Acute Effects of Oatmeal on Exercise-Induced Reactive Oxygen Species Production Following High-Intensity Interval Training in Women: A Randomized Controlled Trial

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
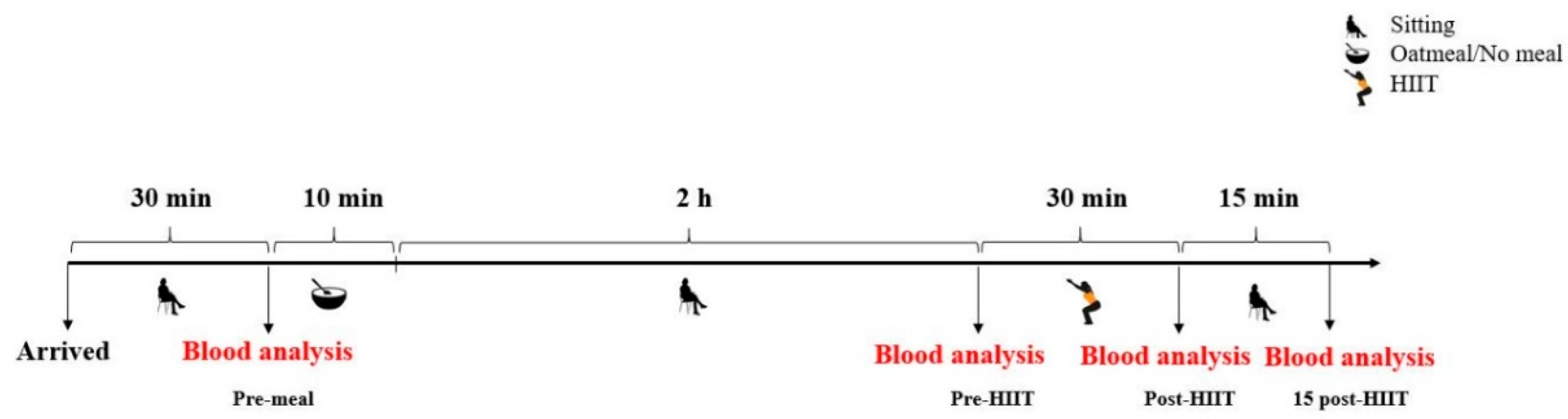
2.2. Study Design
2.3. High-Intensity Interval Training (HIIT)
2.4. Oatmeal
2.5. Blood Analysis
2.6. Body Composition
2.7. Borg Scale of Perceived Exertion
2.8. Statistical Analysis
3. Results
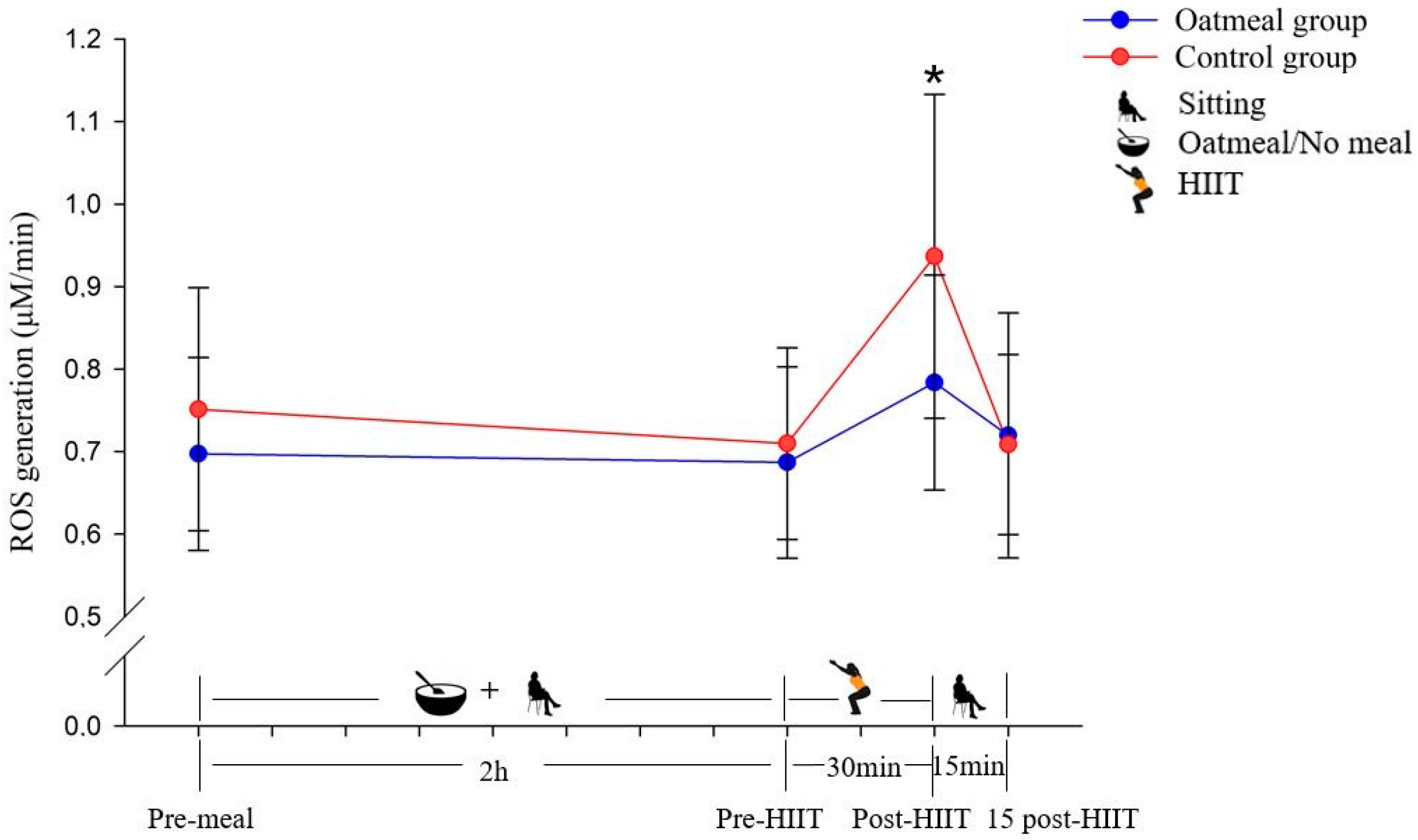
3.1. Reactive Oxygen Species Production
3.2. Glucose
3.3. Blood Lactate
3.4. Borg Scale
4. Discussion
5. Practical Application
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bloomer, R.J. Effect of Exercise on Oxidative Stress Biomarkers. Adv. Clin. Chem. 2008, 46, 1–50. [Google Scholar] [CrossRef] [PubMed]
- Hoidal, J.R. Reactive Oxygen Species and Cell Signaling. Am. J. Respir. Cell Mol. Biol. 2001, 25, 661–663. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, E.; Sestili, P. Reactive Oxygen Species in Skeletal Muscle Signaling. J. Signal. Transduct. 2012, 2012, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pingitore, A.; Lima, G.P.P.; Mastorci, F.; Quinones, A.; Iervasi, G.; Vassalle, C. Exercise and Oxidative Stress: Potential Effects of Antioxidant Dietary Strategies in Sports. Nutrition 2015, 31, 916–922. [Google Scholar] [CrossRef]
- Hudson, M.B.; Hosick, P.A.; Mccaulley, G.O.; Schrieber, L.; Wrieden, J.; Mcanulty, S.R.; Triplett, N.T.; Mcbride, J.M.; Quindry, J.C. The Effect of Resistance Exercise on Humoral Markers of Oxidative Stress. Med. Sci. Sports Exerc. 2008, 40, 542–548. [Google Scholar] [CrossRef]
- Owens, D.J.; Twist, C.; Cobley, J.N.; Howatson, G.; Close, G.L. Exercise-Induced Muscle Damage: What Is It, What Causes It and What Are the Nutritional Solutions? Eur. J. Sport Sci. 2019, 19, 71–85. [Google Scholar] [CrossRef]
- Nikolaidis, M.G.; Jamurtas, A.Z.; Paschalis, V.; Fatouros, I.G.; Koutedakis, Y.; Kouretas, D. The Effect of Muscle-Damaging Exercise on Blood and Skeletal Muscle Oxidative Stress: Magnitude and Time-Course Considerations. Sports Med. 2008, 38, 579–606. [Google Scholar] [CrossRef]
