
Encode a Letter and Get Its Location for Free? Assessing Incidental Binding of Verbal and Spatial Features


Abstract
:1. Introduction
2. Experiment 1
2.1. Method
2.1.1. Participants
2.1.2. Apparatus and Materials
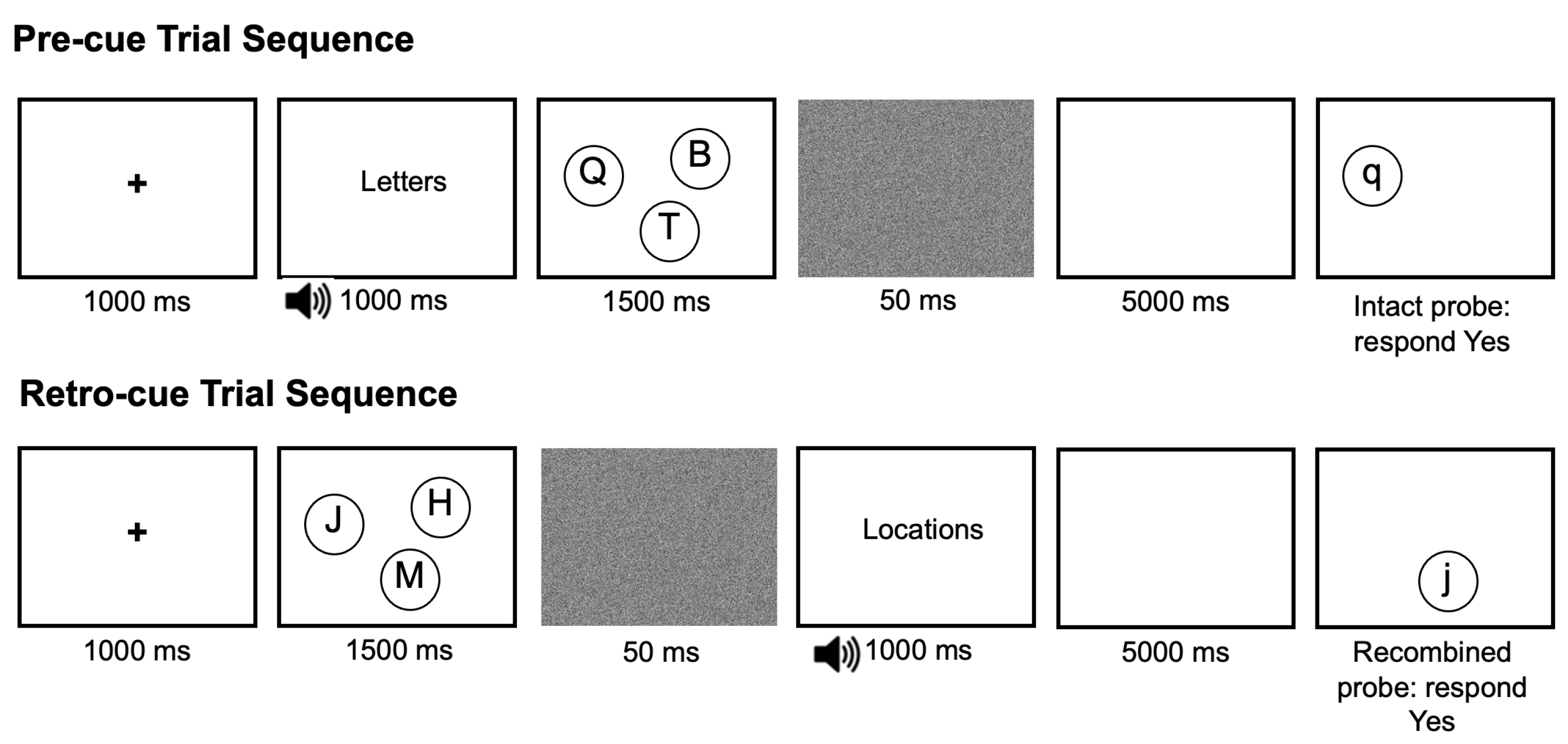
2.1.3. Procedure
2.2. Results
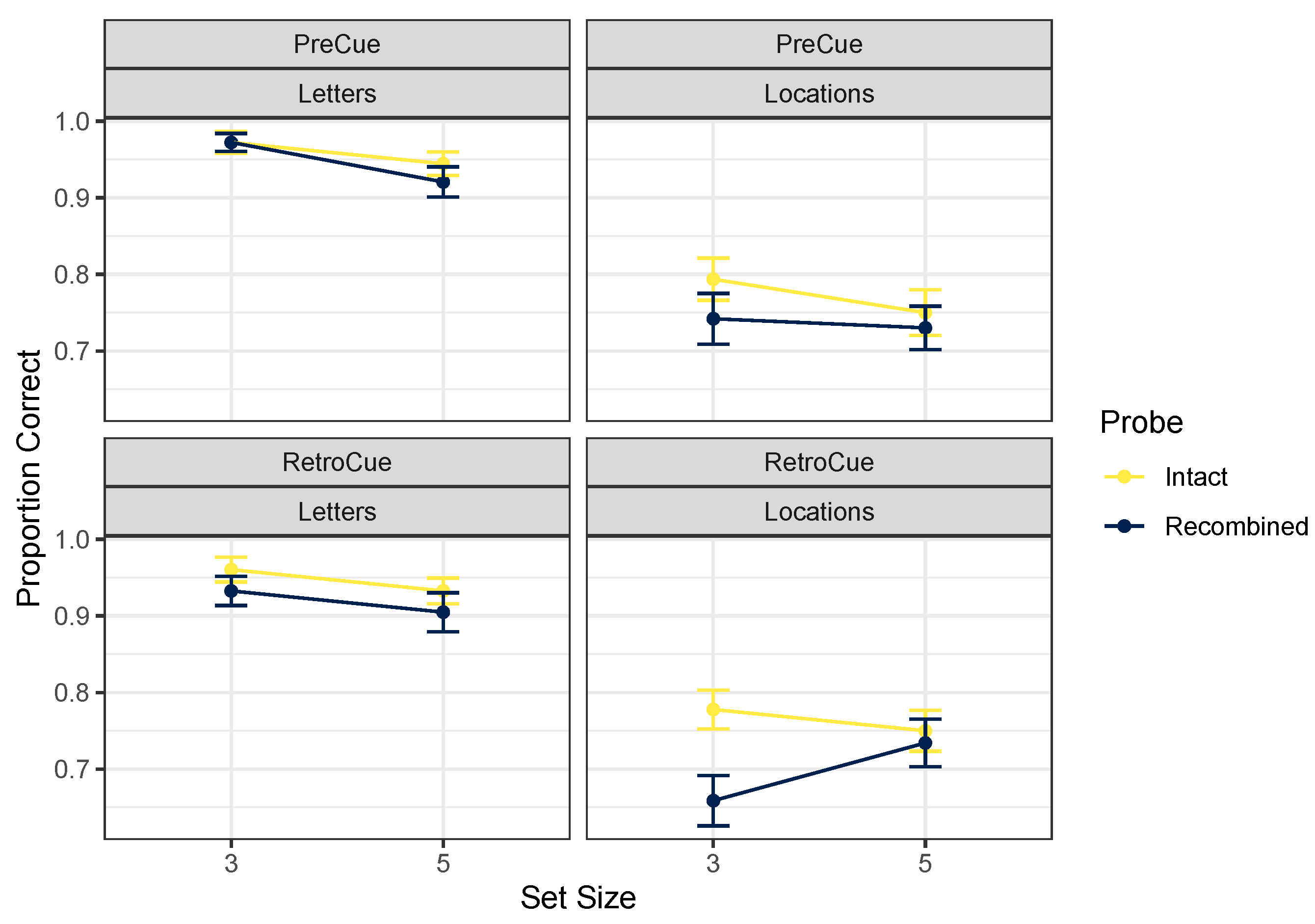
2.2.1. Accuracy
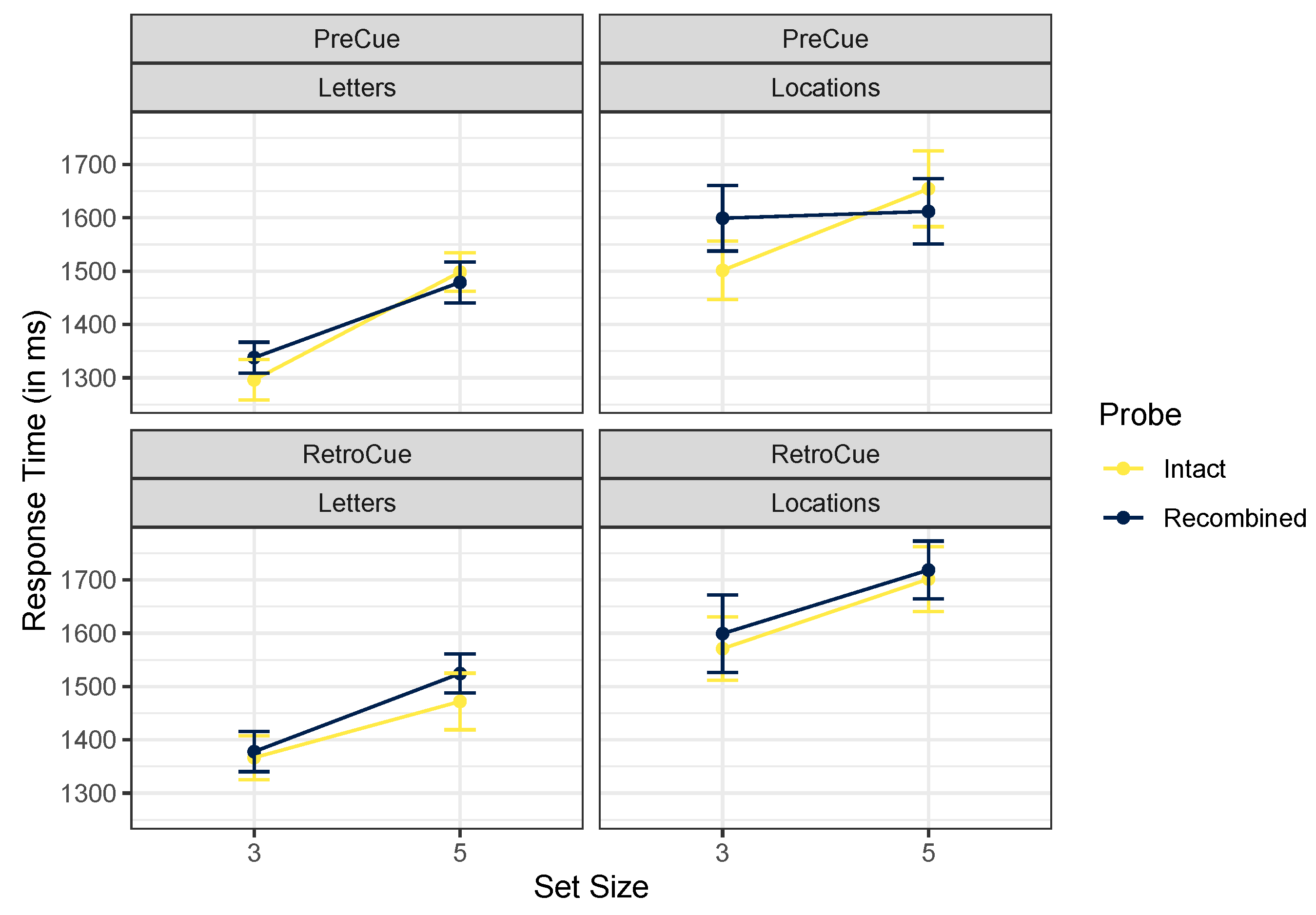
2.2.2. Response Times
2.3. Discussion
3. Experiment 2
3.1. Method
3.1.1. Participants
3.1.2. Materials
3.1.3. Design
3.1.4. Procedure
3.2. Results
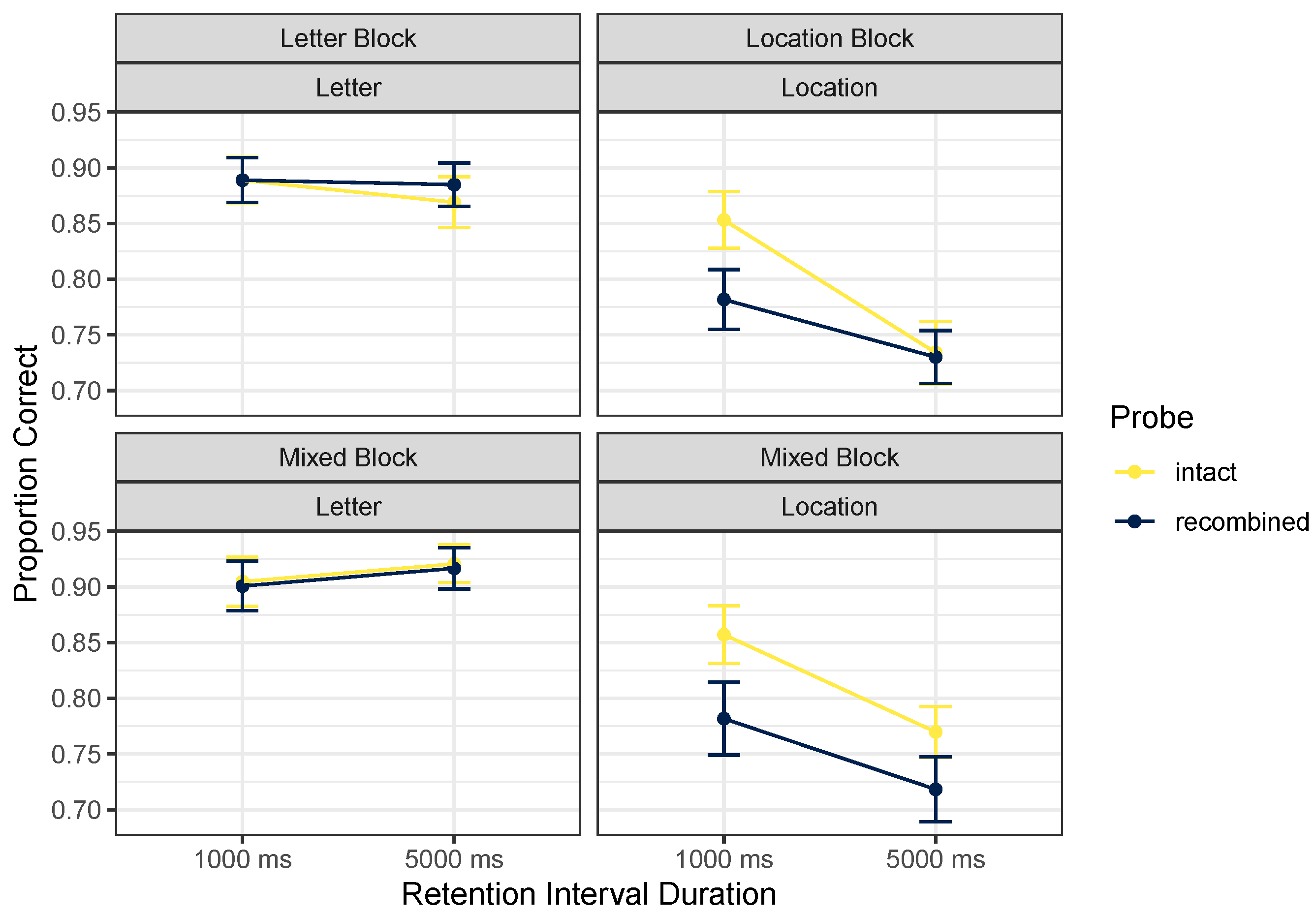
3.2.1. Accuracy
3.2.2. Response Times
3.3. Discussion
4. General Discussion
Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morey, C.C. Maintaining binding in working memory: Comparing the effects of intentional goals and incidental affordances. Conscious. Cogn. Int. J. 2011, 20, 920–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsley, J.V.; Parmentier, F.B.R. The asymmetry and temporal dynamics of incidental letter–location bindings in working memory. Q. J. Exp. Psychol. 2015, 68, 433–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabhakaran, V.; Narayanan, K.; Zhao, Z.; Gabrieli, J.D.E. Integration of diverse information in working memory within the frontal lobe. Nat. Neurosci. 