
Methionine Sulfoxide Reductase B Regulates the Activity of Ascorbate Peroxidase of Banana Fruit


 and
and 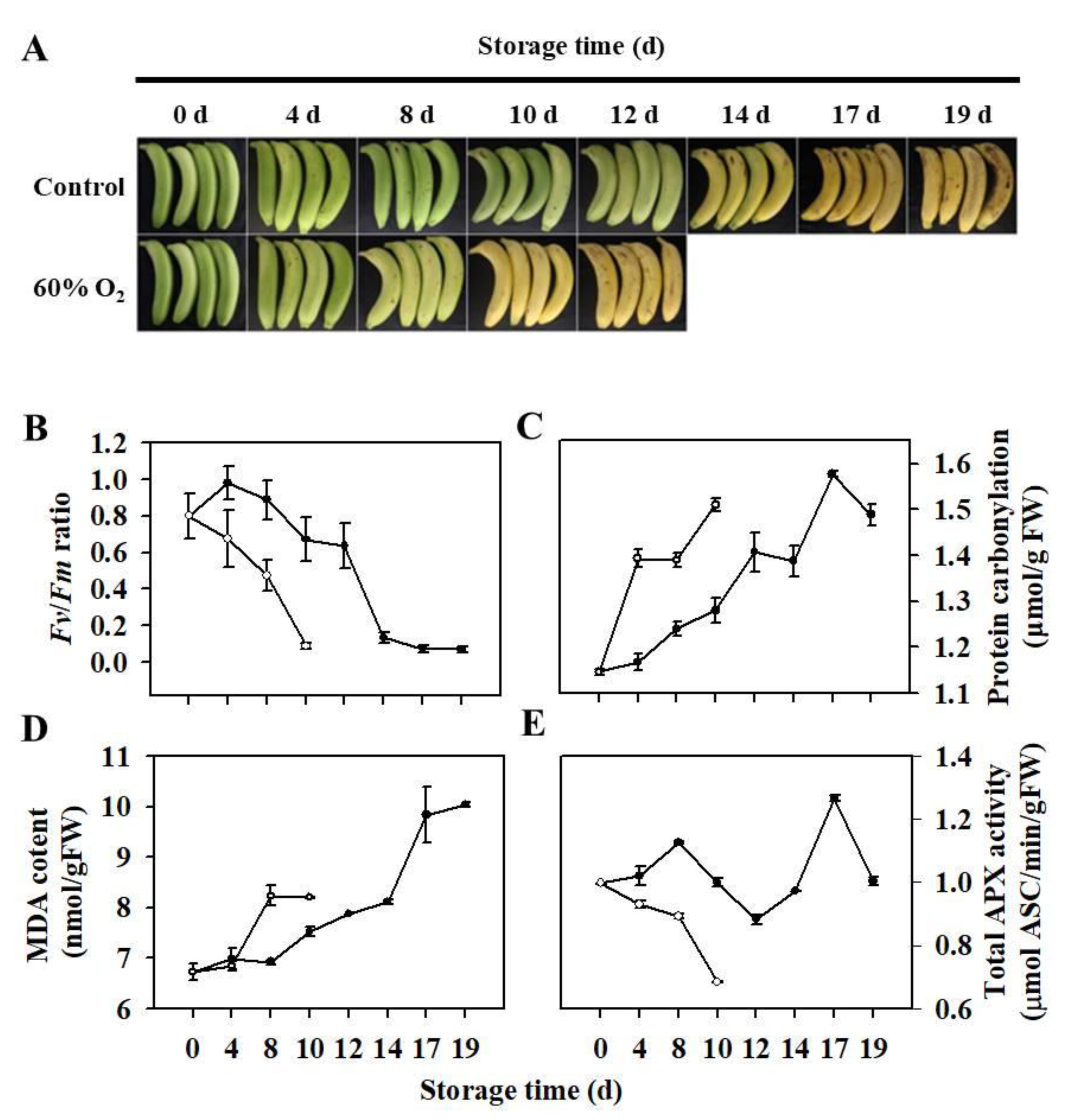{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Measurement of Physiological Parameters
2.3. RNA Extraction, Gene Isolation, and Expression Analyses
2.4. Generation of Transgenic Lines and Dark Treatment
2.5. Subcellular Localization Analysis of MaMsrB2 and MaAPX1
2.6. Site-Directed Mutagenesis of Met36 Residues
2.7. Preparation of Recombinant Proteins
2.8. Yeast Two-Hybrid (Y2H) Assay
2.9. Bimolecular Fluorescence Complementation (BiFC) Assay
2.10. GST Pull-down Assay
2.11. Oxidation and Reduction of MaAPX1
2.12. Enzymatic Activity Measurement
2.13. Spectroscopic Analysis
2.14. Statistical Analysis
3. Results
3.1. Ripening Characteristics and Redox Status of Harvested Banana Fruit under High Oxygen Stress
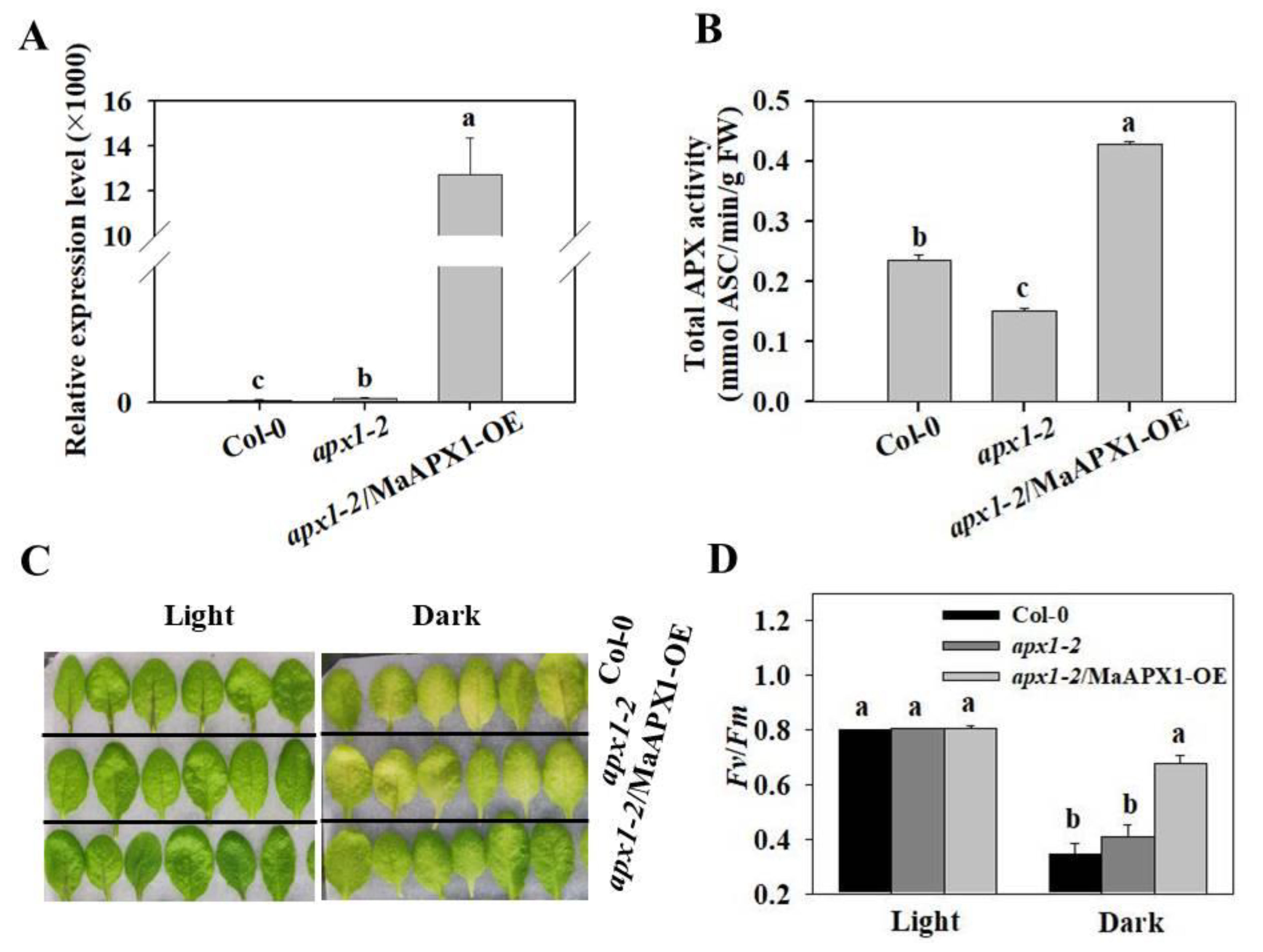
3.2. Overexpression of MaAPX1 in apx1-2 Delays Senescence under Dark Condition
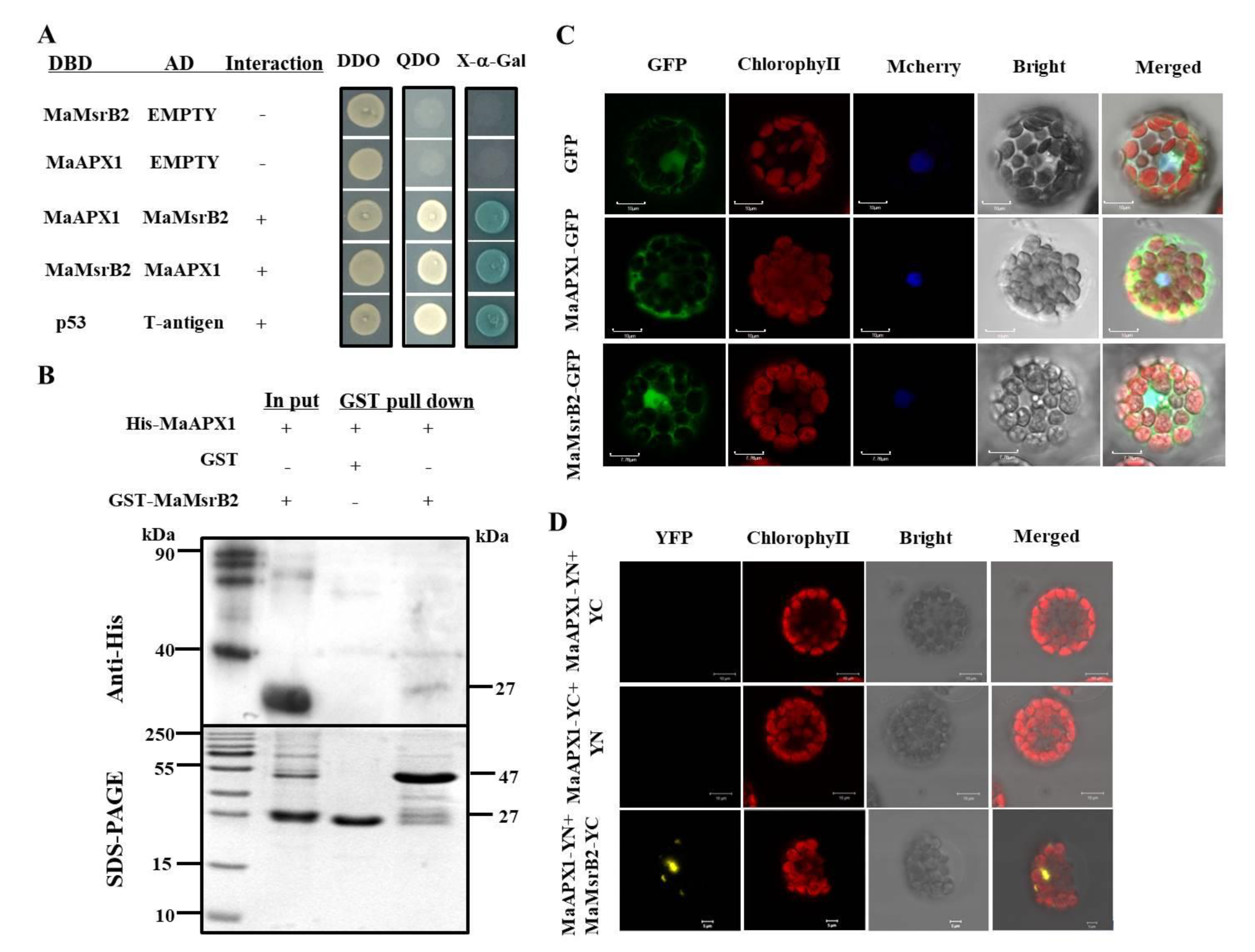
3.3. MaAPX1 Interacts Physically with MaMsrB2
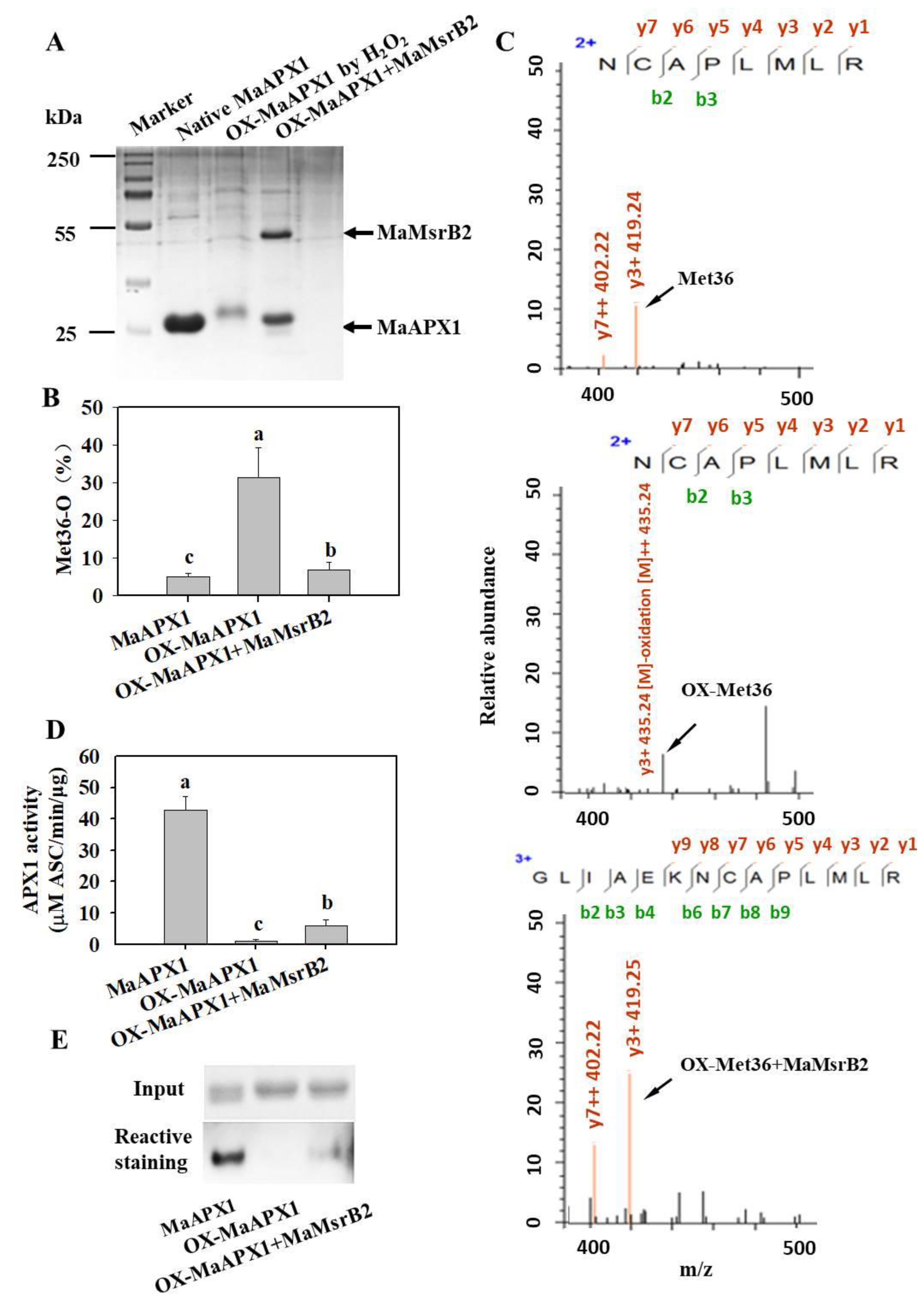
3.4. MaMsrB2 Regulates Redox State and Activity of MaAPX1
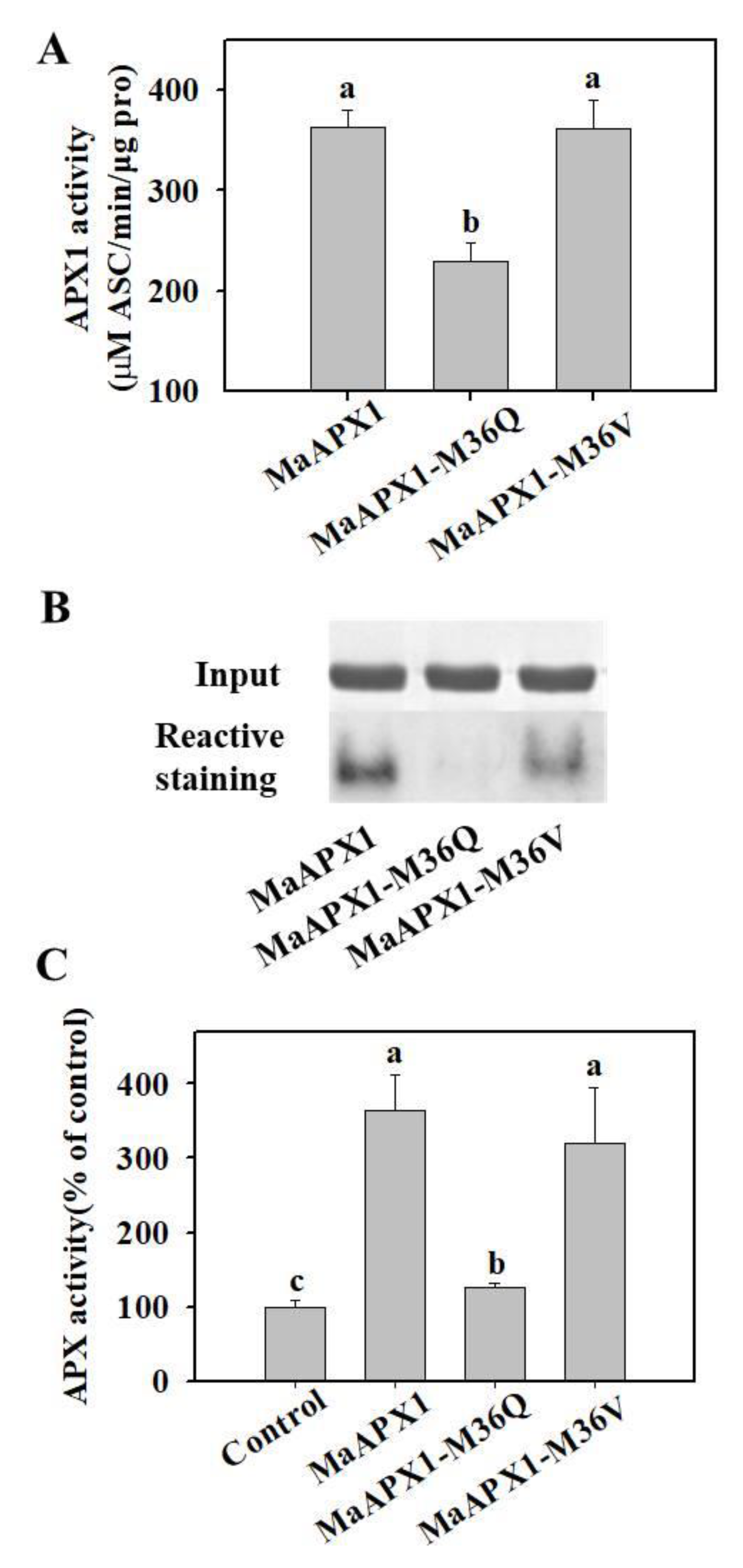
3.5. Mimicked Oxidation of Met in MaAPX1 Decreases the Activity of MaAPX1 In Vitro and In Vivo
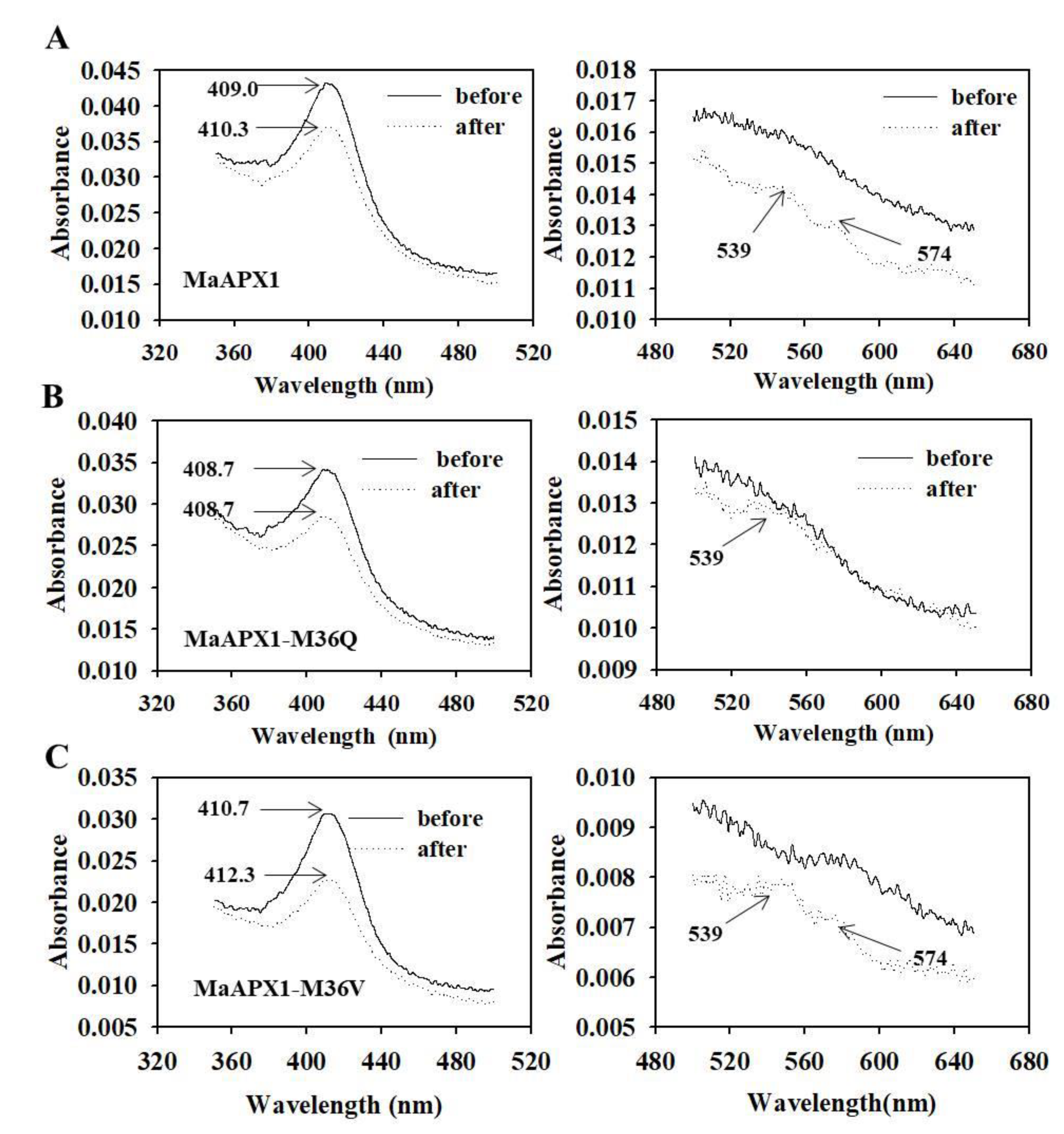
3.6. Visible Spectral Analysis of MaAPX1, MaAPX1-M36Q, and MaAPX1-M36V Reacting with H2O2
4. Discussion
4.1. MaAPX 1 Might Be Involved in Ripening and Senescence in Relation to Oxidative Stress
4.2. MaAPX1 Is a Target of MaMsrB2
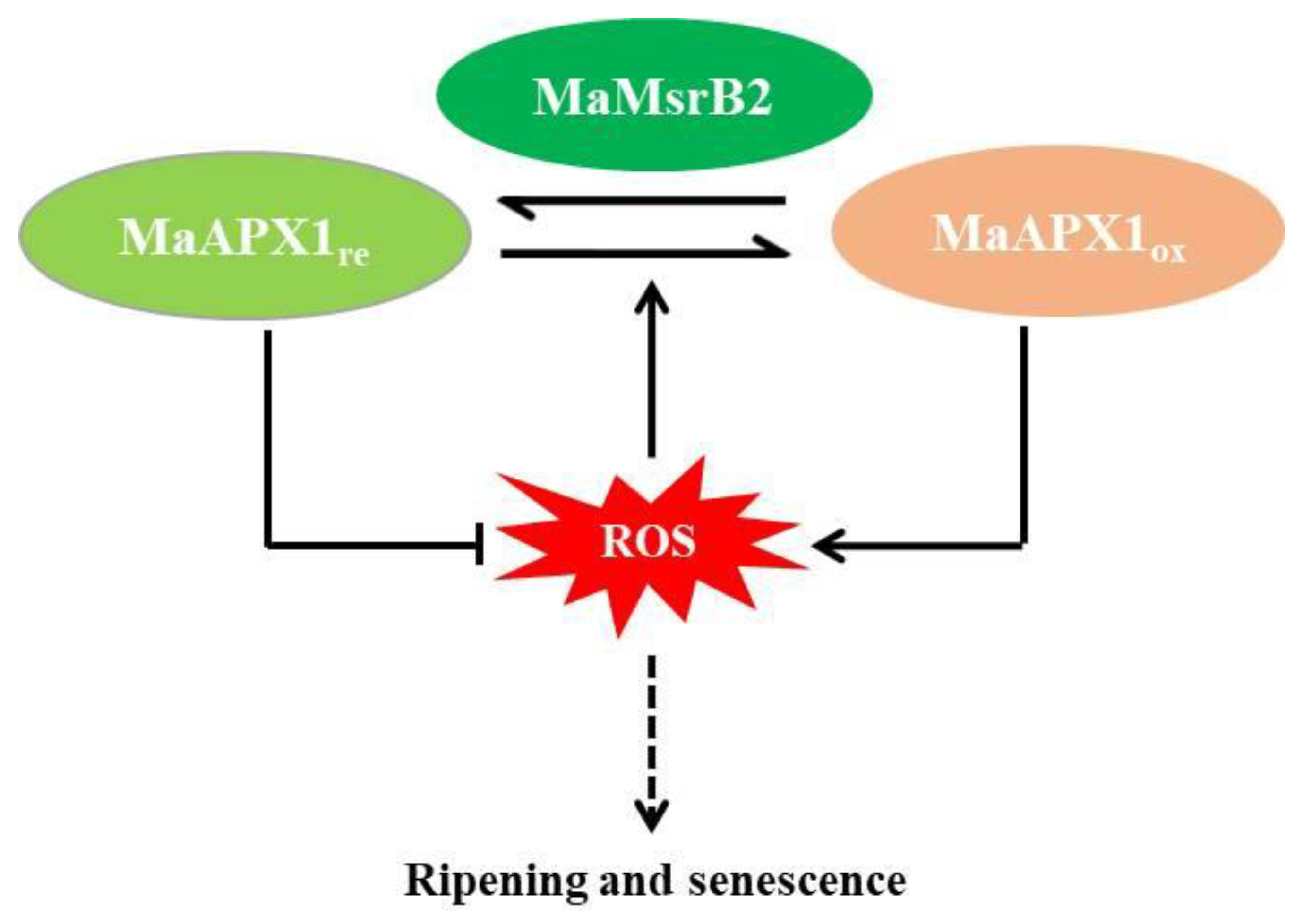
4.3. MaMsrB2 Modifies the Redox Status of MaAPX1 and Affects Its Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2016, 90, 856–867. [Google Scholar] [CrossRef]
- Pitzschke, A.; Hirt, H. Mitogen-activated protein kinases and reactive oxygen species signaling in plants. Plant Physiol. 2006, 141, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [Green Version]
- Rey, P.; Tarrago, L. Physiological roles of plant methionine sulfoxide reductases in redox homeostasis and signaling. Antioxidants 2018, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, R.L.; Moskovitz, J.; Stadtman, E.R. Oxidation of methionine in proteins: Roles in antioxidant defense and cellular regulation. IUBMB Life 2000, 50, 301–307. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Agrawal, M. Reduction in metal toxicity by applying different soil amendments in agricultural field and its consequent effects on characteristics of radish plants (Raphanus sativus L.). J. Agric. Sci Technol. 2013, 15, 1553–1564. [Google Scholar]