- Harper, M.E.; Bevilacqua, L.; Hagopian, K.; Weindruch, R.; Ramsey, J.J. Ageing, Oxidative Stress, and Mitochondrial Uncoupling. Acta. Physiol. Scand. 2004, 182, 321–331. [Google Scholar] [CrossRef]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, Oxidants, and Aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic Inflammation and Oxidative Stress as a Major Cause of Age- Related Diseases and Cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 73–80. [Google Scholar] [CrossRef]
- Di Meo, S.; Napolitano, G.; Venditti, P. Mediators of Physical Activity Protection against ROS-Linked Skeletal Muscle Damage. Int. J. Mol. Sci. 2019, 20, 3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Brown, A.M.; Frontera, W.R. Principles of Exercise Physiology: Responses to Acute Exercise and Long-term Adaptations to Training. PM&R 2012, 4, 797–804. [Google Scholar] [CrossRef]
- McArdle, A.; Pattwell, D.; Vasilaki, A.; Griffiths, R.D.; Jackson, M.J. Contractile activity-induced oxidative stress: Cellular origin and adaptive responses. Am. J. Physiol. Cell Physiol. 2001, 280, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Alessio, H.M.; Goldfarb, A.H.; Cao, G. Exercise-Induced Oxidative Stress before and after Vitamin C Supplementation. Int. J. Sport Nutr. 1997, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Ohkuwa, T.; Yamazaki, Y.; Shimoda, T.; Wakayama, A.; Tamura, S.; Yamamoto, T.; Sato, Y.; Miyamura, M. Vitamin E Supplementation Attenuates Leakage of Enzymes Following 6 Successive Days of Running Training. Int. J. Sports Med. 2000, 21, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Lázaro, D.; Fernandez-Lazaro, C.I.; Mielgo-Ayuso, J.; Navascués, L.J.; Martínez, A.C.; Seco-Calvo, J. The Role of Selenium Mineral Trace Element in Exercise: Antioxidant Defense System, Muscle Performance, Hormone Response, and Athletic Performance. A Systematic Review. Nutrients 2020, 12, 1790. [Google Scholar] [CrossRef]
