Functional Connectivity between the Resting-State Olfactory Network and the Hippocampus in Alzheimer’s Disease

Abstract
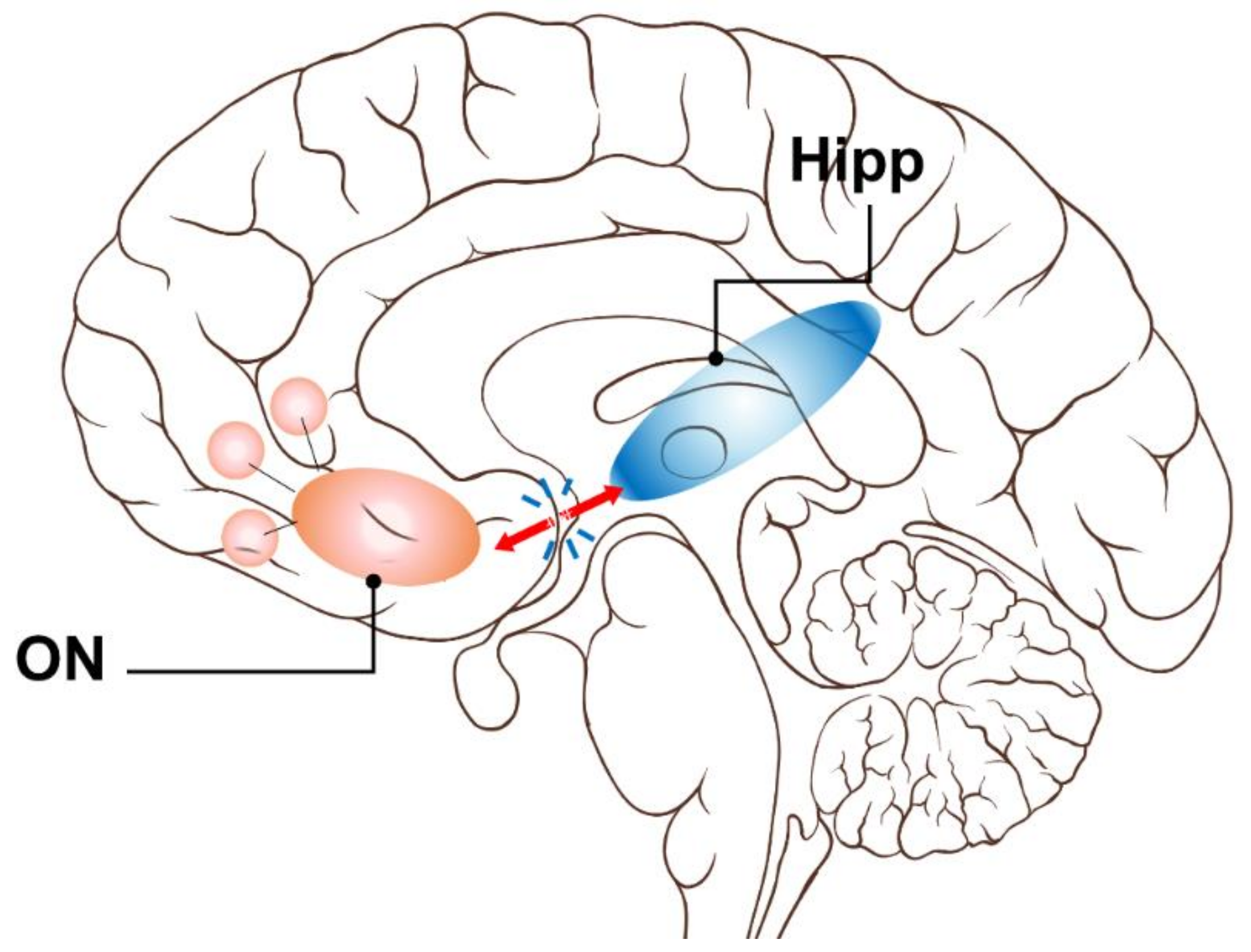
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Image Preprocessing
2.3. Olfactory Network
2.4. Statistical Analysis
2.5. Volumetric Analysis
2.6. Relationship between ON FC and Neuropsychological Test Scores
3. Results
3.1. Demographic and Neuropsychological Data
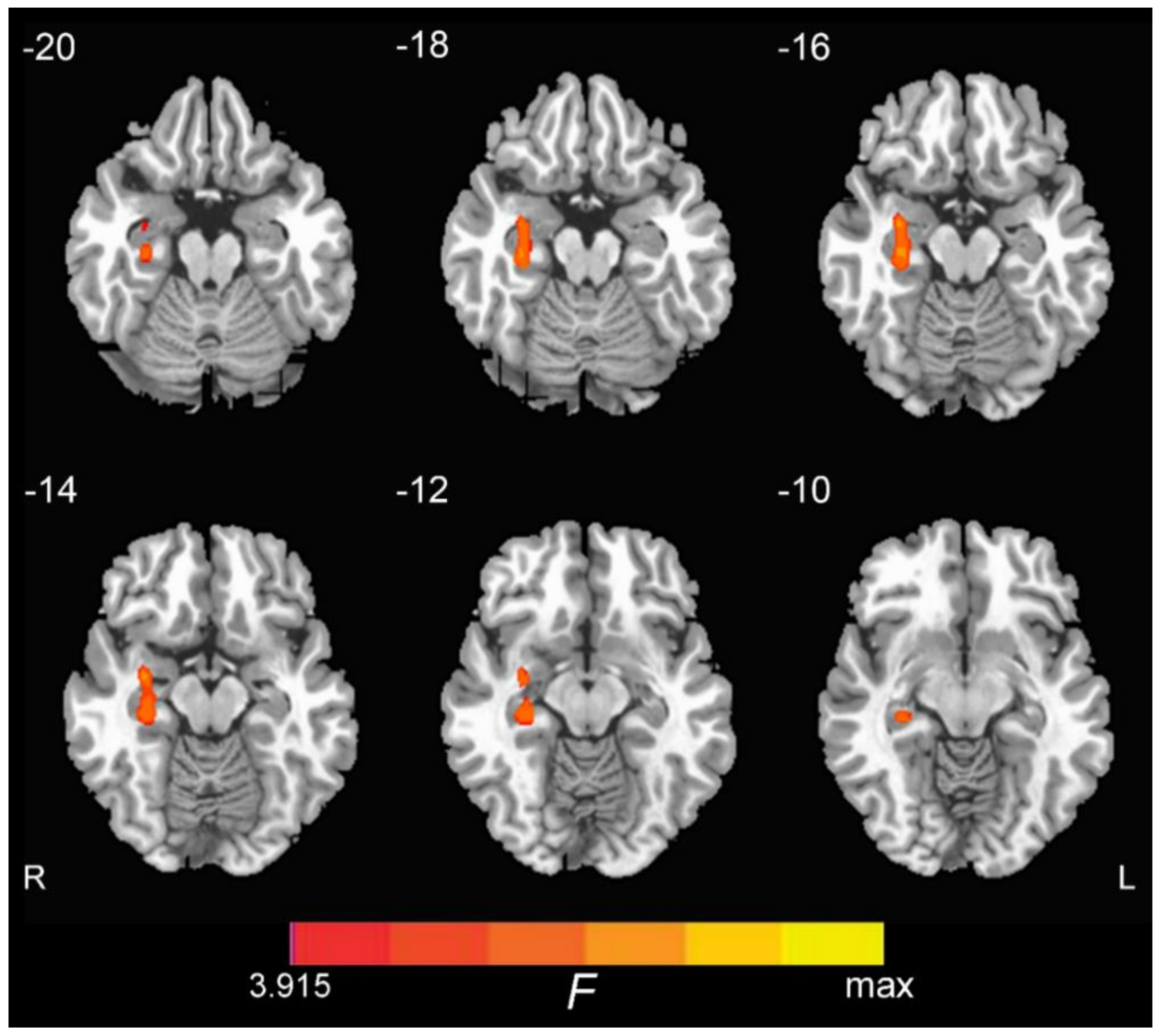
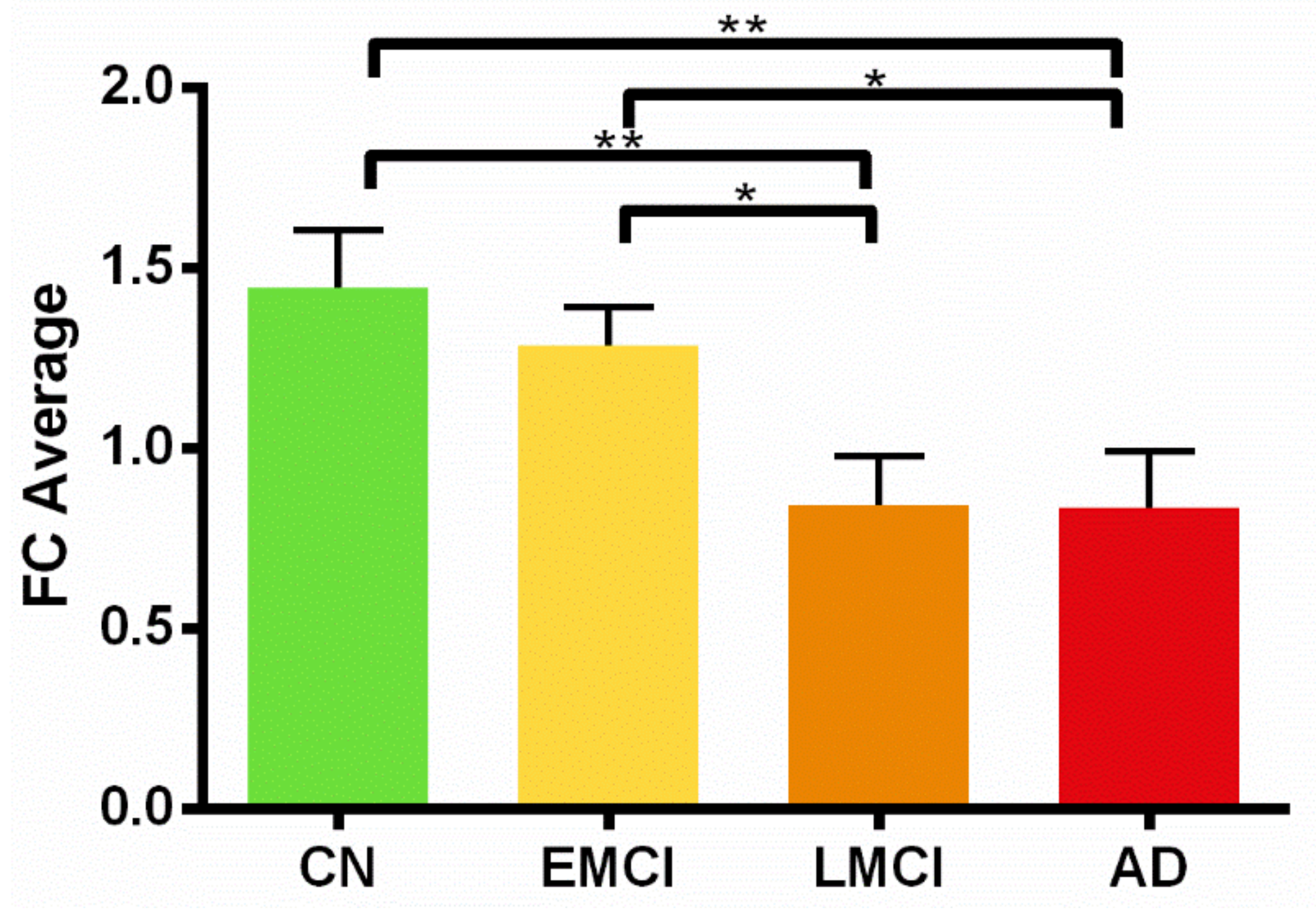
3.2. ON FC Differences and the Region of Interest (ROI) Analysis
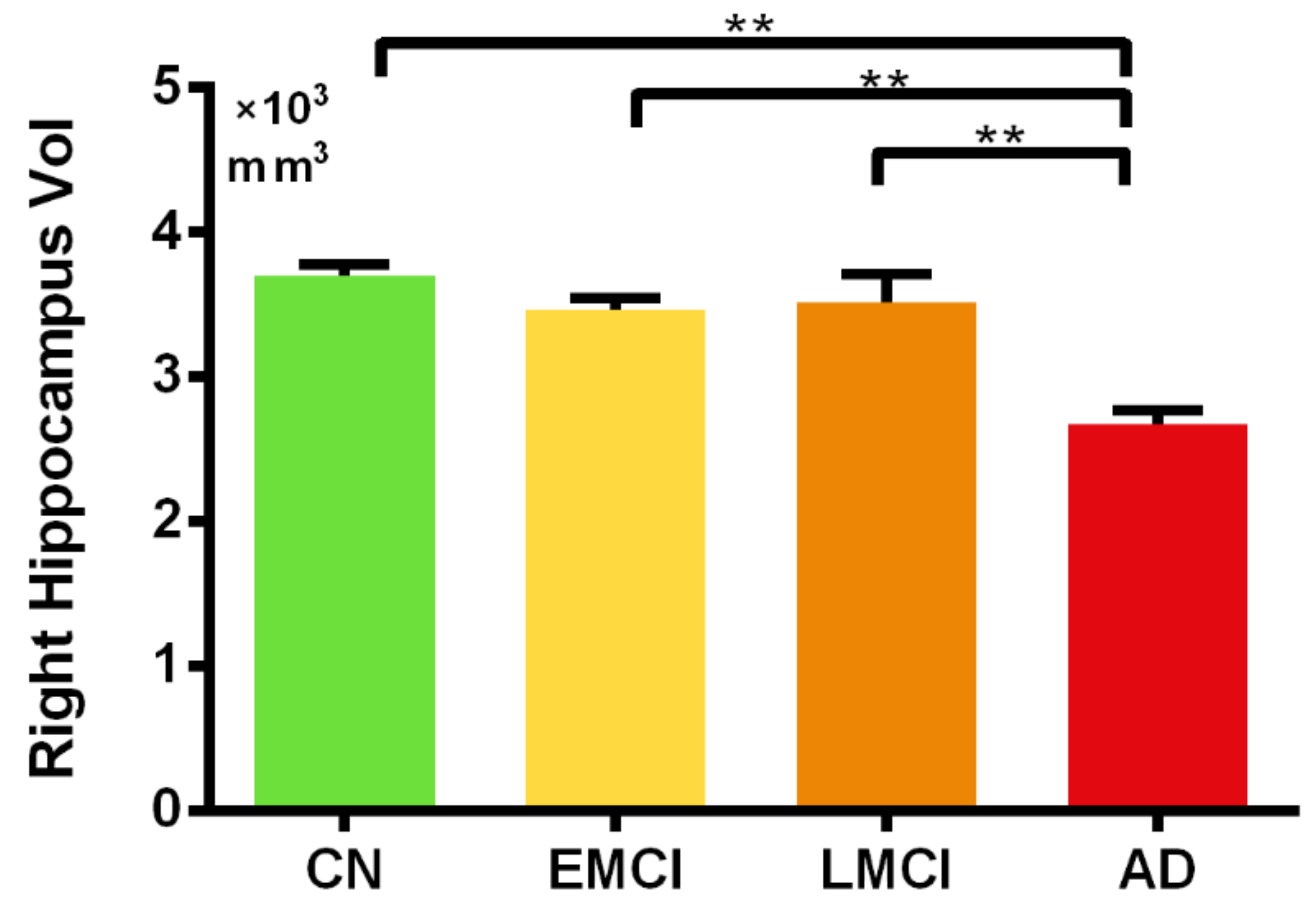
3.3. Region of Interest (ROI) Analysis of the Hippocampal Volume
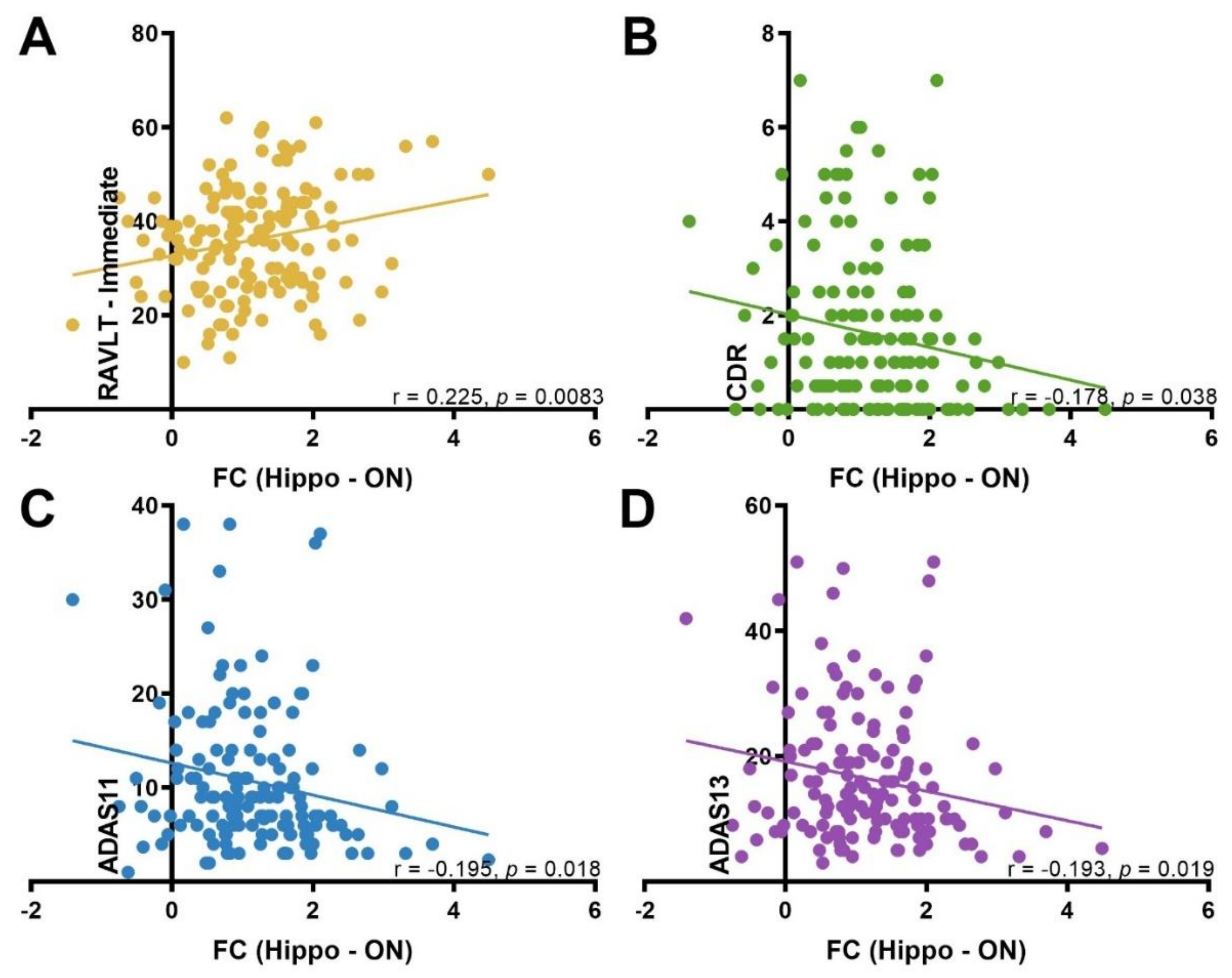
3.4. Correlation between ON FC and Neuropsychological Test Scores
4. Discussion
Limitations of the Study
5. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Murphy, C. Olfactory and other sensory impairments in Alzheimer disease. Nat. Rev. Neurol. 2019, 15, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Albers, M.W.; Gilmore, G.C.; Kaye, J.; Murphy, C.; Wingfield, A.; Bennett, D.A.; Boxer, A.L.; Buchman, A.S.; Cruickshanks, K.J.; Devanand, D.P.; et al. At the interface of sensory and motor dysfunctions and Alzheimer’s disease. Alzheimers Dement. 2015, 11, 70–98. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.E.; Cruickshanks, K.J.; Schubert, C.R.; Pinto, A.A.; Carlsson, C.M.; Klein, B.E.; Klein, R.; Tweed, T.S. Age-Related Sensory Impairments and Risk of Cognitive Impairment. J. Am. Geriatr. Soc. 2016, 64, 1981–1987. [Google Scholar] [CrossRef] [PubMed]
- Davies, H.R.; Cadar, D.; Herbert, A.; Orrell, M.; Steptoe, A. Hearing Impairment and Incident Dementia: Findings from the English Longitudinal Study of Ageing. J. Am. Geriatr. Soc. 2017, 65, 2074–2081. [Google Scholar] [CrossRef]
- Deal, J.A.; Betz, J.; Yaffe, K.; Harris, T.; Purchase-Helzner, E.; Satterfield, S.; Pratt, S.; Govil, N.; Simonsick, E.M.; Lin, F.R. Hearing Impairment and Incident Dementia and Cognitive Decline in Older Adults: The Health ABC Study. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 703–709. [Google Scholar] [CrossRef]
- Hyman, B.T. The neuropathological diagnosis of Alzheimer’s disease: Clinical-pathological studies. Neurobiol. Aging 1997, 18 (Suppl. 4), S27–S32. [Google Scholar] [CrossRef]