2000, 3, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Darling, S.; Allen, R.J.; Havelka, J. Visuospatial bootstrapping: When visuospatial and verbal memory work together. Curr. Dir. Psychol. Sci. 2017, 26, 3–9. [Google Scholar] [CrossRef]
- Langerock, N.; Vergauwe, E.; Barrouillet, P. The maintenance of cross-domain associations in the episodic buffer. J. Exp. Psychol.-Learn. Mem. Cogn. 2014, 40, 1096–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baddeley, A.D. Working memory: Theories, models, and controversies. Annu. Rev. Psychol. 2012, 63, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.M.; Macken, B. In the beginning was the deed: Verbal short-term memory as object-oriented action. Curr. Dir. Psychol. Sci. 2018, 27, 351–356. [Google Scholar] [CrossRef]
- Cowan, N. The magical number 4 in short-term memory: A reconsideration of mental storage capacity. Behav. Brain Sci. 2001, 24, 87–185. [Google Scholar] [CrossRef] [Green Version]
- Luck, S.J.; Vogel, E.K. The capacity of visual working memory for features and conjunctions. Nature 1997, 390, 279–281. [Google Scholar] [CrossRef]
- Hardman, K.O.; Cowan, N. Remembering complex objects in visual working memory: Do capacity limits restrict objects or features? J. Exp. Psychol.-Learn. Mem. Cogn. 2015, 41, 325–347. [Google Scholar] [CrossRef] [Green Version]
- Oberauer, K.; Eichenberger, S. Visual working memory declines when more features must be remembered for each object. Mem. Cogn. 2013, 41, 1212–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logie, R.H.; Brockmole, J.R.; Jaswal, S. Feature binding in visual short-term memory is unaffected by task-irrelevant changes of location, shape, and color. Mem. Cogn. 2010, 39, 24–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingworth, A.; Rasmussen, I.P. Binding objects to locations: The relationship between object files and visual working memory. J. Exp. Psychol. Hum. Percept. Perform. 2010, 36, 543–564. [Google Scholar] [CrossRef] [Green Version]
- Morey, C.C. Integrated cross-domain object storage in working memory: Evidence from a verbal-spatial memory task. Q. J. Exp. Psychol. 2009, 62, 2235–2251. [Google Scholar] [CrossRef] [Green Version]
