed Voltage-Gated Channels of the Biological Membrane: A Mathematical Model and Implications. Quantum Rep. 2019, 1, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Qaswal, A.B.; Ababneh, O.; Khreesha, L.; Al-Ani, A.; Suleihat, A.; Abbad, M. Mathematical Modeling of Ion Quantum Tunneling Reveals Novel Properties of Voltage-Gated Channels and Quantum Aspects of Their Pathophysiology in Excitability-Related Disorders. Pathophysiology 2021, 28, 116–154. [Google Scholar] [CrossRef]
- Qaswal, A. A Theoretical Study to Explain the Referred Pain Phenomenon and Its Characteristics via Quantum Tunneling of Potassium Ions through the Channels of Neuronal Membrane. NeuroQuantology 2019, 17, 43–52. [Google Scholar] [CrossRef]
- Qaswal, A.B. The Myelin Sheath Maintains the Spatiotemporal Fidelity of Action Potentials by Eliminating the Effect of Quantum Tunneling of Potassium Ions through the Closed Channels of the Neuronal Membrane. Quantum Rep. 2019, 1, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Alrabayah, M.; Qaswal, A.B.; Suleiman, A.; Khreesha, L. Role of Potassium Ions Quantum Tunneling in the Pathophysiology of Phantom Limb Pain. Brain Sci. 2020, 10, 241. [Google Scholar] [CrossRef] [Green Version]
- Balzani, E.; Fanelli, A.; Malafoglia, V.; Tenti, M.; Ilari, S.; Corraro, A.; Muscoli, C.; Raffaeli, W. A Review of the Clinical and Therapeutic Implications of Neuropathic Pain. Biomedicines 2021, 9, 1239. [Google Scholar] [CrossRef]
- Bridges, D.; Thompson, S.W.; Rice, A.S. Mechanisms of neuropathic pain. Br. J. Anaesth. 2001, 87, 12–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryal, P.; Sansom, M.S.; Tucker, S.J. Hydrophobic Gating in Ion Channels. J. Mol. Biol. 2014, 427, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oelstrom, K.; Goldschen-Ohm, M.P.; Holmgren, M.; Chanda, B. Evolutionarily conserved intracellular gate of voltage-dependent sodium channels. Nat. Commun. 2014, 5, 3420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labro, A.J.; Snyders, D.J. Being Flexible: The Voltage-Controllable Activation Gate of Kv Channels. Front. Pharmacol. 2012, 3, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hering, S.; Zangerl-Plessl, E.M.; Beyl, S.; Hohaus, A.; Andranovits, S.; Timin, E.N. Calcium channel gating. Pflügers Arch.-Eur. J. Physiol. 2018, 470, 1291–1309. [Google Scholar] [CrossRef] [Green Version]
- Chandra, A.K. Introductory Quantum Chemistry; Tata McGraw-Hill Education: New York, NY, USA, 1994. [Google Scholar]
- Eckart, C. The Penetration of a Potential Barrier by Electrons. Phys. Rev. 1930, 35, 1303–1309. [Google Scholar] [CrossRef]
- Miyazaki, T. Atom Tunneling Phenomena in Physics, Chemistry and Biology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Rao, S.; Klesse, G.; Stansfeld, P.J.; Tucker, S.J.; Sansom, M.S.P. A heuristic derived from analysis of the ion channel structural proteome permits the rapid identification of hydrophobic gates. Proc. Natl. Acad. Sci. USA 2019, 116, 13989–13995. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.; Lynch, C.I.; Klesse, G.; Oakley, G.E.; Stansfeld, P.J.; Tucker, S.J.; Sansom, M.S.P. Water and hydrophobic gates in ion channels and nanopores. Faraday Discuss. 2018, 209, 231–247. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.; Klesse, G.; Lynch, C.I.; Tucker, S.J.; Sansom, M.S.P. Molecular Simulations of Hydrophobic Gating of Pentameric Ligand Gated Ion Channels: Insights into Water and Ions. J. Phys. Chem. B 2021, 125, 981–994. [Google Scholar] [CrossRef]
- Tepper, H.L.; Voth, G.A. Mechanisms of Passive Ion Permeation through Lipid Bilayers: Insights from Simulations. J. Phys. Chem. B 2006, 110, 21327–21337. [Google Scholar] [CrossRef] [Green Version]
- Khavrutskii, I.V.; Gorfe, A.A.; Lu, B.; McCammon, J.A. Free Energy for the Permeation of Na+ and Cl− Ions and Their Ion-Pair through a Zwitterionic Dimyristoyl Phosphatidylcholine Lipid Bilayer by Umbrella Integration with Harmonic Fourier Beads. J. Am. Chem. Soc. 2009, 131, 1706–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Rawashdeh, B.M.; Qaswal, A.B.; Suleiman, A.; Zayed, F.M.; Al-Rawashdeh, S.M.; Tawalbeh, M.; Khreesha, L.; Alzubaidi, A.; Al-Zubidi, E.; Ghala, Z.; et al. The Quantum Tunneling of Ions Model Can Explain the Pathophysiology of Tinnitus. Brain Sci. 2022, 12, 426. [Google Scholar] [CrossRef] [PubMed]