- Waldton, S. Clinical observations of impaired cranial nerve function in senile dementia. Acta Psychiatr. Scand. 1974, 50, 539–547. [Google Scholar] [CrossRef]
- Doty, R.L.; Reyes, P.F.; Gregor, T. Presence of both odor identification and detection deficits in Alzheimer’s disease. Brain Res. Bull. 1987, 18, 597–600. [Google Scholar] [CrossRef]
- Ferreyra-Moyano, H.; Barragan, E. The olfactory system and Alzheimer’s disease. Int. J. Neurosci. 1989, 49, 157–197. [Google Scholar] [CrossRef]
- Knupfer, L.; Spiegel, R. Differences in olfactory test performance between normal aged, Alzheimer and vascular type dementia individuals. Int. J. Geriatr. Psychiatry 1986, 1, 3–14. [Google Scholar] [CrossRef]
- Murphy, C.; Gilmore, M.M.; Seery, C.S.; Salmon, D.P.; Lasker, B.R. Olfactory thresholds are associated with degree of dementia in Alzheimer’s disease. Neurobiol. Aging 1990, 11, 465–469. [Google Scholar] [CrossRef]
- Roberts, R.O.; Christianson, T.J.; Kremers, W.K.; Mielke, M.M.; Machulda, M.M.; Vassilaki, M.; Alhurani, R.E.; Geda, Y.E.; Knopman, D.S.; Petersen, R.C. Association between olfactory dysfunction and amnestic mild cognitive impairment and Alzheimer disease dementia. JAMA Neurol. 2016, 73, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Roalf, D.R.; Moberg, M.J.; Turetsky, B.I.; Brennan, L.; Kabadi, S.; Wolk, D.A.; Moberg, P.J. A quantitative meta-analysis of olfactory dysfunction in mild cognitive impairment. A quantitative meta-analysis of olfactory dysfunction in mild cognitive impairment. J. Neurol. Neurosurg. Psychiatry 2017, 88, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Rahayel, S.; Frasnelli, J.; Joubert, S. The effect of Alzheimer’s disease and Parkinson’s disease on olfaction: A meta-analysis. Behav. Brain Res. 2012, 231, 60–74. [Google Scholar] [CrossRef]
- Wilson, R.S.; Arnold, S.E.; Schneider, J.A.; Tang, Y.; Bennett, D.A. The relationship between cerebral Alzheimer’s disease pathology and odour identification in old age. J. Neurol. Neurosurg. Psychiatry 2007, 78, 30–35. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Evolution of the neuropathology of Alzheimer’s disease. Acta Neurol. Scand. Suppl. 1996, 165, 3–12. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Evolution of neuronal changes in the course of Alzheimer’s disease. J. Neural. Transm. Suppl. 1998, 53, 127–140. [Google Scholar]