- Guérard, K.; Tremblay, S.; Saint-Aubin, J. Similarity and binding in memory: Bound to be detrimental. Q. J. Exp. Psychol. 2009, 62, 26–32. [Google Scholar] [CrossRef]
- Parmentier, F.B.R.; Elford, G.; Maybery, M. Transitional information in spatial serial memory: Path characteristics affect recall performance. J. Exp. Psychol. Learn. Mem. Cogn. 2005, 31, 412–427. [Google Scholar] [CrossRef]
- Parmentier, F.B.R.; Andrés, P. The impact of path crossing on visuo-spatial serial memory: Encoding or rehearsal effect? Q. J. Exp. Psychol. 2006, 59, 1867–1874. [Google Scholar] [CrossRef]
- Allan, A.; Morey, C.C.; Darling, S.; Allen, R.J.; Havelka, J. On the right track? Investigating the effect of path characteristics on visuospatial bootstrapping in verbal serial recall. J. Cogn. 2017, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Guerard, K.; Morey, C.C.; Lagace, S.; Tremblay, S. Asymmetric binding in serial memory for verbal and spatial information. Mem. Cogn. 2013, 41, 378–391. [Google Scholar] [CrossRef] [Green Version]
- Schneider, W.; Eschmann, A.; Zuccolotto, A. E-Prime User’s Guide; Psychology Software Tools, Inc.: Pittsburgh, PA, USA, 2002. [Google Scholar]
