
Distribution of Adiponectin Receptors in the Brain of Adult Mouse: Effect of a Single Dose of the Adiponectin Receptor Agonist, AdipoRON, on Ischemic Stroke

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethic
2.2. Middle Cerebral Artery Occlusion (MCAO) Procedure and AdipoRON Treatment
2.3. Measurement of Infarct Volume
2.4. Neurological Evaluation
2.5. Tissue Preparation
2.6. RNA Extraction qPCR Analysis
2.7. Immunohistofluorescence
2.8. Western Blot
2.9. ELISA Assay
2.10. Microscopy
2.11. Statistical Analysis
3. Results
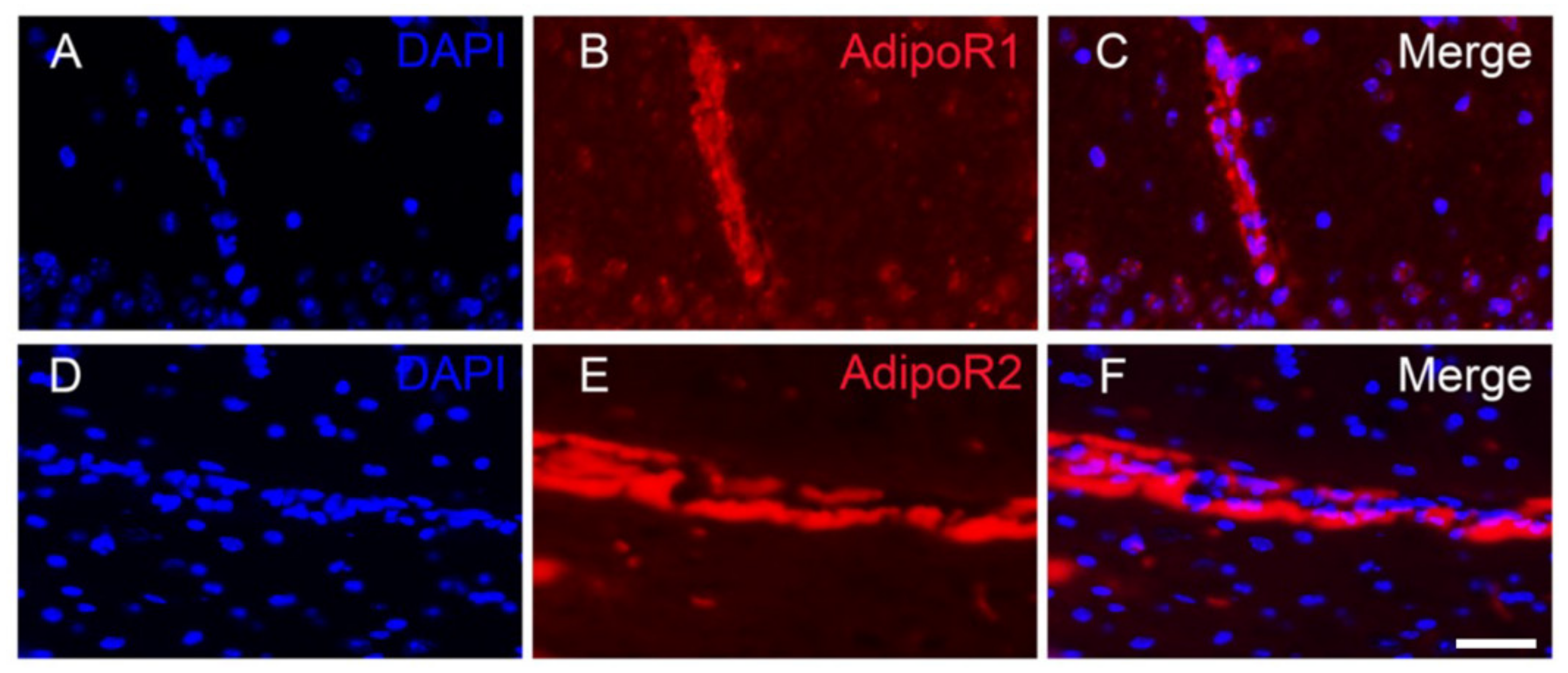
3.1. Adiponectin Receptors Are Widely Distributed in the Brain of Adult Mice
3.2. Adiponectin Receptors Are Mainly Expressed by Neurons
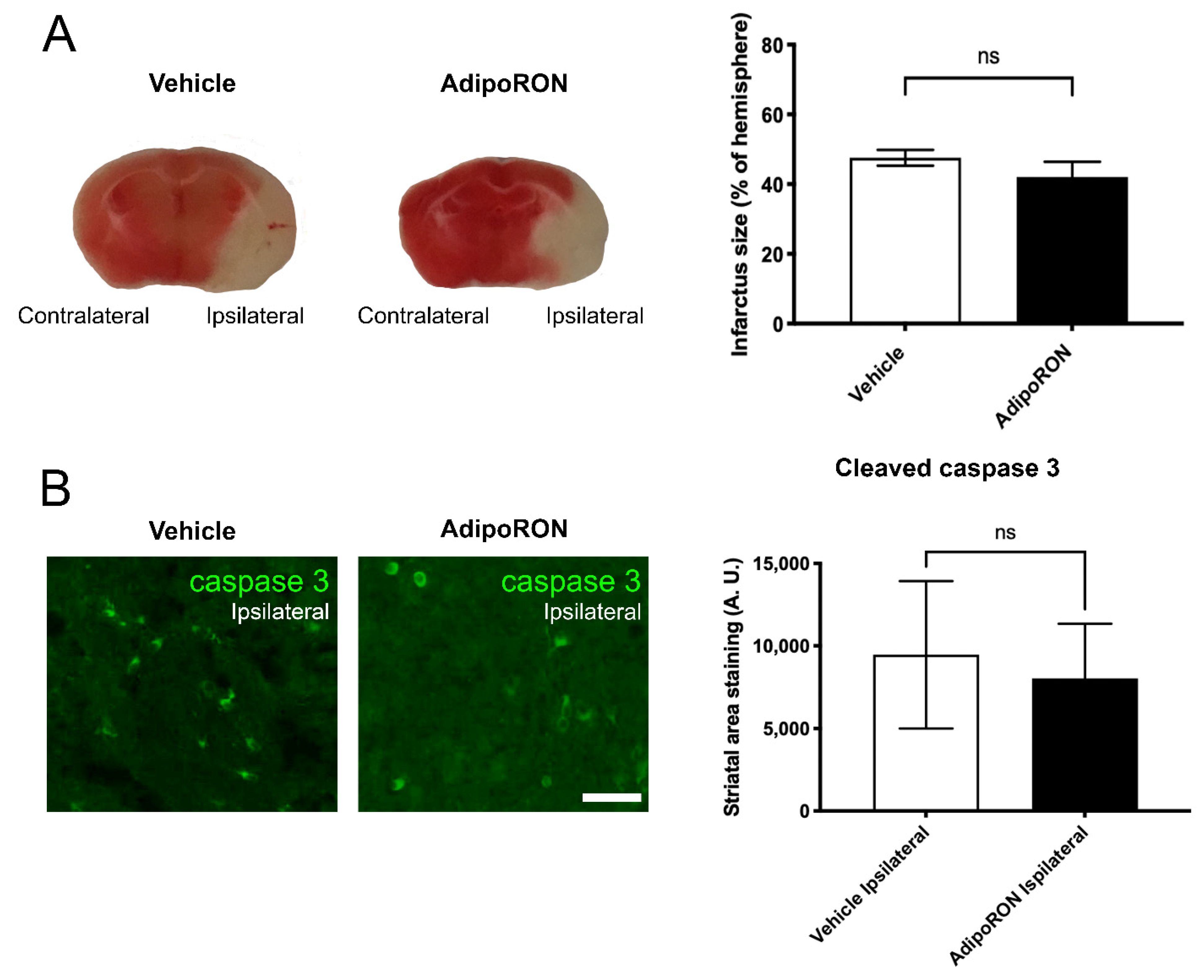
3.3. AdipoRON Treatment Has No Therapeutical Effect on the Infarct Size
4. Discussion
4.1. Adiponectin Receptors Are Widely Expressed throughout the Brain
4.2. Adiponectin Signaling as a Therapeutical Target for Stroke?
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Thundyil, J.; Pavlovski, D.; Sobey, C.G.; Arumugam, T.V. Adiponectin Receptor Signalling in the Brain: Adiponectin Receptor Signalling in the Brain. Br. J. Pharmacol. 2012, 165, 313–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, A.H.; Combs, T.P.; Du, X.; Brownlee, M.; Scherer, P.E. The Adipocyte-Secreted Protein Acrp30 Enhances Hepatic Insulin Action. Nat. Med. 2001, 7, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The Fat-Derived Hormone Adiponectin Reverses Insulin Resistance Associated with Both Lipoatrophy and Obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, M.; Asghari, S.; Zohoori, E.; Karamian, M. Hypoglycemic Effects of Three Iranian Edible Plants; Jujube, Barberry and Saffron: Correlation with Serum Adiponectin Level. Pak. J. Pharm. Sci. 2015, 28, 2095–2099. [Google Scholar] [PubMed]
- Hug, C.; Wang, J.; Ahmad, N.S.; Bogan, J.S.; Tsao, T.-S.; Lodish, H.F. T-Cadherin Is a Receptor for Hexameric and High-Molecular-Weight Forms of Acrp30/Adiponectin. Proc. Natl. Acad. Sci. USA 2004, 101, 10308–10313. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Kadowaki, T. Adiponectin Receptors: A Review of Their Structure, Function and How They Work. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 15–23. [Google Scholar] [CrossRef]
- Kumada, M.; Kihara, S.; Ouchi, N.; Kobayashi, H.; Okamoto, Y.; Ohashi, K.; Maeda, K.; Nagaretani, H.; Kishida, K.; Maeda, N.; et al. Adiponectin Specifically Increased Tissue Inhibitor of Metalloproteinase-1 Through Interleukin-10 Expression in Human Macrophages. Circulation 2004, 109, 2046–2049. [Google Scholar] [CrossRef] [Green Version]