- Braak, H.; Thal, D.R.; Ghebremedhin, E.; Del Tredici, K. Stages of the pathologic process in Alzheimer disease: Age categories from 1 to 100 years. J. Neuropathol. Exp. Neurol. 2011, 70, 960–969. [Google Scholar] [CrossRef]
- Sheline, Y.I.; Raichle, M.E. Resting state functional connectivity in preclinical Alzheimer’s disease. Biol. Psychiatry 2013, 74, 340–347. [Google Scholar] [CrossRef]
- Wang, L.; Zang, Y.; He, Y.; Liang, M.; Zhang, X.; Tian, L.; Wu, T.; Jiang, T.; Li, K. Changes in hippocampal connectivity in the early stages of Alzheimer’s disease: Evidence from resting state fMRI. Neuroimage 2006, 31, 496–504. [Google Scholar] [CrossRef]
- Greicius, M.D.; Srivastava, G.; Reiss, A.L.; Menon, V. Default-mode network activity distinguishes Alzheimer’s disease from healthy aging: Evidence from functional MRI. Proc. Natl. Acad. Sci. USA 2004, 101, 4637–4642. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Li, Z.; Wu, G.; Zhang, M.J.; Franczak, M.; Antuono, P.G. Alzheimer Disease: Evaluation of a functional MR imaging index as a marker. Radiology 2002, 225, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Deco, G.; Jirsa, V.K.; McIntosh, A.R. Emerging concepts for the dynamical organization of resting-state activity in the brain. Nat. Rev. Neurosci. 2011, 12, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.W.; Bassett, D.S.; Power, J.D.; Braver, T.S.; Petersen, S.E. Intrinsic and task-evoked network architectures of the human brain. Neuron 2014, 83, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Osada, T.; Sporns, O.; Watanabe, T.; Matsui, T.; Miyamoto, K.; Miyashita, Y. Functional connectivity between anatomically unconnected areas is shaped by collective network-level effects in the macaque cortex. Cereb. Cortex 2012, 22, 1586–1592. [Google Scholar] [CrossRef]
- Goni, J.; van den Heuvel, M.P.; Avena-Koenigsberger, A.; Velez de Mendizabal, N.; Betzel, R.F.; Griffa, A.; Hagmann, P.; Corominas-Murtra, B.; Thiran, J.P.; Sporns, O. Resting-brain functional connectivity predicted by analytic measures of network communication. Proc. Natl. Acad. Sci. USA 2014, 111, 833–838. [Google Scholar] [CrossRef]
- Tobia, M.J.; Yang, Q.X.; Karunanayaka, P. Intrinsic intranasal chemosensory brain networks shown by resting-state functional MRI. Neuroreport 2016, 27, 527–531. [Google Scholar] [CrossRef]