- Campo, P.; Maestú, F.; Capilla, A.; Morales, M.; Fernández, S.; del Río, D.; Ortiz, T. Temporal dynamics of parietal activity during word-location binding. Neuropsychology 2008, 22, 85–99. [Google Scholar] [CrossRef]
- Elsley, J.V.; Parmentier, F.B.R. Is verbal–spatial binding in working memory impaired by a concurrent memory load? Q. J. Exp. Psychol. 2009, 62, 1696–1705. [Google Scholar] [CrossRef] [PubMed]
- Morey, R.D.; Rouder, J.N.; Jamil, T.; Urbanek, S.; Forner, K.; Ly, A. BayesFactor: Computation of Bayes Factors for Common Designs. 2018. Available online: https://CRAN.R-project.org/package=BayesFactor (accessed on 20 February 2022).
- Rouder, J.N.; Morey, R.D.; Speckman, P.L.; Province, J.M. Default Bayes factors for ANOVA designs. J. Math. Psychol. 2012, 56, 356–374. [Google Scholar] [CrossRef]
- Morey, R.D. Confidence intervals from normalized data: A correction to Cousineau (2005). Tutor. Quant. Methods Psychol. 2008, 4, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Grange, J. Trimr: An Implementation of Common Response Time Trimming Methods. 2015. Available online: https://cran.r-project.org/web/packages/trimr/index.html (accessed on 20 February 2022).