- Ferreira, I.C.F.R.; Barros, L.; Abreu, R.M.V. Antioxidants in wild mushrooms. Curr. Med. Chem. 2009, 16, 1543–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, S.B.; Caverzan, A.; Teixeira, F.K.; Lazzarotto, F.; Silveira, J.A.; Ferreira-Silva, S.L.; Abreu-Neto, J.; Margis, R.; Margis-Pinheiro, M. Cytosolic APx knockdown indicates an ambiguous redox responses in rice. Phytochemistry 2010, 71, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Jia, J.; Zhao, X.; Zhang, M.; Huang, X.; Ji, E.; Ni, L.; Jiang, M. The ascorbate peroxidase APX1 is a direct target of a zinc finger transcription factor ZFP36 and a late embryogenesis abundant protein OsLEA5 interacts with ZFP36 to co-regulate OsAPX1 in seed germination in rice. Biochem. Biophys. Res. Commun. 2018, 495, 339–345. [Google Scholar] [CrossRef]
- Ribeiro, C.W.; Korbes, A.P.; Garighan, J.A.; Jardim-Messeder, D.; Carvalho, F.E.; Sousa, R.H.; Caverzan, A.; Teixeira, F.K.; Silveira, J.A.; Margis-Pinheiro, M. Rice peroxisomal ascorbate peroxidase knockdown affects ROS signaling and triggers early leaf senescence. Plant Sci. 2017, 263, 55–65. [Google Scholar] [CrossRef] [PubMed]
- De Pinto, M.C.; Locato, V.; Sgobba, A.; Romero-Puertas, M.D.C.; Gadaleta, C.; Delledonne, M.; De Gara, L. S-Nitrosylation of ascorbate peroxidase is part of programmed cell death signaling in tobacco bright yellow-2 Cells. Plant Physiol. 2013, 163, 1766–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-X.; Feng, K.; Duan, A.-Q.; Li, H.; Yang, Q.-Q.; Xu, Z.-S.; Xiong, A.-S. Isolation, purification and characterization of an ascorbate peroxidase from celery and overexpression of the AgAPX1 gene enhanced ascorbate content and drought tolerance in Arabidopsis. BMC Plant Biol. 2019, 19, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Dunajska-Ordak, K.; Skorupa-Kłaput, M.; Kurnik, K.; Tretyn, A.; Tyburski, J. Cloning and expression analysis of a gene encoding for ascorbate peroxidase and responsive to salt stress in beet (Beta vulgaris). Plant Mol. Biol. Rep. 2014, 32, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Yang, J.; Duan, X.; Jiang, Y.; Zhang, P. Increased expression of native cytosolic Cu/Zn superoxide dismutase and ascorbate peroxidase improves tolerance to oxidative and chilling stresses in cassava (Manihot esculenta Crantz). BMC Plant Biol. 2014, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.-L.; Liu, Q.; Li, Y.-Q.; Zhong, M.; Huang, C.-H.; Jia, D.-F.; Xu, X.-B. Identification and expression profiling analysis of ascorbate peroxidase gene family in Actinidia chinensis (Hongyang). J. Plant Res. 2020, 133, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Kuo, E.Y.; Cai, M.-S.; Lee, T.-M. Ascorbate peroxidase 4 plays a role in the tolerance of Chlamydomonas reinhardtii to photo-oxidative stress. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Wang, X.; Huang, W.; Yang, Z.; Liu, J.; Huang, B. Transcriptional regulation of heat shock proteins and ascorbate peroxidase by CtHsfA2b from African bermudagrass conferring heat tolerance in Arabidopsis. Sci. Rep. 2016, 6, 28021. [Google Scholar] [CrossRef]