- Gottfried, J.A.; Dolan, R.J. The nose smells what the eye sees: Crossmodal visual facilitation of human olfactory perception. Neuron 2003, 39, 375–386. [Google Scholar] [CrossRef]
- Doty, R.L. Olfactory capacities in aging and Alzheimer’s disease. Psychophysical and anatomic considerations. Ann. N. Y. Acad. Sci. 1991, 640, 20–27. [Google Scholar] [CrossRef]
- Vasavada, M.M.; Wang, J.; Eslinger, P.J.; Gill, D.J.; Sun, X.; Karunanayaka, P.; Yang, Q.X. Olfactory cortex degeneration in Alzheimer’s disease and mild cognitive impairment. J. Alzheimers Dis. 2015, 45, 947–958. [Google Scholar] [CrossRef]
- Tabert, M.H.; Steffener, J.; Albers, M.W.; Kern, D.W.; Michael, M.; Tang, H.; Brown, T.R.; Devanand, D.P. Validation and optimization of statistical approaches for modeling odorant-induced fMRI signal changes in olfactory-related brain areas. Neuroimage 2007, 34, 1375–1390. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, K.; Freimer, D.; Chen, H.; Asao, K.; Rosso, A.; Rubin, S.; Tranah, G.; Cummings, S.; Simonsick, E. Olfaction and risk of dementia in a biracial cohort of older adults. Neurology 2017, 88, 456–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiner, M.W.; Aisen, P.S.; Jack Jr, C.R.; Jagust, W.J.; Trojanowski, J.Q.; Shaw, L.; Saykin, A.J.; Morris, J.C.; Cairns, N.; Beckett, L.A.J.A.s.; et al. The Alzheimer’s disease neuroimaging initiative: Progress report and future plans. Alzheimers Dement. 2010, 6, 202–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, C.; Zang, Y. DPARSF: A MATLAB toolbox for “pipeline” data analysis of resting-state fMRI. Front. Syst. Neurosci. 2010, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.G.; Wang, X.D.; Zuo, X.N.; Zang, Y.F. DPABI: Data Processing & Analysis for (Resting-State) Brain Imaging. Neuroinformatics 2016, 14, 339–351. [Google Scholar]
- Anderson, J.S.; Druzgal, T.J.; Lopez-Larson, M.; Jeong, E.K.; Desai, K.; Yurgelun-Todd, D. Network anticorrelations, global regression, and phase-shifted soft tissue correction. Hum. Brain Mapp. 2011, 32, 919–934. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.; Birn, R.M.; Handwerker, D.A.; Jones, T.B.; Bandettini, P.A. The impact of global signal regression on resting state correlations: Are anti-correlated networks introduced? Neuroimage 2009, 44, 893–905. [Google Scholar] [CrossRef] [Green Version]
- Seubert, J.; Freiherr, J.; Djordjevic, J.; Lundstrom, J.N. Statistical localization of human olfactory cortex. Neuroimage 2013, 66, 333–342. [Google Scholar] [CrossRef]