- Henninger, F.; Shevchenko, Y.; Mertens, U.K.; Kieslich, P.J.; Hilbig, B.E. Lab.js: A free, open, online study builder. Behav. Res. Methods 2021, 54, 556–573. [Google Scholar] [CrossRef] [PubMed]
- Maybery, M.T.; Clissa, P.J.; Parmentier, F.B.R.; Leung, D.; Harsa, G.; Fox, A.M.; Jones, D.M. Binding of verbal and spatial features in auditory working memory. J. Mem. Lang. 2009, 61, 112–133. [Google Scholar] [CrossRef]
- Campo, P.; Poch, C.; Parmentier, F.B.; Moratti, S.; Elsley, J.V.; Castellanos, N.P.; Ruiz-Vargas, J.M.; del Pozo, F.; Maestú, F. Oscillatory activity in prefrontal and posterior regions during implicit letter-location binding. Neuroimage 2010, 49, 2807–2815. [Google Scholar] [CrossRef] [Green Version]
- Treisman, A.; Zhang, W. Location and binding in visual working memory. Mem. Cogn. 2006, 34, 1704–1719. [Google Scholar] [CrossRef] [Green Version]
- Udale, R.; Farrell, S.; Kent, C. No evidence of binding items to spatial configuration representations in visual working memory. Mem. Cogn. 2018, 46, 955–968. [Google Scholar] [CrossRef]
- Rouder, J.N.; Haaf, J.M. Are there reliable qualitative individual differences in cognition? J. Cogn. 2021, 4, 46. [Google Scholar] [CrossRef]
- Chen, H.; Wyble, B. Amnesia for object attributes: Failure to report attended information that had just reached conscious awareness. Psychol. Sci. 2015, 26, 203–210. [Google Scholar] [CrossRef] [Green Version]
- van Dijck, J.-P.; Fias, W. A working memory account for spatial–numerical associations. Cognition 2011, 119, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Schneegans, S.; Harrison, W.J.; Bays, P.M. Location-independent feature binding in visual working memory for sequentially presented objects. Atten. Percept. Psychophys. 2021, 83, 2377–2393. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cue Timing | Cue Content | Probe | Set Size | Mean | SD |
|---|---|---|---|---|---|
| Pre-Cue | Letters | Intact | 3 | 0.97 | 0.07 |
| 5 | 0.95 | 0.09 | |||
| New-Both | 3 | 0.97 | 0.07 | ||
| 5 | 0.96 | 0.08 | |||
| New-Relevant | 3 | 0.95 | 0.12 | ||
| 5 | 0.94 | 0.11 | |||
| Recombined | 3 | 0.97 | 0.06 | ||
| 5 | 0.93 | 0.12 | |||
| Locations | Intact | 3 | 0.80 | 0.18 | |
| 5 | 0.75 | 0.22 | |||
| New-Both | 3 | 0.86 | 0.19 | ||
| 5 | 0.82 | 0.19 | |||
| New-Relevant | 3 | 0.81 | 0.18 | ||
| 5 | 0.76 | 0.24 | |||
| Recombined | 3 | 0.74 | 0.25 | ||
| 5 | 0.74 | 0.21 | |||
| Retro-Cue | Letters | Intact | 3 | 0.96 | 0.10 |
| 5 | 0.93 | 0.10 | |||