- Wulster-Radcliffe, M.C.; Ajuwon, K.M.; Wang, J.; Christian, J.A.; Spurlock, M.E. Adiponectin Differentially Regulates Cytokines in Porcine Macrophages. Biochem. Biophys. Res. Commun. 2004, 316, 924–929. [Google Scholar] [CrossRef]
- Ajuwon, K.M.; Spurlock, M.E. Adiponectin Inhibits LPS-Induced NF-ΚB Activation and IL-6 Production and Increases PPARγ2 Expression in Adipocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1220–R1225. [Google Scholar] [CrossRef]
- Terjung, R. (Ed.) Comprehensive Physiology, 1st ed.; Wiley: Hoboken, NJ, USA, 2011; ISBN 978-0-470-65071-4. [Google Scholar]
- Liao, Y.; Takashima, S.; Maeda, N.; Ouchi, N.; Komamura, K.; Shimomura, I.; Hori, M.; Matsuzawa, Y.; Funahashi, T.; Kitakaze, M. Exacerbation of Heart Failure in Adiponectin-Deficient Mice Due to Impaired Regulation of AMPK and Glucose Metabolism. Cardiovasc. Res. 2005, 67, 705–713. [Google Scholar] [CrossRef]
- Ohashi, K.; Iwatani, H.; Kihara, S.; Nakagawa, Y.; Komura, N.; Fujita, K.; Maeda, N.; Nishida, M.; Katsube, F.; Shimomura, I.; et al. Exacerbation of Albuminuria and Renal Fibrosis in Subtotal Renal Ablation Model of Adiponectin-Knockout Mice. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1910–1917. [Google Scholar] [CrossRef]
- Sharma, K.; RamachandraRao, S.; Qiu, G.; Usui, H.K.; Zhu, Y.; Dunn, S.R.; Ouedraogo, R.; Hough, K.; McCue, P.; Chan, L.; et al. Adiponectin Regulates Albuminuria and Podocyte Function in Mice. J. Clin. Invest. 2008, 118, 1645–1656. [Google Scholar] [CrossRef]
- Almabouada, F.; Diaz-Ruiz, A.; Rabanal-Ruiz, Y.; Peinado, J.R.; Vazquez-Martinez, R.; Malagon, M.M. Adiponectin Receptors Form Homomers and Heteromers Exhibiting Distinct Ligand Binding and Intracellular Signaling Properties. J. Biol. Chem. 2013, 288, 3112–3125. [Google Scholar] [CrossRef] [Green Version]
- Guillod-Maximin, E.; Roy, A.F.; Vacher, C.M.; Aubourg, A.; Bailleux, V.; Lorsignol, A.; Penicaud, L.; Parquet, M.; Taouis, M. Adiponectin Receptors Are Expressed in Hypothalamus and Colocalized with Proopiomelanocortin and Neuropeptide Y in Rodent Arcuate Neurons. J. Endocrinol. 2009, 200, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Bloemer, J.; Pinky, P.D.; Govindarajulu, M.; Hong, H.; Judd, R.; Amin, R.H.; Moore, T.; Dhanasekaran, M.; Reed, M.N.; Suppiramaniam, V. Role of Adiponectin in Central Nervous System Disorders. Neural Plast. 2018, 2018, 4593530. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-Sequencing Transcriptome and Splicing Database of Glia, Neurons, and Vascular Cells of the Cerebral Cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef]