- Ugarte, N.; Petropoulos, I.; Friguet, B. Oxidized mitochondrial protein degradation and repair in aging and oxidative stress. Antioxid. Redox Signal. 2010, 13, 539–549. [Google Scholar] [CrossRef]
- Châtelain, E.; Satour, P.; Laugier, E.; Vu, B.L.; Payet, N.; Rey, P.; Montrichard, F. Evidence for participation of the methionine sulfoxide reductase repair system in plant seed longevity. Proc. Natl. Acad. Sci. USA 2013, 110, 3633–3638. [Google Scholar] [CrossRef] [Green Version]
- Laugier, E.; Tarrago, L.; Dos Santos, C.V.; Eymery, F.; Havaux, M.; Rey, P. Arabidopsis thaliana plastidial methionine sulfoxide reductases B, MSRBs, account for most leaf peptide MSR activity and are essential for growth under environmental constraints through a role in the preservation of photosystem antennae. Plant J. 2009, 61, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Li, C.-W.; Koh, K.W.; Chuang, H.-Y.; Chen, Y.-R.; Lin, C.-S.; Chan, M.-T. MSRB7 reverses oxidation of GSTF2/3 to confer tolerance of Arabidopsis thaliana to oxidative stress. J. Exp. Bot. 2014, 65, 5049–5062. [Google Scholar] [CrossRef] [Green Version]
- Stolarska, E.; Bilska, K.; Wojciechowska, N.; Bagniewska-Zadworna, A.; Rey, P.; Kalemba, E.M. Integration of MsrB1 and MsrB2 in the redox network during the development of orthodox and recalcitrant acer seeds. Antioxidants 2020, 9, 1250. [Google Scholar] [CrossRef] [PubMed]
- Khor, H.K.; Fisher, M.T.; Schöneich, C. Potential role of methionine sulfoxide in the inactivation of the chaperone GroEL by hypochlorous acid (HOCl) and peroxynitrite (ONOO–). J. Biol. Chem. 2004, 279, 19486–19493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezraty, B.; Aussel, L.; Barras, F. Methionine sulfoxide reductases in prokaryotes. Biochim. Biophys. Acta Proteins Proteom. 2005, 1703, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Limberis, J.; Martin, R.L.; Xu, R.; Kolbe, K.; Heinemann, S.H.; Hoshi, T.; Cox, B.F.; Gintant, G.A. Functional consequences of methionine oxidation of hERG potassium channels. Biochem. Pharmacol. 2007, 74, 702–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, B.; Cavigiolio, G.; Brot, N.; Oda, M.N.; Heinecke, J.W. Methionine oxidation impairs reverse cholesterol transport by apolipoprotein A-I. Proc. Natl. Acad. Sci. USA 2008, 105, 12224–12229. [Google Scholar] [CrossRef] [Green Version]