- Bloomer, R.J.; Goldfarb, A.H.; McKenzie, M.J. Oxidative Stress Response to Aerobic Exercise: Comparison of Antioxidant Supplements. Med. Sci. Sports Exerc. 2006, 38, 1098–1105. [Google Scholar] [CrossRef] [Green Version]
- Koenig, R.T.; Dickman, J.R.; Kang, C.H.; Zhang, T.; Chu, Y.F.; Ji, L.L. Avenanthramide Supplementation Attenuates Eccentric Exercise-Inflicted Blood Inflammatory Markers in Women. Eur. J. Appl. Physiol. 2016, 116, 67–76. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, T.; Zhang, Y.; Liu, T.; Gagnon, G.; Ebrahim, J.; Johnson, J.; Chu, Y.F.; Ji, L.L. Avenanthramide Supplementation Reduces Eccentric Exercise-Induced Inflammation in Young Men and Women. J. Int. Soc. Sports Nutr. 2020, 17, 1–12. [Google Scholar] [CrossRef]
- Ji, L.L.; Lay, D.; Chung, E.; Fu, Y.; Peterson, D.M. Effects of Avenanthramides on Oxidant Generation and Antioxidant Enzyme Activity in Exercised Rats. Nutr. Res. 2003, 23, 1579–1590. [Google Scholar] [CrossRef]
- Koenig, R.; Dickman, J.R.; Kang, C.; Zhang, T.; Chu, Y.F.; Ji, L.L. Avenanthramide Supplementation Attenuates Exercise-Induced Inflammation in Postmenopausal Women. Nutr. J. 2014, 13, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sang, S.; Chu, Y.F. Whole Grain Oats, More than Just a Fiber: Role of Unique Phytochemicals. Mol. Nutr. Food Res. 2017, 61, 1600715. [Google Scholar] [CrossRef] [PubMed]
- Kohen, R.; Nyska, A. Oxidation of Biological Systems: Oxidative Stress Phenomena, Antioxidants, Redox Reactions, and Methods for Their Quantification. Toxicol. Pathol. 2002, 30, 620–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, C.L.; Davies, M.J. Detection and Characterisation of Radicals in Biological Materials Using EPR Methodology. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Dikalov, S.; Griendling, K.K.; Harrison, D.G. Measurement of Reactive Oxygen Species in Cardiovascular Studies. Hypertension 2007, 49, 717–727. [Google Scholar] [CrossRef] [Green Version]
