
Effects of Natural Antioxidants on Phospholipid and Ceramide Profiles of 3D-Cultured Skin Fibroblasts Exposed to UVA or UVB Radiation



Abstract
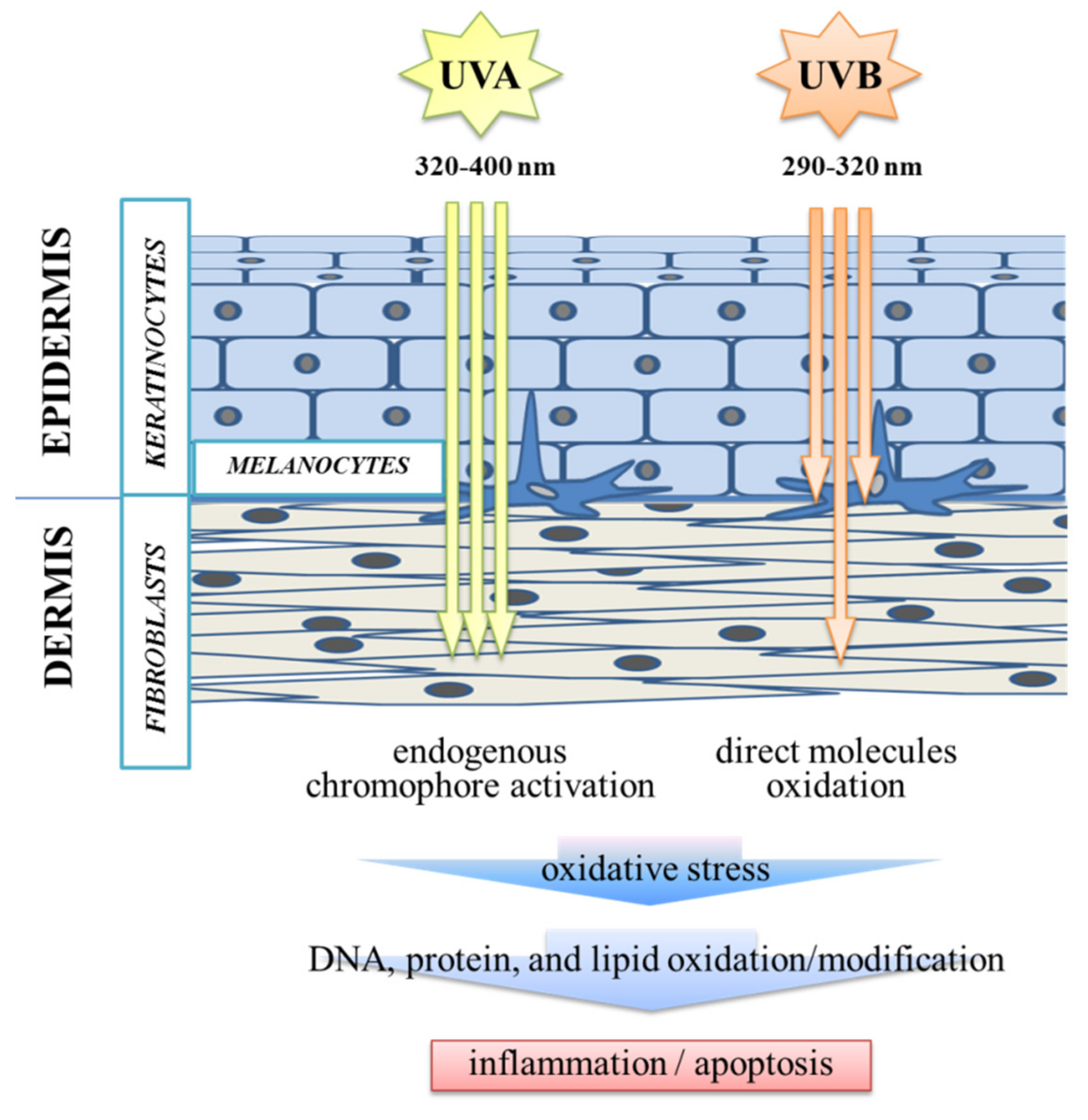
:1. Introduction
2. Materials and Methods
2.1. Reagents/Chemicals
2.2. Cell Culture
2.3. Lipidomic Analysis
2.3.1. Extraction of Lipids and Total Phospholipid Quantification
2.3.2. Phospholipid Separation and Quantification by UPLC-ESI-MS
2.3.3. RPLC-ESI-MS Analysis
2.3.4. Data Processing
2.3.5. Statistical Analysis
3. Results
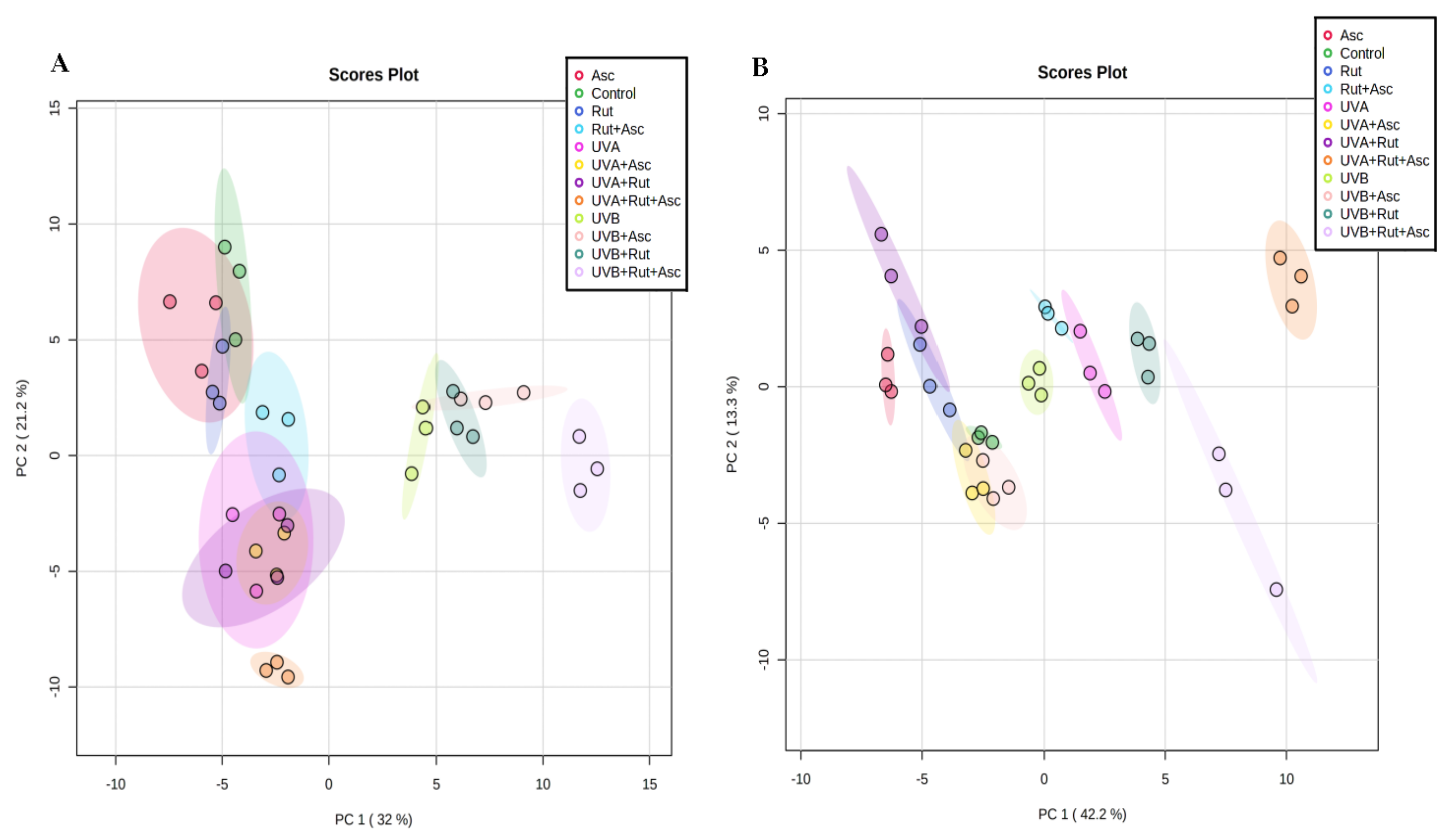
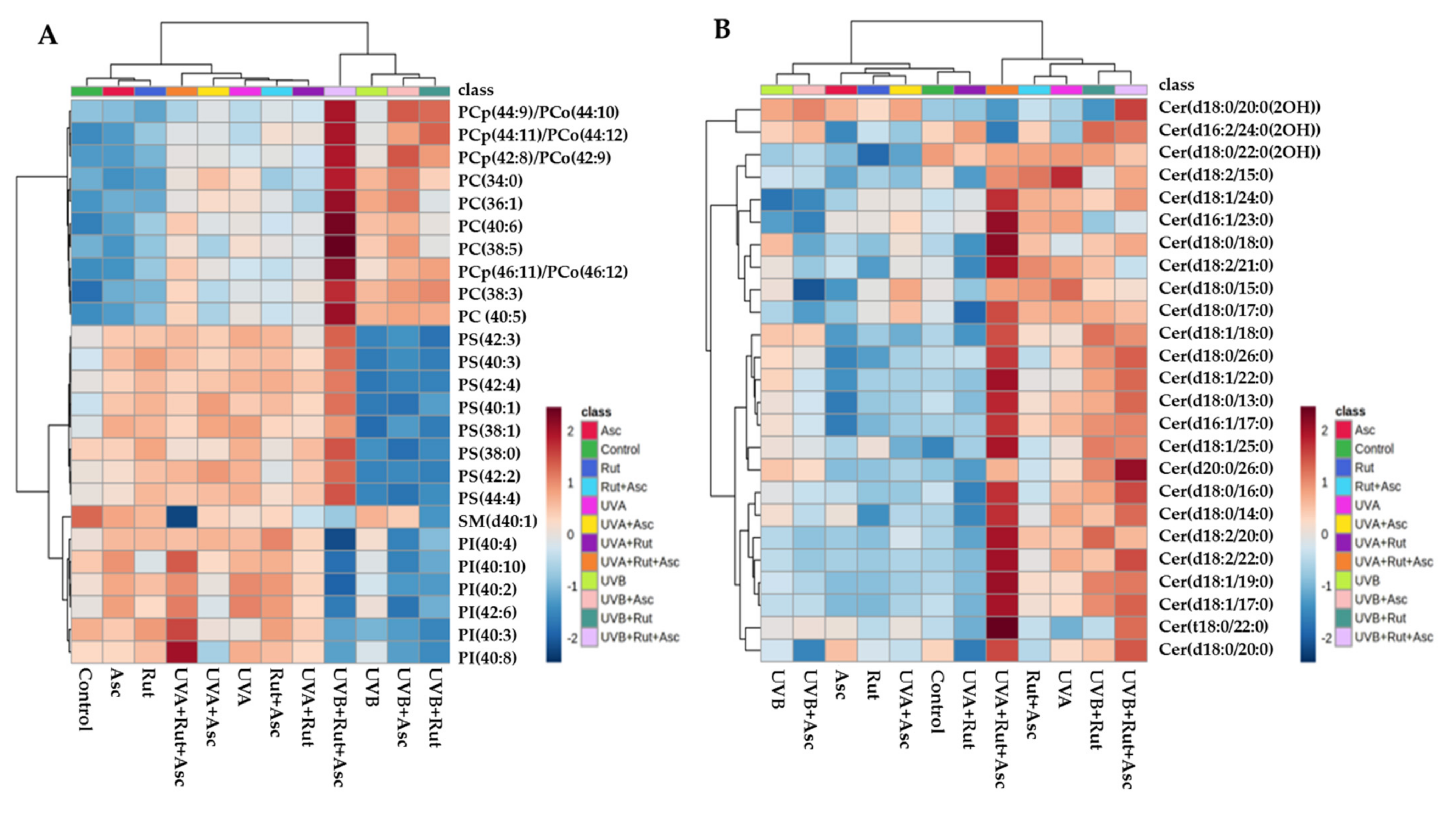
3.1. Comparison of Phospholipid and Ceramide Profiles of Control Fibroblasts vs. Those Treated with Rutin or Ascorbic Acid Separately, as well as with Both Compounds together (Control vs. Rut vs. Asc vs. Rut+Asc)
3.2. Comparison of the Phospholipid and Ceramide Profiles of Fibroblasts Exposed to UVA and then either Not Treated, Treated Separately with Rutin or Ascorbic Acid, or Treated with Both Compounds Together (UVA vs. UVA+Rut/UVA+Asc/UVA+Rut+Asc)
3.3. Comparison of the Phospholipid and Ceramide Profile of Fibroblasts Exposed to UVB and then either Not Treated, Treated Separately with Rutin or Ascorbic Acid, or Treated with Both Compounds Together (UVB vs. UVB+Rut vs. UVB+Asc vs. UVB+Rut+Asc)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Archer, C.B. Functions of the Skin. In Rook’s Textbook of Dermatology; Wiley-Blackwell: Oxford, UK, 2010; Volume 1, pp. 1–11. [Google Scholar]