- Devanand, D.P.; Tabert, M.H.; Cuasay, K.; Manly, J.J.; Schupf, N.; Brickman, A.M.; Andrews, H.; Brown, T.R.; DeCarli, C.; Mayeux, R. Olfactory identification deficits and MCI in a multi-ethnic elderly community sample. Neurobiol. Aging 2010, 31, 1593–1600. [Google Scholar] [CrossRef] [Green Version]
- Jack, C.R., Jr.; Knopman, D.S.; Jagust, W.J.; Shaw, L.M.; Aisen, P.S.; Weiner, M.W.; Petersen, R.C.; Trojanowski, J.Q. Hypothetical model of dynamic biomarkers of the Alzheimer’s pathological cascade. Lancet Neurol. 2010, 9, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Hirano, A.; Zimmerman, H.M. Alzheimer’s neurofibrillary changes: A topographic study. Arch. Neurol. 1962, 7, 227–242. [Google Scholar] [CrossRef] [PubMed]
- De Lacoste, M.C.; White, C.L., 3rd. The role of cortical connectivity in Alzheimer’s disease pathogenesis: A review and model system. Neurobiol. Aging 1993, 14, 1–16. [Google Scholar] [CrossRef]
- Brier, M.R.; Thomas, J.B.; Ances, B.M. Network dysfunction in Alzheimer’s disease: Refining the disconnection hypothesis. Brain Connect. 2014, 4, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Morphological criteria for the recognition of Alzheimer’s disease and the distribution pattern of cortical changes related to this disorder. Neurobiol. Aging 1994, 15, 355–356. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Frequency of stages of Alzheimer-related lesions in different age categories. Neurobiol. Aging 1997, 18, 351–357. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. The human entorhinal cortex: Normal morphology and lamina-specific pathology in various diseases. Neurosci. Res. 1992, 15, 6–31. [Google Scholar] [CrossRef]
- Hyman, B.T.; Van Hoesen, G.W.; Damasio, A.R.; Barnes, C.L. Alzheimer’s disease: Cell-specific pathology isolates the hippocampal formation. Science 1984, 225, 1168–1170. [Google Scholar] [CrossRef]
- Hyman, B.T.; Van Hoesen, G.W.; Kromer, L.J.; Damasio, A.R. Perforant pathway changes and the memory impairment of Alzheimer’s disease. Ann. Neurol. 1986, 20, 472–481. [Google Scholar] [CrossRef]
- Delbeuck, X.; van der Linden, M.; Collette, F. Alzheimer’s disease as a disconnection syndrome? Neuropsychol. Rev. 2003, 13, 79–92. [Google Scholar] [CrossRef]