- Rastegar, S.; Parimisetty, A.; Cassam Sulliman, N.; Narra, S.S.; Weber, S.; Rastegar, M.; Viranaicken, W.; Couret, D.; Planesse, C.; Strähle, U.; et al. Expression of Adiponectin Receptors in the Brain of Adult Zebrafish and Mouse: Links with Neurogenic Niches and Brain Repair. J. Comp. Neurol. 2019, 527, 2317–2333. [Google Scholar] [CrossRef]
- Bloemer, J.; Pinky, P.D.; Smith, W.D.; Bhattacharya, D.; Chauhan, A.; Govindarajulu, M.; Hong, H.; Dhanasekaran, M.; Judd, R.; Amin, R.H.; et al. Adiponectin Knockout Mice Display Cognitive and Synaptic Deficits. Front. Endocrinol. 2019, 10, 819. [Google Scholar] [CrossRef] [Green Version]
- Formolo, D.A.; Lee, T.H.-Y.; Yau, S.-Y. Increasing Adiponergic System Activity as a Potential Treatment for Depressive Disorders. Mol. Neurobiol. 2019, 56, 7966–7976. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Choi, S.-M.; Whitcomb, D.J.; Kim, B.C. Adiponectin Controls the Apoptosis and the Expression of Tight Junction Proteins in Brain Endothelial Cells through AdipoR1 under Beta Amyloid Toxicity. Cell Death Dis. 2017, 8, e3102. [Google Scholar] [CrossRef] [Green Version]
- Jian, M.; Kwan, J.S.-C.; Bunting, M.; Ng, R.C.-L.; Chan, K.H. Adiponectin Suppresses Amyloid-β Oligomer (AβO)-Induced Inflammatory Response of Microglia via AdipoR1-AMPK-NF-ΚB Signaling Pathway. J. Neuroinflamm. 2019, 16, 110. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Zhao, L.; Liu, H.; Guo, H.; Guo, W.; Zheng, L.; Liu, X.; Wu, X.; Luo, J.; Li, X.; et al. Adiponectin Confers Neuroprotection against Cerebral Ischemia-Reperfusion Injury through Activating the CAMP/PKA-CREB-BDNF Signaling. Brain Res. Bull. 2018, 143, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Luo, J.; Liu, H.; Cui, W.; Guo, K.; Zhao, L.; Bai, H.; Guo, W.; Guo, H.; Feng, D.; et al. Recombinant Adiponectin Peptide Ameliorates Brain Injury Following Intracerebral Hemorrhage by Suppressing Astrocyte-Derived Inflammation via the Inhibition of Drp1-Mediated Mitochondrial Fission. Transl. Stroke Res. 2020, 11, 924–939. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, X.; Lu, X.-Y. Adiponectin Exerts Neurotrophic Effects on Dendritic Arborization, Spinogenesis, and Neurogenesis of the Dentate Gyrus of Male Mice. Endocrinology 2016, 157, 2853–2869. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Liang, Y.; Chen, K.; Yau, S.-Y.; Sun, X.; Cheng, K.K.-Y.; Xu, A.; So, K.-F.; Li, A. Potential Involvement of Adiponectin Signaling in Regulating Physical Exercise-Elicited Hippocampal Neurogenesis and Dendritic Morphology in Stressed Mice. Front. Cell. Neurosci. 2020, 14, 189. [Google Scholar] [CrossRef]