- Riebe, D.; Franklin, B.A.; Thompson, P.D.; Garber, C.E.; Whitfield, G.P.; Magal, M.; Pescatello, L.S. Updating ACSM’s Recommendations for Exercise Preparticipation Health Screening. Med. Sci. Sports Exerc. 2015, 47, 2473–2479. [Google Scholar] [CrossRef] [Green Version]
- Frey, I.; Berg, A.; Grathwohl, D.; Keul, J. [Freiburg Questionnaire of Physical Activity—Development, Evaluation and Application]. Soz. Praventivmed. 1999, 44, 55–64. [Google Scholar] [CrossRef]
- Kander, M.C.; Cui, Y.; Liu, Z. Gender Difference in Oxidative Stress: A New Look at the Mechanisms for Cardiovascular Diseases. J. Cell. Mol. Med. 2017, 21, 1024–1032. [Google Scholar] [CrossRef]
- Urbaniak, G.C.; Plous, S. Research Randomizer (Version 4.0). Available online: http://www.randomizer.org/ (accessed on 1 May 2019).
- Ito, S. High-Intensity Interval Training for Health Benefits and Care of Cardiac Diseases—The Key to an Efficient Exercise Protocol. World J. Cardiol. 2019, 11, 171–188. [Google Scholar] [CrossRef]
- Klika, B.; Jordan, C. High-Intensity Circuit Training Using Body Weight: Maximum Results with Minimal Investment. ACSM’s Heal. Fit. J. 2013, 17, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Bird, S.P.; Tarpenning, K.M.; Marino, F.E. Designing Resistance Training Programmes to Enhance Muscular Fitness: A Review of the Acute Programme Variables. Sport. Med. 2005, 35, 841–851. [Google Scholar] [CrossRef] [PubMed]
- American College of Sports Medicine. American College of Sports Medicine Position Stand. Progression Models in Resistance Training for Healthy Adults. Med. Sci. Sports Exerc. 2009, 41, 687–708. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.J.S.; Midgley, A.W.; Thomas, G.; Thurlow, S.; McNaughton, L.R. The Effects of Low- And High-Glycemic Index Meals on Time Trial Performance. Int. J. Sports Physiol. Perform. 2009, 4, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Haferflocken von Kölln. Available online: http://www.koelln.de (accessed on 1 May 2019).
- Schwarzwaldmilch. Available online: https://www.schwarzwaldmilch.de (accessed on 1 May 2019).