| New-Both | 3 | 0.93 | 0.17 | ||
| 5 | 0.94 | 0.14 | |||
| New-Relevant | 3 | 0.96 | 0.10 | ||
| 5 | 0.94 | 0.10 | |||
| Recombined | 3 | 0.93 | 0.12 | ||
| 5 | 0.91 | 0.16 | |||
| Locations | Intact | 3 | 0.78 | 0.18 | |
| 5 | 0.75 | 0.18 | |||
| New-Both | 3 | 0.81 | 0.20 | ||
| 5 | 0.82 | 0.18 | |||
| New-Relevant | 3 | 0.80 | 0.16 | ||
| 5 | 0.72 | 0.21 | |||
| Recombined | 3 | 0.67 | 0.25 | ||
| 5 | 0.74 | 0.22 |
| Cue Timing | Cue Content | Probe | Set Size | Mean | SD |
|---|---|---|---|---|---|
| Pre-Cue | Letters | Intact | 3 | 1298 | 332 |
| 5 | 1500 | 497 | |||
| New-Both | 3 | 1401 | 478 | ||
| 5 | 1623 | 867 | |||
| New-Relevant | 3 | 1417 | 519 | ||
| 5 | 1576 | 548 | |||
| Recombined | 3 | 1339 | 413 | ||
| 5 | 1480 | 382 | |||
| Locations | Intact | 3 | 1503 | 646 | |
| 5 | 1656 | 771 | |||
| New-Both | 3 | 1258 | 332 | ||
| 5 | 1384 | 486 | |||
| New-Relevant | 3 | 1362 | 433 | ||
| 5 | 1429 | 525 | |||
| Recombined | 3 | 1585 | 627 | ||
| 5 | 1613 | 681 | |||
| Retro-Cue | Letters | Intact | 3 | 1368 | 344 |
| 5 | 1473 | 445 | |||
| New-Both | 3 | 1528 | 537 | ||
| 5 | 1665 | 695 | |||
| New-Relevant | 3 | 1484 | 696 | ||
| 5 | 1679 | 647 | |||
| Recombined | 3 | 1379 | 389 | ||
| 5 | 1526 | 336 | |||
| Locations | Intact | 3 | 1572 | 526 | |
| 5 | 1703 | 659 | |||
| New-Both | 3 | 1384 | 428 | ||
| 5 | 1554 | 505 | |||
| New-Relevant | 3 | 1405 | 381 | ||
| 5 | 1553 | 697 | |||
| Recombined | 3 | 1592 | 639 | ||
| 5 | 1720 | 688 |
| Block | Cue Content | Probe | Retention Interval | Mean | SD |
|---|---|---|---|---|---|
| Letter | Letter | intact | 1000 ms | 0.89 | 0.17 |
| 5000 ms | 0.87 | 0.20 | |||
| newLetter | 1000 ms | 0.95 | 0.10 | ||
| 5000 ms | 0.92 | 0.14 | |||
| newLocation | 1000 ms | 0.88 | 0.16 | ||
| 5000 ms | 0.91 | 0.15 | |||
| recombined | 1000 ms | 0.89 | 0.18 | ||
| 5000 ms | 0.88 | 0.17 | |||
| Location | Location | intact | 1000 ms | 0.85 | 0.22 |
| 5000 ms | 0.73 | 0.25 | |||
| newLetter | 1000 ms | 0.79 | 0.24 | ||
| 5000 ms | 0.70 | 0.28 | |||
| newLocation | 1000 ms | 0.85 | 0.16 | ||
| 5000 ms | 0.81 | 0.13 | |||
| recombined | 1000 ms | 0.78 | 0.25 | ||
| 5000 ms | 0.73 | 0.20 | |||
| Mixed | Letter | intact | 1000 ms | 0.90 | 0.19 |
| 5000 ms | 0.92 | 0.15 | |||
| newLetter | 1000 ms | 0.91 | 0.10 | ||
| 5000 ms | 0.90 | 0.13 | |||
| newLocation | 1000 ms | 0.89 | 0.19 | ||
| 5000 ms | 0.85 | 0.19 | |||
| recombined | 1000 ms | 0.90 | 0.18 | ||
| 5000 ms | 0.92 | 0.15 | |||