- Dávalos, A.; Alvarez-Sabín, J.; Castillo, J.; Díez-Tejedor, E.; Ferro, J.; Martínez-Vila, E.; Serena, J.; Segura, T.; Cruz, V.T.; Masjuan, J.; et al. Citicoline in the Treatment of Acute Ischaemic Stroke: An International, Randomised, Multicentre, Placebo-Controlled Study (ICTUS Trial). Lancet 2012, 380, 349–357. [Google Scholar] [CrossRef]
- Holland, W.L.; Scherer, P.E.; Scherer, P.E. Ronning After the Adiponectin Receptors. Science 2014, 342, 1460–1461. [Google Scholar] [CrossRef] [Green Version]
- Hong, K.; Lee, S.; Li, R.; Yang, Y.; Tanner, M.A.; Wu, J.; Hill, M.A. Adiponectin Receptor Agonist, AdipoRon, Causes Vasorelaxation Predominantly Via a Direct Smooth Muscle Action. Microcirculation 2016, 23, 207–220. [Google Scholar] [CrossRef]
- Okada-Iwabu, M.; Yamauchi, T.; Iwabu, M.; Honma, T.; Hamagami, K.; Matsuda, K.; Yamaguchi, M.; Tanabe, H.; Kimura-Someya, T.; Shirouzu, M.; et al. A Small-Molecule AdipoR Agonist for Type 2 Diabetes and Short Life in Obesity. Nature 2013, 503, 493–499. [Google Scholar] [CrossRef]
- Otvos, L., Jr. Potential Adiponectin Receptor Response Modifier Therapeutics. Front. Endocrinol. 2019, 10, 539. [Google Scholar] [CrossRef] [Green Version]
- Dib, J.; Thomas, A.; Delahaut, P.; Fichant, E.; Schänzer, W.; Thevis, M. Identification and Characterization of in Vitro and in Vivo Generated Metabolites of the Adiponectin Receptor Agonists AdipoRon and 112254. J. Pharm. Biomed. Anal. 2016, 125, 68–76. [Google Scholar] [CrossRef]
- Akimoto, M.; Maruyama, R.; Kawabata, Y.; Tajima, Y.; Takenaga, K. Antidiabetic Adiponectin Receptor Agonist AdipoRon Suppresses Tumour Growth of Pancreatic Cancer by Inducing RIPK1/ERK-Dependent Necroptosis. Cell Death Dis. 2018, 9, 804. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, T.; Lakota, K.; Taniguchi, T.; Yoshizaki, A.; Sato, S.; Hong, W.; Zhou, X.; Sodin-Semrl, S.; Fang, F.; Asano, Y.; et al. An Orally-Active Adiponectin Receptor Agonist Mitigates Cutaneous Fibrosis, Inflammation and Microvascular Pathology in a Murine Model of Systemic Sclerosis. Sci. Rep. 2018, 8, 11843. [Google Scholar] [CrossRef] [Green Version]