- Mariappan, N.; Elks, C.M.; Fink, B.; Francis, J. TNF-Induced Mitochondrial Damage: A Link between Mitochondrial Complex I Activity and Left Ventricular Dysfunction. Free Radic. Biol. Med. 2009, 46, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Mrakic-Sposta, S.; Gussoni, M.; Montorsi, M.; Porcelli, S.; Vezzoli, A. Assessment of a Standardized ROS Production Profile in Humans by Electron Paramagnetic Resonance. Oxid. Med. Cell. Longev. 2012, 2012, 1–10. [Google Scholar] [CrossRef]
- Kyle, U.G.; Bosaeus, I.; De Lorenzo, A.D.; Deurenberg, P.; Elia, M.; Gomez, J.M.; Heitmann, B.L.; Kent-Smith, L.; Melchior, J.C.; Pirlich, M.; et al. Bioelectrical Impedance Analysis—Part II: Utilization in Clinical Practice. Clin. Nutr. 2004, 23, 1430–1453. [Google Scholar] [CrossRef]
- Borg, G.A.V. Psychophysical Bases of Perceived Exertion. Plast. Reconstr. Surg. 1954, 14, 377–381. [Google Scholar] [CrossRef]
- Peterson, D.M. Oat Antioxidants. J. Cereal Sci. 2001, 33, 115–129. [Google Scholar] [CrossRef]
- Khan, I.T.; Bule, M.; Ullah, R.; Nadeem, M.; Asif, S.; Niaz, K. The Antioxidant Components of Milk and Their Role in Processing, Ripening, and Storage: Functional Food. Vet. World 2019, 12, 12–33. [Google Scholar] [CrossRef] [Green Version]
- Peterson, D.M.; Hahn, M.J.; Emmons, C.L. Oat Avenanthramides Exhibit Antioxidant Activities in Vitro. Food Chem. 2002, 79, 473–478. [Google Scholar] [CrossRef]
- Chen, C.Y.O.; Milbury, P.E.; Collins, F.W.; Blumberg, J.B. Avenanthramides Are Bioavailable and Have Antioxidant Activity in Humans after Acute Consumption of an Enriched Mixture from Oats. J. Nutr. 2007, 137, 1375–1382. [Google Scholar] [CrossRef] [Green Version]
- Molteberg, E.L.; Solheim, R.; Dimberg, L.H.; Frølich, W. Variation in Oat Groats Due to Variety, Storage and Heat Treatment. Ii: Sensory Quality. J. Cereal Sci. 1996, 24, 273–282. [Google Scholar] [CrossRef]
- Dimburg, L.H.; Theander, O.; Lingnert, H. Avenanthramides-A Group of Phenolic Antioxidants in Oats. Cereal Chem. 1993, 70, 637–641. [Google Scholar]
- Lee-Manion, A.M.; Price, R.K.; Strain, J.J.; Dimberg, L.H.; Sunnerheim, K.; Welch, R.W. In Vitro Antioxidant Activity and Antigenotoxic Effects of Avenanthramides and Related Compounds. J. Agric. Food Chem. 2009, 57, 10619–10624. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Zhu, Y.; Yerke, A.; Wise, M.L.; Johnson, J.; Chu, Y.; Sang, S. Oat Avenanthramides Induce Heme Oxygenase-1 Expression via Nrf2-Mediated Signaling in HK-2 Cells. Mol. Nutr. Food Res. 2015, 59, 2471–2479. [Google Scholar] [CrossRef] [PubMed]
- Stephens, N.G.; Parsons, A.; Schofield, P.M.; Kelly, F.; Cheeseman, K.; Mitchinson, M.J.; Brown, M.J. Randomised Controlled Trial of Vitamin E in Patients with Coronary Disease: Cambridge Heart Antioxidant Study (CHAOS). Lancet 1996, 347, 781–786. [Google Scholar] [CrossRef]