| Location | intact | 1000 ms | 0.86 | 0.21 | |
| 5000 ms | 0.77 | 0.21 | |||
| newLetter | 1000 ms | 0.76 | 0.23 | ||
| 5000 ms | 0.69 | 0.30 | |||
| newLocation | 1000 ms | 0.82 | 0.13 | ||
| 5000 ms | 0.80 | 0.18 | |||
| recombined | 1000 ms | 0.78 | 0.30 | ||
| 5000 ms | 0.72 | 0.27 |
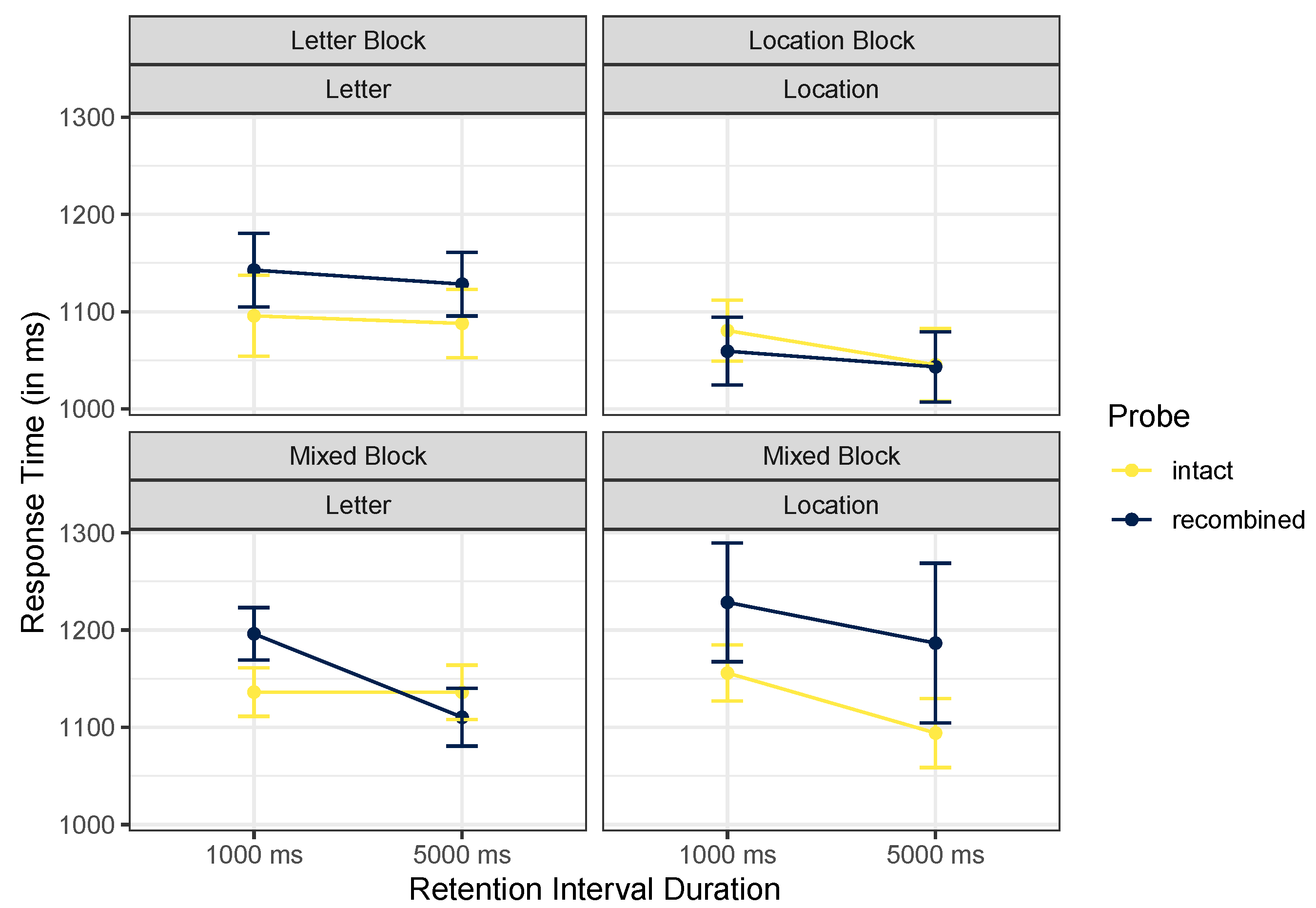
| Block | Cue Content | Probe | Retention Interval | Mean | SD |
|---|---|---|---|---|---|
| Letter Block | Letter | intact | 1000 ms | 1096 | 421 |
| 5000 ms | 1088 | 361 | |||
| newLetter | 1000 ms | 1199 | 370 | ||
| 5000 ms | 1136 | 359 | |||
| newLocation | 1000 ms | 1071 | 279 | ||
| 5000 ms | 1042 | 287 | |||
| recombined | 1000 ms | 1143 | 462 | ||
| 5000 ms | 1129 | 359 | |||
| Location Block | Location | intact | 1000 ms | 1079 | 316 |
| 5000 ms | 1042 | 308 | |||
| newLetter | 1000 ms | 1062 | 316 | ||
| 5000 ms | 1080 | 397 | |||
| newLocation | 1000 ms | 1061 | 287 | ||
| 5000 ms | 985 | 263 | |||
| recombined | 1000 ms | 1061 | 320 | ||
| 5000 ms | 1044 | 290 | |||
| Mixed Block | Letter | intact | 1000 ms | 1137 | 282 |
| 5000 ms | 1137 | 318 | |||
| newLetter | 1000 ms | 1186 | 282 | ||
| 5000 ms | 1182 | 346 | |||
| newLocation | 1000 ms | 1219 | 342 | ||
| 5000 ms | 1155 | 407 | |||
| recombined | 1000 ms | 1197 | 379 | ||
| 5000 ms | 1109 | 337 | |||
| Location | intact | 1000 ms | 1156 | 385 | |
| 5000 ms | 1095 | 315 | |||
| newLetter | 1000 ms | 1232 | 568 | ||
| 5000 ms | 1082 | 344 | |||
| newLocation | 1000 ms | 1114 | 342 | ||
| 5000 ms | 1067 | 310 | |||
| recombined | 1000 ms | 1224 | 645 | ||
| 5000 ms | 1190 | 825 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delooze, M.A.; Langerock, N.; Macy, R.; Vergauwe, E.; Morey, C.C. Encode a Letter and Get Its Location for Free? Assessing Incidental Binding of Verbal and Spatial Features. Brain Sci. 2022, 12, 685. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12060685
Delooze MA, Langerock N, Macy R, Vergauwe E, Morey CC. Encode a Letter and Get Its Location for Free? Assessing Incidental Binding of Verbal and Spatial Features. Brain Sciences. 2022; 12(6):685. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12060685
Chicago/Turabian StyleDelooze, Molly A., Naomi Langerock, Robin Macy, Evie Vergauwe, and Candice C. Morey. 2022. "Encode a Letter and Get Its Location for Free? Assessing Incidental Binding of Verbal and Spatial Features" Brain Sciences 12, no. 6: 685. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12060685