- Zuo, X.N.; Ehmke, R.; Mennes, M.; Imperati, D.; Castellanos, F.X.; Sporns, O.; Milham, M.P. Network centrality in the human functional connectome. Cereb. Cortex 2012, 22, 1862–1875. [Google Scholar] [CrossRef]
- Lacalle-Aurioles, M.; Navas-Sanchez, F.J.; Aleman-Gomez, Y.; Olazaran, J.; Guzman-De-Villoria, J.A.; Cruz-Orduna, I.; Mateos-Perez, J.M.; Desco, M. The Disconnection Hypothesis in Alzheimer’s Disease Studied Through Multimodal Magnetic Resonance Imaging: Structural, Perfusion, and Diffusion Tensor Imaging. J. Alzheimers Dis. 2016, 50, 1051–1064. [Google Scholar] [CrossRef]
- Allen, G.; Barnard, H.; McColl, R.; Hester, A.L.; Fields, J.A.; Weiner, M.F.; Ringe, W.K.; Lipton, A.M.; Brooker, M.; McDonald, E.; et al. Reduced hippocampal functional connectivity in Alzheimer disease. Arch. Neurol. 2007, 64, 1482–1487. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.Y.; Wang, S.J.; Xing, J.; Liu, B.; Ma, Z.L.; Yang, M.; Zhang, Z.J.; Teng, G.J. Detection of PCC functional connectivity characteristics in resting-state fMRI in mild Alzheimer’s disease. Behav. Brain Res. 2009, 197, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Karunanayaka, P.R.; Wilson, D.A.; Tobia, M.J.; Martinez, B.E.; Meadowcroft, M.D.; Eslinger, P.J.; Yang, Q.X. Default mode network deactivation during odor—Visual association. Hum. Brain Mapp. 2017, 38, 1125–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Yang, Q.X.; Zhang, H.; Eslinger, P.J.; Zhang, X.; Wu, S.; Zhang, B.; Zhu, B.; Karunanayaka, P.R. Disruptions of the olfactory and default mode networks in Alzheimer’s disease. Brain Behav. 2019, 9, e01296. [Google Scholar] [CrossRef]
- Filippi, M.; Basaia, S.; Canu, E.; Imperiale, F.; Meani, A.; Caso, F.; Magnani, G.; Falautano, M.; Comi, G.; Falini, A.; et al. Brain network connectivity differs in early-onset neurodegenerative dementia. Neurology 2017, 89, 1764–1772. [Google Scholar] [CrossRef] [Green Version]
- Hamalainen, A.; Pihlajamaki, M.; Tanila, H.; Hanninen, T.; Niskanen, E.; Tervo, S.; Karjalainen, P.A.; Vanninen, R.L.; Soininen, H. Increased fMRI responses during encoding in mild cognitive impairment. Neurobiol. Aging 2007, 28, 1889–1903. [Google Scholar] [CrossRef]
- Heun, R.; Freymann, K.; Erb, M.; Leube, D.T.; Jessen, F.; Kircher, T.T.; Grodd, W. Mild cognitive impairment (MCI) and actual retrieval performance affect cerebral activation in the elderly. Neurobiol. Aging 2007, 28, 404–413. [Google Scholar] [CrossRef]