- Lindfors, S.; Polianskyte-Prause, Z.; Bouslama, R.; Lehtonen, E.; Mannerla, M.; Nisen, H.; Tienari, J.; Salmenkari, H.; Forsgård, R.; Mirtti, T.; et al. Adiponectin Receptor Agonist AdipoRon Ameliorates Renal Inflammation in Diet-Induced Obese Mice and Endotoxin-Treated Human Glomeruli Ex Vivo. Diabetologia 2021, 64, 1866–1879. [Google Scholar] [CrossRef]
- Ng, R.C.-L.; Jian, M.; Ma, O.K.-F.; Bunting, M.; Kwan, J.S.-C.; Zhou, G.-J.; Senthilkumar, K.; Iyaswamy, A.; Chan, P.-K.; Li, M.; et al. Chronic Oral Administration of AdipoRon Reverses Cognitive Impairments and Ameliorates Neuropathology in an Alzheimer’s Disease Mouse Model. Mol. Psychiatry 2020, 26, 5669–5689. [Google Scholar] [CrossRef]
- Zheng, J.; Sun, Z.; Liang, F.; Xu, W.; Lu, J.; Shi, L.; Shao, A.; Yu, J.; Zhang, J. AdipoRon Attenuates Neuroinflammation After Intracerebral Hemorrhage Through AdipoR1-AMPK Pathway. Neuroscience 2019, 412, 116–130. [Google Scholar] [CrossRef]
- Chen, B.; Liao, W.-Q.; Xu, N.; Xu, H.; Wen, J.-Y.; Yu, C.-A.; Liu, X.-Y.; Li, C.-L.; Zhao, S.-M.; Campbell, W. Adiponectin Protects against Cerebral Ischemia–Reperfusion Injury through Anti-Inflammatory Action. Brain Res. 2009, 1273, 129–137. [Google Scholar] [CrossRef]
- Song, W.; Huo, T.; Guo, F.; Wang, H.; Wei, H.; Yang, Q.; Dong, H.; Wang, Q.; Xiong, L. Globular Adiponectin Elicits Neuroprotection by Inhibiting NADPH Oxidase-Mediated Oxidative Damage in Ischemic Stroke. Neuroscience 2013, 248, 136–144. [Google Scholar] [CrossRef]
- Song, W.; Guo, F.; Zhong, H.; Liu, L.; Yang, R.; Wang, Q.; Xiong, L. Therapeutic Window of Globular Adiponectin against Cerebral Ischemia in Diabetic Mice: The Role of Dynamic Alteration of Adiponectin/Adiponectin Receptor Expression. Sci. Rep. 2015, 5, 17310. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Ito, Y.; Tsuchida, A.; Yokomizo, T.; Kita, S.; Sugiyama, T.; Miyagishi, M.; Hara, K.; Tsunoda, M.; et al. Cloning of Adiponectin Receptors That Mediate Antidiabetic Metabolic Effects. Nature 2003, 423, 762–769. [Google Scholar] [CrossRef]
- Zhang, C.; Zhen, L.; Fang, Z.; Yu, L.; Zhang, Y.; Wei, H.; Jia, J.; Wang, S. Adiponectin Treatment Attenuates Cerebral Ischemia-Reperfusion Injury through HIF-1α-Mediated Antioxidation in Mice. Oxid. Med. Cell. Longev. 2021, 2021, 5531048. [Google Scholar] [CrossRef] [PubMed]
- Zatorski, H.; Salaga, M.; Zielińska, M.; Majchrzak, K.; Binienda, A.; Kordek, R.; Małecka-Panas, E.; Fichna, J. AdipoRon, an Orally Active, Synthetic Agonist of AdipoR1 and AdipoR2 Receptors Has Gastroprotective Effect in Experimentally Induced Gastric Ulcers in Mice. Molecules 2021, 26, 2946. [Google Scholar] [CrossRef] [PubMed]