- Singh, R.; De, S.; Belkheir, A. Avena Sativa (Oat), A Potential Neutraceutical and Therapeutic Agent: An Overview. Crit. Rev. Food Sci. Nutr. 2013, 53, 126–144. [Google Scholar] [CrossRef]
- Ciecierska, A.; Drywień, M.E.; Hamulka, J.; Sadkowski, T. Nutraceutical Functions of Beta-Glucans in Human Nutrition. Rocz. Panstw. Zakl. Hig. 2019, 70, 315–324. [Google Scholar] [CrossRef]
- Grażyna, C.; Hanna, C.; Adam, A.; Magdalena, B.M. Natural Antioxidants in Milk and Dairy Products. Int. J. Dairy Technol. 2017, 70, 1–14. [Google Scholar] [CrossRef]
- Scherr, J.; Wolfarth, B.; Christle, J.W.; Pressler, A.; Wagenpfeil, S.; Halle, M. Associations between Borg’s Rating of Perceived Exertion and Physiological Measures of Exercise Intensity. Eur. J. Appl. Physiol. 2013, 113, 147–155. [Google Scholar] [CrossRef]
- Peter Adams, O. The Impact of Brief High-Intensity Exercise on Blood Glucose Levels. Diabetes Metab. Syndr. Obes. Targets Ther. 2013, 6, 113–122. [Google Scholar] [CrossRef] [Green Version]
- McAnulty, S.R.; McAnulty, L.S.; Nieman, D.C.; Dumke, C.L.; Morrow, J.D.; Utter, A.C.; Henson, D.A.; Proulx, W.R.; George, G.L. Consumption of Blueberry Polyphenols Reduces Exercise-Induced Oxidative Stress Compared to Vitamin C. Nutr. Res. 2004, 24, 209–221. [Google Scholar] [CrossRef]
- Lafay, S.; Jan, C.; Nardon, K.; Lemaire, B.; Ibarra, A.; Roller, M.; Houvenaeghel, M.; Juhel, C.; Cara, L. Grape Extract Improves Antioxidant Status and Physical Performance in Elite Male Athletes. J. Sport. Sci. Med. 2009, 8, 468–480. [Google Scholar]
- Chang, W.H.; Hu, S.P.; Huang, Y.F.; Yeh, T.S.; Liu, J.F. Effect of Purple Sweet Potato Leaves Consumption on Exercise-Induced Oxidative Stress and IL-6 and HSP72 Levels. J. Appl. Physiol. 2010, 109, 1710–1715. [Google Scholar] [CrossRef]
- Dernbach, A.R.; Sherman, W.M.; Simonsen, J.C.; Flowers, K.M.; Lamb, D.R. No Evidence of Oxidant Stress during High-Intensity Rowing Training. J. Appl. Physiol. 1993, 74, 2140–2145. [Google Scholar] [CrossRef]
- Kerksick, C.; Taylor IV, L.; Harvey, A.; Willoughby, D. Gender-Related Differences in Muscle Injury, Oxidative Stress, and Apoptosis. Med. Sci. Sports Exerc. 2008, 40, 1772–1780. [Google Scholar] [CrossRef] [PubMed]
- Strehlow, K.; Rotter, S.; Wassmann, S.; Adam, O.; Grohé, C.; Laufs, K.; Böhm, M.; Nickenig, G. Modulation of Antioxidant Enzyme Expression and Function by Estrogen. Circ. Res. 2003, 93, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Margaritelis, N.V.; Kyparos, A.; Paschalis, V.; Theodorou, A.A.; Panayiotou, G.; Zafeiridis, A.; Dipla, K.; Nikolaidis, M.G.; Vrabas, I.S. Reductive Stress after Exercise: The Issue of Redox Individuality. Redox Biol. 2014, 2, 520–528. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: New York, NY, USA, 2015; ISBN 978-0198717485. [Google Scholar]