- Spiekstra, S.W.; Breetveld, M.; Rustemeyer, T.; Scheper, R.J.; Gibbs, S. Wound-healing factors secreted by epidermal keratinocytes and dermal fibroblasts in skin substitutes. Wound Repair Regen. 2007, 15, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Russo, B.; Brembilla, N.C.; Chizzolini, C. Interplay between keratinocytes and fibroblasts: A systematic review providing a new angle for understanding skin fibrotic disorders. Front. Immunol. 2020, 11, 648. [Google Scholar] [CrossRef] [PubMed]
- Breitkreutz, D.; Mirancea, N.; Nischt, R. Basement membranes in skin: Unique matrix structures with diverse functions? Histochem. Cell Biol. 2009, 132, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gegotek, A.; Biernacki, M.; Ambrozewicz, E.; Surazyński, A.; Wroński, A.; Skrzydlewska, E.E.; Gęgotek, A.; Biernacki, M.; Ambrożewicz, E.; Surażyński, A.; et al. The cross-talk between electrophiles, antioxidant defence and the endocannabinoid system in fibroblasts and keratinocytes after UVA and UVB irradiation. J. Dermatol. Sci. 2016, 81, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Kaisers, W.; Boukamp, P.; Stark, H.-J.; Schwender, H.; Tigges, J.; Krutmann, J.; Schaal, H. Age, gender and UV-exposition related effects on gene expression in in vivo aged short term cultivated human dermal fibroblasts. PLoS ONE 2017, 12, e0175657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gruijl, F.R. Photocarcinogenesis: UVA vs. UVB Radiation. Skin Pharmacol. Physiol. 2002, 15, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Gonzaga, E.R. Role of UV light in photodamage, skin aging, and skin cancer: Importance of photoprotection. Am. J. Clin. Dermatol. 2009, 10, 19–24. [Google Scholar] [CrossRef]