- Erickson, J.R.; Joiner, M.-L.A.; Guan, X.; Kutschke, W.; Yang, J.; Oddis, C.V.; Bartlett, R.K.; Lowe, J.S.; O’Donnell, S.E.; Aykin-Burns, N.; et al. A dynamic pathway for calcium-independent activation of CaMKII by methionine oxidation. Cell 2008, 133, 462–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, G.; Lee, K.P.; van der Wijst, J.; De Graaf, M.; van der Kemp, A.; Bindels, R.J.M.; Hoenderop, J.G.J. Methionine sulfoxide reductase B1 (MsrB1) recovers TRPM6 channel activity during oxidative stress. J. Biol. Chem. 2010, 285, 26081–26087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drazic, A.; Miura, H.; Peschek, J.; Le, Y.; Bach, N.C.; Kriehuber, T.; Winter, J. Methionine oxidation activates a transcription factor in response to oxidative stress. Proc. Natl. Acad. Sci. USA 2013, 110, 9493–9498. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.C.; Péterfi, Z.; Hoffmann, F.W.; Moore, R.E.; Kaya, A.; Avanesov, A.; Tarrago, L.; Zhou, Y.; Weerapana, E.; Fomenko, D.E.; et al. MsrB1 and MICALs regulate actin assembly and macrophage function via reversible stereoselective methionine oxidation. Mol. Cell 2013, 51, 397–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, G.; Wu, F.; Li, Z.; Li, T.; Gupta, V.K.; Duan, X.; Jiang, Y. Sulfoxidation Regulation of Musa acuminata calmodulin (MaCaM) influences the functions of MaCaM-binding proteins. Plant Cell Physiol. 2018, 59, 1214–1224. [Google Scholar] [CrossRef]
- Jiang, G.; Xiao, L.; Yan, H.; Zhang, D.; Wu, F.; Liu, X.; Su, X.; Dong, X.; Wang, J.; Duan, X.; et al. Redox regulation of methionine in calmodulin affects the activity levels of senescence-related transcription factors in litchi. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Allu, P.K.; Marada, A.; Boggula, Y.; Karri, K.S.; Krishnamoorthy, T. Methionine sulfoxide reductase 2 reversibly regulates Mge1, a cochaperone of mitochondrial Hsp70, during oxidative stress. Mol. Biol. Cell 2014, 26, 406–419. [Google Scholar] [CrossRef] [Green Version]
- Ding, P.; Fang, L.; Wang, G.; Li, X.; Huang, S.; Gao, Y.; Zhu, J.; Xiao, L.; Tong, J.; Chen, F.; et al. Wheat methionine sulfoxide reductase A4.1 interacts with heme oxygenase 1 to enhance seedling tolerance to salinity or drought stress. Plant Mol. Biol. 2019, 101, 203–220. [Google Scholar] [CrossRef]
- Xu, Y.; Li, F.; Zhao, X.; Tan, C.; Wang, B.; Chen, Y.; Cao, J.; Wu, D.; Yu, H. Methionine sulfoxide reductase A attenuates atherosclerosis via repairing dysfunctional HDL in scavenger receptor class B type I deficient mice. FASEB J. 2020, 34, 3805–3819. [Google Scholar] [CrossRef] [Green Version]
- Bigelow, D.J.; Squier, T.C. Thioredoxin-dependent redox regulation of cellular signaling and stress response through reversible oxidation of methionines. Mol. BioSyst. 2011, 7, 2101–2109. [Google Scholar] [CrossRef] [PubMed]