- Dickerson, B.C.; Sperling, R.A. Large-scale functional brain network abnormalities in Alzheimer’s disease: Insights from functional neuroimaging. Behav. Neurol. 2009, 21, 63–75. [Google Scholar] [CrossRef]
- Machulda, M.M.; Ward, H.A.; Borowski, B.; Gunter, J.L.; Cha, R.H.; O’Brien, P.C.; Petersen, R.C.; Boeve, B.F.; Knopman, D.; Tang-Wai, D.F.; et al. Comparison of memory fMRI response among normal, MCI, and Alzheimer’s patients. Neurology 2003, 61, 500–506. [Google Scholar] [CrossRef]
- Bakker, A.; Krauss, G.L.; Albert, M.S.; Speck, C.L.; Jones, L.R.; Stark, C.E.; Yassa, M.A.; Bassett, S.S.; Shelton, A.L.; Gallagher, M. Reduction of hippocampal hyperactivity improves cognition in amnestic mild cognitive impairment. Neuron 2012, 74, 467–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zott, B.; Simon, M.M.; Hong, W.; Unger, F.; Chen-Engerer, H.J.; Frosch, M.P.; Sakmann, B.; Walsh, D.M.; Konnerth, A. A vicious cycle of beta amyloid-dependent neuronal hyperactivation. Science 2019, 365, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Postuma, R.B.; Gagnon, J.F.; Vendette, M.; Desjardins, C.; Montplaisir, J.Y. Olfaction and color vision identify impending neurodegeneration in rapid eye movement sleep behavior disorder. Ann. Neurol. 2011, 69, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage 2012, 59, 2142–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Hao, C.; Jiang, J.; Feng, Y.; Chen, X.; Zheng, Y.; Liu, J.; Zhang, Z.; Long, C.; Yang, L. The mechanisms underlying olfactory deficits in apolipoprotein E-deficient mice: Focus on olfactory epithelium and olfactory bulb. Neurobiol. Aging 2018, 62, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.C. Mild cognitive impairment. Lancet 2006, 367, 1979. [Google Scholar] [CrossRef] [Green Version]
- Lafaille-Magnan, M.E.; Poirier, J.; Etienne, P.; Tremblay-Mercier, J.; Frenette, J.; Rosa-Neto, P.; Breitner, J.C.S. Odor identification as a biomarker of preclinical AD in older adults at risk. Neurology 2017, 89, 327–335. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CN (n = 44) | EMCI (n = 46) | LMCI (n = 31) | AD (n = 26) | p # | |
|---|---|---|---|---|---|
| Male, No. (%) | 17 (38.6) | 19 (41.3) | 18 (58.1) | 11 (42.3) | 0.518 |
| Age | 74.18 (6.1) | 71.69 (7.3) | 72.41 (7.4) | 71.55 (7.3) | 0.37 |
| Education | 16.50 (2.7) | 15.65 (2.6) | 16.90 (2.3) | 15.31 (2.8) | 0.052 |