- Isayama, K.; Pitts, L.H.; Nishimura, M.C. Evaluation of 2,3,5-Triphenyltetrazolium Chloride Staining to Delineate Rat Brain Infarcts. Stroke 1991, 22, 1394–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couret, D.; Bourane, S.; Catan, A.; Nativel, B.; Planesse, C.; Dorsemans, A.-C.; Ait-Arsa, I.; Cournot, M.; Rondeau, P.; Patche, J.; et al. A Hemorrhagic Transformation Model of Mechanical Stroke Therapy with Acute Hyperglycemia in Mice. J. Comp. Neurol. 2018, 526, 1006–1016. [Google Scholar] [CrossRef]
- Menzies, S.A.; Hoff, J.T.; Betz, A.L. Middle Cerebral Artery Occlusion in Rats: A Neurological and Pathological Evaluation of a Reproducible Model. Neurosurgery 1992, 31, 100–106; discussion 106–107. [Google Scholar] [CrossRef]
- Nativel, B.; Marimoutou, M.; Thon-Hon, V.G.; Gunasekaran, M.K.; Andries, J.; Stanislas, G.; Planesse, C.; Da Silva, C.R.; Césari, M.; Iwema, T.; et al. Soluble HMGB1 Is a Novel Adipokine Stimulating IL-6 Secretion through RAGE Receptor in SW872 Preadipocyte Cell Line: Contribution to Chronic Inflammation in Fat Tissue. PLoS ONE 2013, 8, e76039. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Kang, S.M.; Kim, E.; Kim, C.-H.; Song, H.-T.; Lee, J.E. Adiponectin Receptor-Mediated Signaling Ameliorates Cerebral Cell Damage and Regulates the Neurogenesis of Neural Stem Cells at High Glucose Concentrations: An in Vivo and in Vitro Study. Cell Death Dis. 2015, 6, e1844. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, J.H.; Sherchan, P.; Krafft, P.R.; Zhao, W.; Wang, S.; Chen, S.; Guo, Z.; Tang, J. Administration of RCTRP9 Attenuates Neuronal Apoptosis Through AdipoR1/PI3K/Akt Signaling Pathway after ICH in Mice. Cell Transplant. 2019, 28, 756–766. [Google Scholar] [CrossRef]
- Kawanami, D.; Maemura, K.; Takeda, N.; Harada, T.; Nojiri, T.; Imai, Y.; Manabe, I.; Utsunomiya, K.; Nagai, R. Direct Reciprocal Effects of Resistin and Adiponectin on Vascular Endothelial Cells: A New Insight into Adipocytokine–Endothelial Cell Interactions. Biochem. Biophys. Res. Commun. 2004, 314, 415–419. [Google Scholar] [CrossRef]
- Adya, R.; Tan, B.K.; Chen, J.; Randeva, H.S. Protective Actions of Globular and Full-Length Adiponectin on Human Endothelial Cells: Novel Insights into Adiponectin-Induced Angiogenesis. J. Vasc. Res. 2012, 49, 534–543. [Google Scholar] [CrossRef]