- Gennaris, A.; Ezraty, B.; Henry, C.; Agrebi, R.; Vergnes, A.; Oheix, E.; Bos, J.; Leverrier, P.; Espinosa, L.; Szewczyk, J.; et al. Repairing oxidized proteins in the bacterial envelope using respiratory chain electrons. Nat. Cell Biol. 2015, 528, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.; Adams, Z.; Moonah, S.; Shi, H.; Maupin-Furlow, J.; Moskovitz, J. The antioxidant enzyme methionine sulfoxide reductase A (MsrA) interacts with Jab1/CSN5 and regulates its function. Antioxidants 2020, 9, 452. [Google Scholar] [CrossRef]
- Lee, H.-J.; Park, J.S.; Yoo, H.J.; Lee, H.M.; Lee, B.C.; Kim, J.H. The selenoprotein MsrB1 Instructs dendritic cells to induce t-helper 1 immune responses. Antioxidants 2020, 9, 1021. [Google Scholar] [CrossRef]
- Tossounian, M.-A.; Van Molle, I.; Wahni, K.; Jacques, S.; Gevaert, K.; Van Breusegem, F.; Vertommen, D.; Young, D.; Rosado, L.A.; Messens, J. Disulfide bond formation protects Arabidopsis thaliana glutathione transferase tau 23 from oxidative damage. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 775–789. [Google Scholar] [CrossRef]
- Qin, G.; Meng, X.; Wang, Q.; Tian, S. Oxidative damage of mitochondrial proteins contributes to fruit senescence: A redox proteomics analysis. J. Proteome Res. 2009, 8, 2449–2462. [Google Scholar] [CrossRef]
- Yan, H.; Jiang, G.; Wu, F.; Li, Z.; Xiao, L.; Jiang, Y.; Duan, X. Sulfoxidation regulation of MaNAC42 transcription factor influences its functions in relation to stress-induced fruit ripening in banana. J. Exp. Bot. 2020, 10, 682–699. [Google Scholar] [CrossRef]
- Ueda, H.; Kusaba, M. Strigolactone regulates leaf senescence in concert with ethylene in Arabidopsis. Plant Physiol. 2015, 169, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Wan, C.Y.; Wilkins, T.A. Isolation of multiple cDNAs encoding the vacuolar H+-ATPase subunit B from developing cotton (Cossypium hirsutum L.) ovules. Plant Physiol. 1994, 106, 393–394. [Google Scholar] [CrossRef]
- Zhang, X.; Henriques, R.; Lin, S.-S.; Niu, Q.-W.; Chua, N.-H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.-D.; Cho, Y.-H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, G.; Zeng, J.; Li, Z.; Song, Y.; Yan, H.; He, J.; Jiang, Y.; Duan, X. Redox regulation of the NOR Transcription factor is involved in the regulation of fruit ripening in tomato. Plant Physiol. 2020, 183, 671–685. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Mu, J.; Chen, L.; Feng, J.; Huanjie, Y.; Lichao, C.; Zhou, J.-M.; Zuo, J. S-nitrosylation positively regulates ascorbate peroxidase activity during plant stress responses. Plant Physiol. 2015, 167, 1604–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R.; Zilinskas, B.A. Purification and characterization of pea cytosolic ascorbate peroxidase. Plant Physiol. 1991, 97, 926–968. [Google Scholar] [CrossRef]