| MMSE | 28.86 (1.4) | 28.39 (1.6) | 27.74 (1.6) | 22.54 (2.6) | <0.001 |
| MoCA | 25.68 (2.1) | 24.00 (2.8) | 22.47 (3.2) | 15.88 (5.7) | <0.001 |
| CDR | 0.045 (0.2) | 1.42 (0.9) | 1.73 (0.93) | 4.46 (1.4) | <0.001 |
| RAVLT | 44.23 (8.1) | 37.93 (10.3) | 33.16 (7.4) | 22.46 (7.7) | <0.001 |
| ADAS11 | 5.69 (2.1) | 7.93 (3.4) | 11.09 (4.7) | 23.19 (8.2) | <0.001 |
| ADAS13 | 9.19 (3.7) | 12.52 (5.2) | 17.55 (7.0) | 34.23 (9.7) | <0.001 |
| FAQ | 0.11 (0.5) | 2.48(3.9) | 4.871 (4.9) | 15.038 (7.4) | <0.001 |
| CN vs. EMCI | CN vs. LMCI | CN vs. AD | EMCI vs. LMCI | EMCI vs. AD | LMCI vs. AD | |
|---|---|---|---|---|---|---|
| p | ||||||
| Education | 0.095 | 0.632 | 0.048 | 0.046 | <0.001 | 0.024 |
| MMSE | 0.191 | 0.007 | <0.001 | 0.125 | <0.001 | <0.001 |
| MoCA | 0.018 | <0.001 | <0.001 | 0.053 | <0.001 | 0.001 |
| CDR | <0.001 | <0.001 | <0.001 | 0.344 | <0.001 | <0.001 |
| RAVLT | <0.001 | <0.001 | <0.001 | 0.058 | <0.001 | <0.001 |
| ADAS11 | 0.014 | <0.001 | <0.001 | 0.011 | <0.001 | <0.001 |
| ADAS13 | 0.017 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| FAQ | <0.001 | <0.001 | <0.001 | 0.011 | <0.001 | <0.001 |
| Cluster | Cluster Size (Voxel) | MNI Coordinates | t Value | ||
|---|---|---|---|---|---|
| x | y | z | |||
| Right Hippocampus | 74 | 33 | −6 | −15 | 7.59 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, J.; Testa, N.; Jordan, R.; Elyan, R.; Kanekar, S.; Wang, J.; Eslinger, P.; Yang, Q.X.; Zhang, B.; Karunanayaka, P.R. Functional Connectivity between the Resting-State Olfactory Network and the Hippocampus in Alzheimer’s Disease. Brain Sci. 2019, 9, 338. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9120338
Lu J, Testa N, Jordan R, Elyan R, Kanekar S, Wang J, Eslinger P, Yang QX, Zhang B, Karunanayaka PR. Functional Connectivity between the Resting-State Olfactory Network and the Hippocampus in Alzheimer’s Disease. Brain Sciences. 2019; 9(12):338. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9120338
Chicago/Turabian StyleLu, Jiaming, Nicole Testa, Rebecca Jordan, Rommy Elyan, Sangam Kanekar, Jianli Wang, Paul Eslinger, Qing X. Yang, Bing Zhang, and Prasanna R. Karunanayaka. 2019. "Functional Connectivity between the Resting-State Olfactory Network and the Hippocampus in Alzheimer’s Disease" Brain Sciences 9, no. 12: 338. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9120338