- Sun, L.; Li, H.; Tai, L.W.; Gu, P.; Cheung, C.W. Adiponectin Regulates Thermal Nociception in a Mouse Model of Neuropathic Pain. Br. J. Anaesth. 2018, 120, 1356–1367. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Guo, H.; Zhao, L.; Wang, B.; Liu, H.; Yue, L.; Bai, H.; Jiang, H.; Gao, L.; Feng, D.; et al. Adiponectin Attenuates NADPH Oxidase-Mediated Oxidative Stress and Neuronal Damage Induced by Cerebral Ischemia-Reperfusion Injury. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2017, 1863, 3265–3276. [Google Scholar] [CrossRef]
- Gairolla, J.; Kler, R.; Modi, M.; Khurana, D. Leptin and Adiponectin: Pathophysiological Role and Possible Therapeutic Target of Inflammation in Ischemic Stroke. Rev. Neurosci. 2017, 28, 295–306. [Google Scholar] [CrossRef]
- Yu, J.; Zheng, J.; Lu, J.; Sun, Z.; Wang, Z.; Zhang, J. AdipoRon Protects Against Secondary Brain Injury After Intracerebral Hemorrhage via Alleviating Mitochondrial Dysfunction: Possible Involvement of AdipoR1–AMPK–PGC1α Pathway. Neurochem. Res. 2019, 44, 1678–1689. [Google Scholar] [CrossRef]
- Liu, B.; Liu, J.; Wang, J.; Liu, C.; Yan, H. AdipoRon Improves Cognitive Dysfunction of Alzheimer’s Disease and Rescues Impaired Neural Stem Cell Proliferation through AdipoR1/AMPK Pathway. Exp. Neurol. 2020, 327, 113249. [Google Scholar] [CrossRef]
- Kim, Y.; Lim, J.H.; Kim, M.Y.; Kim, E.N.; Yoon, H.E.; Shin, S.J.; Choi, B.S.; Kim, Y.-S.; Chang, Y.S.; Park, C.W. The Adiponectin Receptor Agonist AdipoRon Ameliorates Diabetic Nephropathy in a Model of Type 2 Diabetes. J. Am. Soc. Nephrol. 2018, 29, 1108–1127. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.H.; Christie, B.R.; van Praag, H.; Lin, K.; Siu, P.M.-F.; Xu, A.; So, K.-F.; Yau, S.-Y. AdipoRon Treatment Induces a Dose-Dependent Response in Adult Hippocampal Neurogenesis. Int. J. Mol. Sci. 2021, 22, 2068. [Google Scholar] [CrossRef]
- Rudilosso, S.; Rodríguez-Vázquez, A.; Urra, X.; Arboix, A. The Potential Impact of Neuroimaging and Translational Research on the Clinical Management of Lacunar Stroke. Int. J. Mol. Sci. 2022, 23, 1497. [Google Scholar] [CrossRef]
- Arboix, A.; Cabeza, N.; García-Eroles, L.; Massons, J.; Oliveres, M.; Targa, C.; Balcells, M. Relevance of Transient Ischemic Attack to Early Neurological Recovery after Nonlacunar Ischemic Stroke. Cerebrovasc. Dis. 2004, 18, 304–311. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse | Ensembl ID |
|---|---|---|---|
| Il-6 | CAACAGACTTCCATCCAGTTGC | TTGCCGAGTAGATCTCAAAGTGAC | ENSMUSG00000025746 |