- Hugo, M.; Martínez, A.; Trujillo, M.; Estrad, D.; Mastrogiovanni, M.; Linares, E.; Augusto, O.; Issoglio, F.; Zeida, A.; Estrín, D.A.; et al. Kinetics, subcellular localization, and contribution to parasite virulence of a Trypanosoma cruzi hybrid type A heme peroxidase (TcAPx-CcP). Proc. Natl. Acad. Sci. USA 2017, 114, E1326–E1335. [Google Scholar] [CrossRef] [Green Version]
- Davletova, S.; Rizhsky, L.; Liang, H.; Shengqiang, Z.; Oliver, D.J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen gene network of Arabidopsis. Plant Cell 2004, 17, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasion, V.S.; Polanco, J.A.; Meharenna, Y.T.; Li, H.; Poulos, T.L. Crystal structure of leishmania major peroxidase and characterization of the compound I tryptophan radical. J. Biol. Chem. 2011, 286, 24608–24615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, R.K.; Dolai, S.; Pal, S.; Adak, S. Role of tryptophan-208 residue in cytochrome c oxidation by ascorbate peroxidase from Leishmania major-kinetic studies on Trp208Phe mutant and wild type enzyme. Biochim. Biophys. Acta Proteins Proteom. 2008, 1784, 863–871. [Google Scholar] [CrossRef]
- Artés, F.; Allende, A. Processing lines and alternative preservation techniques to prolong shelf-life of minimally fresh processed leafy vegetables. Eur. J. Hortic. Sci. 2005, 70, 231–245. [Google Scholar]
- Jiang, Y.; Joyce, D.C. Softening response of 1-methylcyclopropene-treated banana fruit to high oxygen atmospheres. Plant Growth Regul. 2003, 41, 225–229. [Google Scholar] [CrossRef]
- Odriozola-Serrano, I.; Oliu, G.O.; Soliva-Fortuny, R.; Martín-Belloso, O. Effect of high-oxygen atmospheres on the antioxidant potential of fresh-cut tomatoes. J. Agric. Food Chem. 2009, 57, 6603–6610. [Google Scholar] [CrossRef]
- Duan, X.; Liu, T.; Zhang, D.; Su, X.; Lin, H.; Jiang, Y. Effect of pure oxygen atmosphere on antioxidant enzyme and antioxidant activity of harvested litchi fruit during storage. Food Res. Int. 2011, 44, 1905–1911. [Google Scholar] [CrossRef]
- Liu, X.; Wang, T.; Lu, Y.; Yang, Q.; Li, Y.; Deng, X.; Liu, Y.; Du, X.; Qiao, L.; Zheng, J. Effect of high oxygen pretreatment of whole tuber on anti-browning of fresh-cut potato slices during storage. Food Chem. 2019, 301, 125287. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Li, Y.; Liu, G.; Zhang, Z.; Zhou, H.; Yang, H. High oxygen treatments enhance the contents of phenolic compound and ganoderic acid, and the antioxidant and DNA damage protective activities of Ganoderma lingzhi fruiting body. Front. Microbiol. 2019, 10, 2363. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, X. Changes in color, antioxidant, and free radical scavenging enzyme activity of mushrooms under high oxygen modified atmospheres. Postharvest Biol. Technol. 2012, 69, 1–6. [Google Scholar] [CrossRef]