- Payandeh, J.; El-Din, T.G.; Scheuer, T.; Zheng, N.; Catterall, W.A. Crystal structure of a voltage-gated sodium channel in two potentially inactivated states. Nature 2012, 486, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Cuello, L.G.; Jogini, V.; Cortes, D.M.; Perozo, E. Structural mechanism of C-type inactivation in K+ channels. Nature 2010, 466, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagnéris, C.; Naylor, C.E.; McCusker, E.C.; Wallace, B.A. Structural model of the open–closed–inactivated cycle of prokaryotic voltage-gated sodium channels. J. Gen. Physiol. 2014, 145, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serway, R.A.; Moses, C.J.; Moyer, C.A. Modern Physics; Cengage Learning: Boston, MA, USA, 2004. [Google Scholar]
- Chen, F.; Hihath, J.; Huang, Z.; Li, X.; Tao, N. Measurement of Single-Molecule Conductance. Annu. Rev. Phys. Chem. 2007, 58, 535–564. [Google Scholar] [CrossRef] [Green Version]
- Tsymbal, E.Y.; Mryasov, O.N.; LeClair, P.R. Spin-dependent tunnelling in magnetic tunnel junctions. J. Phys. Condens. Matter 2003, 15, R109–R142. [Google Scholar] [CrossRef]
- Bertil, H.; Bertil, H. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2001. [Google Scholar]
- Hall, J.E.; Hall, M.E. Guyton and Hall Textbook of Medical Physiology E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Qaswal, A.B. Quantum Electrochemical Equilibrium: Quantum Version of the Goldman–Hodgkin–Katz Equation. Quantum Rep. 2020, 2, 266–277. [Google Scholar] [CrossRef]
- Anastassiou, C.A.; Perin, R.; Markram, H.; Koch, C. Ephaptic coupling of cortical neurons. Nat. Neurosci. 2011, 14, 217–223. [Google Scholar] [CrossRef]
- Anastassiou, C.A.; Koch, C. Ephaptic coupling to endogenous electric field activity: Why bother? Curr. Opin. Neurobiol. 2015, 31, 95–103. [Google Scholar] [CrossRef]
- Bokil, H.; Laaris, N.; Blinder, K.; Ennis, M.; Keller, A. Ephaptic Interactions in the Mammalian Olfactory System. J. Neurosci. 2001, 21, RC173. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, H.; Sun, W.; Fu, Y.; Li, J.; Cheng, J.-X.; Nauman, E.; Shi, R. Compression Induces Acute Demyelination and Potassium Channel Exposure in Spinal Cord. J. Neurotrauma 2010, 27, 1109–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waxman, S.G. Membranes, Myelin, and the Pathophysiology of Multiple Sclerosis. New Engl. J. Med. 1982, 306, 1529–1533. [Google Scholar] [CrossRef] [PubMed]
- Jukkola, P.; Lovett-Racke, A.E.; Zamvil, S.S.; Gu, C. K+ channel alterations in the progression of experimental autoimmune encephalomyelitis. Neurobiol. Dis. 2012, 47, 280–293. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.; Sun, W. Potassium channel blockers as an effective treatment to restore impulse conduction in injured axons. Neurosci. Bull. 2011, 27, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Buonocore, M.; Bonezzi, C.; Barolat, G. Neurophysiological Evidence of Antidromic Activation of Large Myelinated Fibres in Lower Limbs During Spinal Cord Stimulation. Spine 2008, 33, E90–E93. [Google Scholar] [CrossRef]
- Hamada, M.S.; Kole, M.H.P. Myelin Loss and Axonal Ion Channel Adaptations Associated with Gray Matter Neuronal Hyperexcitability. J. Neurosci. 2015, 35, 7272–7286. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, M.; Yamada, Y.; Fukumoto, Y.; Sawano, S.; Minami, S.; Ikezoe, T.; Watanabe, Y.; Kimura, M.; Ichihashi, N. Increase in echo intensity and extracellular-to-intracellular water ratio is independently associated with muscle weakness in elderly women. Eur. J. Appl. Physiol. 2017, 117, 2001–2007. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.E. Voltage-Gated Channel Mechanosensitivity: Fact or Friction? Front. Physiol. 2011, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.A.; Lin, W.; Morris, T.; Banderali, U.; Juranka, P.F.; Morris, C.E. Membrane trauma and Na+ leak from Nav1. 6 channels. Am. J. Physiol.-Cell Physiol. 2009, 297, C823–C834. [Google Scholar] [CrossRef] [Green Version]