| Nfκβ | GTGATGGGCCTTCACACACA | CATTTGAACACTGCTTTGACT | ENSMUSG00000030595 |
| Nrf2 | TCCCATTTGTAGATGACCATGAG | CCATGTCCTGCTCTATGCTG | ENSMUSG00000015839 |
| Tnfa | GTTCTGTCTACTGAACTTCGGG | CAGGCTTGTCACTCGAATTTTG | ENSMUSG00000024401 |
| Gapdh | CTTTGTCAAGCTCATTTCCTGG | TCTTGCTCAGTGTCCTTGC | ENSMUSG00000020932 |
| Antibodies (Marker) | Host | Type | Reference | RRID |
|---|---|---|---|---|
| AdipoR1 | Rabbit | Monoclonal | ab240022 | AB_2221906 |
| AdipoR2 | Rabbit | Polyclonal | LS-C34900 | AB_2222064 |
| GFAP (astrocyte) | Goat | Polyclonal | ab53554 | AB_880202 |
| Iba1 (microglia) | Goat | Polyclonal | ab5076 | AB_2224402 |
| HuC/D (neuron) | Mouse | Monoclonal | A-21271 Clone 16A11 | AB_221448 |
| Cleaved-Caspase 3 (apoptose) | Rabbit | Polyclonal | ab13847 | AB_443014 |
| PCNA (proliferation) | Mouse | Monoclonal | MO879 Clone PC10 | AB_2160651 |
| NRF2 | Host | Polyclonal | ab31163 | AB_881705 |
| Antibodies | Reference | RRID |
|---|---|---|
| Donkey anti-goat Alexa Fluor 488 | A11055 (ThermoFisher) | AB_2534102 |
| Donkey anti-mouse Alexa Fluor 488 | ab150105 (Abcam) | AB_2732856 |
| Donkey anti-rabbit Alexa Fluor 594 | ab150064 (Abcam) | AB_2734146 |
| Donkey anti-rabbit Alexa Fluor 488 | ab150061 (Abcam) | AB_2571722 |
| Donkey anti-rabbit-HRP | 711-035-152 (Jackson IR) | AB_10015282 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clain, J.; Couret, D.; Planesse, C.; Krejbich-Trotot, P.; Meilhac, O.; Lefebvre d’Hellencourt, C.; Viranaicken, W.; Diotel, N. Distribution of Adiponectin Receptors in the Brain of Adult Mouse: Effect of a Single Dose of the Adiponectin Receptor Agonist, AdipoRON, on Ischemic Stroke. Brain Sci. 2022, 12, 680. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050680
Clain J, Couret D, Planesse C, Krejbich-Trotot P, Meilhac O, Lefebvre d’Hellencourt C, Viranaicken W, Diotel N. Distribution of Adiponectin Receptors in the Brain of Adult Mouse: Effect of a Single Dose of the Adiponectin Receptor Agonist, AdipoRON, on Ischemic Stroke. Brain Sciences. 2022; 12(5):680. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050680
Chicago/Turabian StyleClain, Julien, David Couret, Cynthia Planesse, Pascale Krejbich-Trotot, Olivier Meilhac, Christian Lefebvre d’Hellencourt, Wildriss Viranaicken, and Nicolas Diotel. 2022. "Distribution of Adiponectin Receptors in the Brain of Adult Mouse: Effect of a Single Dose of the Adiponectin Receptor Agonist, AdipoRON, on Ischemic Stroke" Brain Sciences 12, no. 5: 680. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050680