- Van de Velde, F.; Méndez-Galarraga, M.P.; Pirovani, M.E. Effect of enriched O2 and CO2 atmospheres on the overall quality and the bioactive potential of fresh blackberries. Postharvest Biol. Technol. 2020, 164, 111166. [Google Scholar] [CrossRef]
- Li, T.; Wu, Q.; Zhou, Y.; Yun, Z.; Duan, X.; Jiang, Y. L-Cysteine hydrochloride delays senescence of harvested longan fruit in relation to modification of redox status. Postharvest Biol. Technol. 2018, 143, 35–42. [Google Scholar] [CrossRef]
- Sun, W.-H.; Liu, X.-Y.; Wang, Y.; Hua, Q.; Song, X.-M.; Gu, Z.; Pu, D.-Z. Effect of water stress on yield and nutrition quality of tomato plant overexpressing StAPX. Biol. Plant. 2013, 58, 99–104. [Google Scholar] [CrossRef]
- Shafi, A.; Pal, A.K.; Sharma, V.; Kalia, S.; Kumar, S.; Ahuja, P.S.; Singh, A.K. Transgenic potato plants overexpressing SOD and APX exhibit enhanced lignification and starch biosynthesis with improved salt stress tolerance. Plant Mol. Biol. Rep. 2017, 35, 504–518. [Google Scholar] [CrossRef]
- Kim, M.D.; Kim, Y.-H.; Kwon, S.-Y.; Yun, D.-J.; Kwak, S.-S.; Lee, H.-S. Enhanced tolerance to methyl viologen-induced oxidative stress and high temperature in transgenic potato plants overexpressing the CuZnSOD, APX and NDPK2 genes. Physiol. Plant 2010, 140, 153–162. [Google Scholar] [CrossRef]
- Wang, Y.; Wisniewski, M.; Meilan, R.; Cui, M.; Webb, R.; Fuchigami, L. Overexpression of cytosolic ascorbate peroxidase in tomato confers tolerance to chilling and salt stress. J. Am. Soc. Hortic. Sci. 2005, 130, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.; Suzuki, N.; Rizhsky, L.; Hegie, A.; Koussevitzky, S.; Mittler, R. Double mutants deficient in cytosolic and thylakoid ascorbate peroxidase reveal a complex mode of interaction between reactive oxygen species, plant development, and response to abiotic stresses. Plant Physiol. 2007, 144, 1777–1785. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic stress tolerance in plants: Myriad roles of ascorbate peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarrago, L.; Laugier, E.; Rey, P. Protein-repairing methionine sulfoxide reductases in photosynthetic organisms: Gene Organization, reduction mechanisms, and physiological roles. Mol. Plant 2009, 2, 202–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghesquière, B.; Gevaert, K. Proteomics methods to study methionine oxidation. Mass Spectrom. Rev. 2013, 33, 147–156. [Google Scholar] [CrossRef]
- Ghesquière, B.; Jonckheere, V.; Colaert, N.; Van Durme, J.; Timmerman, E.; Goethals, M.; Schymkowitz, J.; Rousseau, F.; Vandekerckhove, J.; Gevaert, K. Redox proteomics of protein-bound methionine oxidation. Mol. Cell. Proteom. 2011, 10, M110-006866. [Google Scholar] [CrossRef] [Green Version]