- Beyder, A.; Rae, J.L.; Bernard, C.; Strege, P.R.; Sachs, F.; Farrugia, G. Mechanosensitivity of Nav1.5, a voltage-sensitive sodium channel. J. Physiol. 2010, 588, 4969–4985. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yang, T.M.; Liedtke, W.; Simon, S.A. Chronic IL-1β Signaling Potentiates Voltage-Dependent Sodium Currents in Trigeminal Nociceptive Neurons. J. Neurophysiol. 2006, 95, 1478–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binshtok, A.M.; Wang, H.; Zimmermann, K.; Amaya, F.; Vardeh, D.; Shi, L.; Brenner, G.J.; Ji, R.R.; Bean, B.P.; Woolf, C.J.; et al. Nociceptors are interleukin-1β sensors. J. Neurosci. 2008, 28, 14062–14073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillespie, C.; Sherman, D.L.; Fleetwood-Walker, S.M.; Cottrell, D.F.; Tait, S.; Garry, E.M.; Wallace, V.C.; Ure, J.; Griffiths, I.R.; Smith, A.; et al. Peripheral Demyelination and Neuropathic Pain Behavior in Periaxin-Deficient Mice. Neuron 2000, 26, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Smith, M.T. Multiple sclerosis-induced neuropathic pain: Pharmacological management and pathophysiological insights from rodent EAE models. Inflammopharmacology 2014, 22, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waxman, S.G. Demyelination in spinal cord injury. J. Neurol. Sci. 1989, 91, 1–4. [Google Scholar] [CrossRef]
- Totoiu, M.O.; Keirstead, H.S. Spinal cord injury is accompanied by chronic progressive demyelination. J. Comp. Neurol. 2005, 486, 373–383. [Google Scholar] [CrossRef]
- Stadelmann, C.; Wegner, C.; Brück, W. Inflammation, demyelination, and degeneration—recent insights from MS pathology. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2011, 1812, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Cerda, F.P.; Gomez, M.V.S.; Matute, C. The link of inflammation and neurodegeneration in progressive multiple sclerosis. Mult. Scler. Demyelinating Disord. 2016, 1, 9. [Google Scholar] [CrossRef]
- Kelley, R.E. Ischemic demyelination. Neurol. Res. 2006, 28, 334–340. [Google Scholar] [CrossRef]
- Sharma, K.R.; Cross, J.; Farronay, O.; Ayyar, D.R.; Shebert, R.T.; Bradley, W.G. Demyelinating Neuropathy in Diabetes Mellitus. Arch. Neurol. 2002, 59, 758–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bril, V.; Blanchette, C.M.; Noone, J.M.; Runken, M.C.; Gelinas, D.; Russell, J.W. The dilemma of diabetes in chronic inflammatory demyelinating polyneuropathy. J. Diabetes Its Complicat. 2016, 30, 1401–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, J.; Jiménez-Martínez, R.; Troullinou, C.; Lucivero, V.G.; Tóth, G.; Mitchell, M.W. Measurement-induced, spatially-extended entanglement in a hot, strongly-interacting atomic system. Nat. Commun. 2020, 11, 2415. [Google Scholar] [CrossRef] [PubMed]
- Salari, V.; Tuszynski, J.; Rahnama, M.; Bernroider, G. Plausibility of quantum coherent states in biological systems. J. Phys. Conf. Ser. 2011, 306, 012075. [Google Scholar] [CrossRef]
- Hagan, S.; Hameroff, S.R.; Tuszyński, J.A. Quantum computation in brain microtubules: Decoherence and biological feasibility. Phys. Rev. E 2002, 65, 061901. [Google Scholar] [CrossRef] [Green Version]
- Gerlich, S.; Eibenberger, S.; Tomandl, M.; Nimmrichter, S.; Hornberger, K.; Fagan, P.J.; Tüxen, J.; Mayor, M.; Arndt, M. Quantum interference of large organic molecules. Nat. Commun. 2011, 2, 263. [Google Scholar] [CrossRef] [Green Version]
- Shayeghi, A.; Rieser, P.; Richter, G.; Sezer, U.; Rodewald, J.H.; Geyer, P.; Martinez, T.J.; Arndt, M. Matter-wave interference of a native polypeptide. Nat. Commun. 2020, 11, 1447. [Google Scholar] [CrossRef] [Green Version]

















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawafleh, S.; Qaswal, A.B.; Alali, O.; Zayed, F.M.; Al-Azzam, A.M.; Al-Kharouf, K.; Ali, M.B.; Albliwi, M.A.; Al-Hamarsheh, R.; Iswaid, M.; et al. Quantum Mechanical Aspects in the Pathophysiology of Neuropathic Pain. Brain Sci. 2022, 12, 658. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050658
Nawafleh S, Qaswal AB, Alali O, Zayed FM, Al-Azzam AM, Al-Kharouf K, Ali MB, Albliwi MA, Al-Hamarsheh R, Iswaid M, et al. Quantum Mechanical Aspects in the Pathophysiology of Neuropathic Pain. Brain Sciences. 2022; 12(5):658. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050658
Chicago/Turabian StyleNawafleh, Sager, Abdallah Barjas Qaswal, Obada Alali, Fuad Mohammed Zayed, Ahmed Mahmoud Al-Azzam, Khaled Al-Kharouf, Mo’ath Bani Ali, Moath Ahmad Albliwi, Rawan Al-Hamarsheh, Mohammad Iswaid, and et al. 2022. "Quantum Mechanical Aspects in the Pathophysiology of Neuropathic Pain" Brain Sciences 12, no. 5: 658. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050658