- Hargreaves, M.; Hawley, J.A.; Jeukendrup, A. Pre-Exercise Carbohydrate and Fat Ingestion: Effects on Metabolism and Performance. J. Sports Sci. 2004, 22, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M. Pre-Exercise Nutritional Strategies: Effects on Metabolism and Performance. Can. J. Appl. Physiol. 2001, 26, 64–70. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| TitleNutrients | Oat (per 100 g) [35] | Skim Milk (per 100 mL) [36] |
|---|---|---|
| Energy (kcal) | 361 | 48 |
| Carbohydrate (g) | 56 | 5.0 |
| Fat (g) | 6.7 | 1.5 |
| Protein (g) | 14 | 3.5 |
| Unsaturated fatty acids (g) | 5.4 | 0.5 |
| Saturated fatty acids (g) | 1.3 | 1.0 |
| Sugar (g) | 1.2 | 5.0 |
| Fiber (g) | 11 | 0 |
| Variable | Mean ± SD | MIN | MAX |
|---|---|---|---|
| Age (years) | 25.5 ± 5.1 | 21 | 37 |
| Height (cm) | 167.4 ± 6.4 | 156 | 180 |
| Weight (kg) | 62.5 ± 6.1 | 54.2 | 73.8 |
| BMI (kg·m−2) | 22.3 ± 1.9 | 18.7 | 25.5 |
| Body fat (%) | 31.4 ± 3.8 | 21.8 | 39.2 |
| Variable | Time Point | Oatmeal | Control | RMANOVA b (4 × 2) |
|---|---|---|---|---|
| ROS (μM/min) | Pre-meal | 0.70 ± 0.1 | 0.75 ± 0.15 | p < 0.05 |
| Pre-HIIT | 0.69 ± 0.12 | 0.71 ± 0.12 | ||
| Post-HIIT | 0.78 ± 0.13 a | 0.94 ± 0.20 | ||
| 15 post-HIIT | 0.72 ± 0.15 | 0.71 ± 0.11 | ||
| Glucose (mmol/L) | Pre-meal | 5.18 ± 0.26 | 5.19 ± 0.43 | p < 0.05 |
| Pre-HIIT | 5.60 ± 0.65 a | 4.69 ± 0.34 | ||
| Post-HIIT | 4.80 ± 0.48 a | 5.33 ± 0.46 | ||
| 15 post-HIIT | 5.4 3± 0.42 | 5.40 ± 0.49 | ||
| Lactate (mmol/L) | Pre-meal | 1.31 ± 0.48 | 1.26 ± 0.36 | p = 0.579 |
| Pre-HIIT | 2.13 ± 0.94 a | 1.61 ± 0.38 | ||
| Post-HIIT | 5.18 ± 1.72 | 4.74 ± 2.04 | ||
| 15 post-HIIT | 2.96 ± 1.19 | 3.13 ±1.30 |
| Variable | Time Point | Oatmeal | Control | RMANOVA a (4 × 2) |
|---|---|---|---|---|
| Borg | Pre-meal | 6.88 ± 1.41 | 6.06 ± 0.25 | p = 0.987 |
| Pre-HIIT | 6.88 ± 1.26 | 6.31 ± 0.60 | ||
| Post-HIIT | 16.00 ± 1.41 | 15.25 ± 2.32 | ||
| 15 post-HIIT | 8.19 ± 2.10 | 7.25 ± 1.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Z.; Jendricke, P.; Centner, C.; Storck, H.; Gollhofer, A.; König, D. Acute Effects of Oatmeal on Exercise-Induced Reactive Oxygen Species Production Following High-Intensity Interval Training in Women: A Randomized Controlled Trial. Antioxidants 2021, 10, 3. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010003
Zeng Z, Jendricke P, Centner C, Storck H, Gollhofer A, König D. Acute Effects of Oatmeal on Exercise-Induced Reactive Oxygen Species Production Following High-Intensity Interval Training in Women: A Randomized Controlled Trial. Antioxidants. 2021; 10(1):3. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010003
Chicago/Turabian StyleZeng, Zhen, Patrick Jendricke, Christoph Centner, Helen Storck, Albert Gollhofer, and Daniel König. 2021. "Acute Effects of Oatmeal on Exercise-Induced Reactive Oxygen Species Production Following High-Intensity Interval Training in Women: A Randomized Controlled Trial" Antioxidants 10, no. 1: 3. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010003