- Tarrago, L.; Kieffer-Jaquinod, S.; Lamant, T.; Marcellin, M.; Garin, J.; Rouhier, N.; Rey, P. Affinity chromatography: A valuable strategy to isolate substrates of methionine sulfoxide reductases? Antioxid. Redox Signal. 2012, 16, 79–84. [Google Scholar] [CrossRef]
- Oien, D.B.; Moskovitz, J. Genetic regulation of longevity and age-associated diseases through the methionine sulfoxide reductase system. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1756–1762. [Google Scholar] [CrossRef]
- Valverde, H.; Cantón, F.R.; Aledo, J.C. MetOSite: An integrated resource for the study of methionine residues sulfoxidation. Bioinformatics 2019, 35, 4849–4850. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, S.L.; Petropoulos, I.; Friguet, B. The oxidized protein repair enzymes methionine sulfoxide reductases and their roles in protecting against oxidative stress, in ageing and in regulating protein function. Antioxidants 2018, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, D.; Motohashi, K.; Kasama, T.; Hara, Y.; Hisabori, T. Target proteins of the cytosolic thioredoxins in Arabidopsis thaliana. Plant Cell. Physiol. 2004, 45, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Gelhaye, E.; Navrot, N.; Macdonald, I.K.; Rouhier, N.; Raven, E.L.; Jacquot, J.-P. Ascorbate peroxidase–thioredoxin interaction. Photosynth. Res. 2006, 89, 193–200. [Google Scholar] [CrossRef]
- Bai, X.; Yang, L.; Tian, M.; Chen, J.; Shi, J.; Yang, Y.; Hu, X. Nitric oxide enhances desiccation tolerance of recalcitrant Antiaris toxicaria seeds via protein S-nitrosylation and carbonylation. PLoS ONE 2011, 6, e20714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjum, N.A.; Sharma, P.; Gill, S.S.; Hasanuzzaman, M.; Khan, E.A.; Kachhap, K.; Mohamed, A.A.; Thangavel, P.; Devi, G.D.; Vasudhevan, P.; et al. Catalase and ascorbate peroxidase—representative H2O2-detoxifying heme enzymes in plants. Environ. Sci. Pollut. Res. 2016, 23, 19002–19029. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, L.; Jiang, G.; Yan, H.; Lai, H.; Su, X.; Jiang, Y.; Duan, X. Methionine Sulfoxide Reductase B Regulates the Activity of Ascorbate Peroxidase of Banana Fruit. Antioxidants 2021, 10, 310. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10020310
Xiao L, Jiang G, Yan H, Lai H, Su X, Jiang Y, Duan X. Methionine Sulfoxide Reductase B Regulates the Activity of Ascorbate Peroxidase of Banana Fruit. Antioxidants. 2021; 10(2):310. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10020310
Chicago/Turabian StyleXiao, Lu, Guoxiang Jiang, Huiling Yan, Hongmei Lai, Xinguo Su, Yueming Jiang, and Xuewu Duan. 2021. "Methionine Sulfoxide Reductase B Regulates the Activity of Ascorbate Peroxidase of Banana Fruit" Antioxidants 10, no. 2: 310. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10020310