- Röck, K.; Meusch, M.; Fuchs, N.; Tigges, J.; Zipper, P.; Fritsche, E.; Krutmann, J.; Homey, B.; Reifenberger, J.; Fischer, J.W. Estradiol protects dermal hyaluronan/versican matrix during photoaging by release of epidermal growth factor from keratinocytes. J. Biol. Chem. 2012, 287, 20056–20069. [Google Scholar] [CrossRef] [Green Version]
- Leuti, A.; Fazio, D.; Fava, M.; Piccoli, A.; Oddi, S.; Maccarrone, M. Bioactive lipids, inflammation and chronic diseases. Adv. Drug Deliv. Rev. 2020, 159, 133–169. [Google Scholar] [CrossRef]
- Kendall, A.C.; Pilkington, S.M.; Massey, K.A.; Sassano, G.; Rhodes, L.E.; Nicolaou, A. Distribution of bioactive lipid mediators in human skin. J. Investig. Dermatol. 2015, 135, 1510–1520. [Google Scholar] [CrossRef] [Green Version]
- Berker, K.I.; Demirata, B.; Apak, R. Determination of Total Antioxidant Capacity of Lipophilic and Hydrophilic Antioxidants In the Same Solution by Using Ferric-Ferricyanide Assay. Food Anal. Methods 2012, 5, 1150–1158. [Google Scholar] [CrossRef]
- Gęgotek, A.; Bielawska, K.; Biernacki, M.; Zaręba, I.; Surażyński, A.; Skrzydlewska, E. Comparison of protective effect of ascorbic acid on redox and endocannabinoid systems interactions in in vitro cultured human skin fibroblasts exposed to UV radiation and hydrogen peroxide. Arch. Dermatol. Res. 2017, 309, 285–303. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Domingues, P.; Skrzydlewska, E. Proteins involved in the antioxidant and inflammatory response in rutin-treated human skin fibroblasts exposed to UVA or UVB irradiation. J. Dermatol. Sci. 2018, 90, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Ragheb, S.R.; El Wakeel, L.M.; Nasr, M.S.; Sabri, N.A. Impact of Rutin and Vitamin C combination on oxidative stress and glycemic control in patients with type 2 diabetes. Clin. Nutr. ESPEN 2020, 35, 128–135. [Google Scholar] [CrossRef]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Walingo, K.M. Role of vitamin C (ascorbic acid) on human health—a review. Afr. J. Food Agric. Nutr. Dev. 2005, 5, 1–13. [Google Scholar]
- Yang, J.; Guo, J.; Yuan, J. In vitro antioxidant properties of rutin. LWT Food Sci. Technol. 2008, 41, 1060–1066. [Google Scholar] [CrossRef]
- Gęgotek, A.; Ambrożewicz, E.; Jastrząb, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Rutin and ascorbic acid cooperation in antioxidant and antiapoptotic effect on human skin keratinocytes and fibroblasts exposed to UVA and UVB radiation. Arch. Dermatol. Res. 2019, 311, 203–219. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Synergistic cytoprotective effects of rutin and ascorbic acid on the proteomic profile of 3D-cultured keratinocytes exposed to UVA or UVB radiation. Nutrients 2019, 11, 2672. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Cytoprotective Effect of Ascorbic Acid and Rutin against Oxidative Changes in the Proteome of Skin Fibroblasts Cultured in a Three-Dimensional System. Nutrients 2020, 12, 1074. [Google Scholar] [CrossRef] [Green Version]
- Gȩgotek, A.; Domingues, P.; Skrzydlewska, E. Natural Exogenous Antioxidant Defense against Changes in Human Skin Fibroblast Proteome Disturbed by UVA Radiation. Oxid. Med. Cell. Longev. 2020, 2020, 3216415. [Google Scholar] [CrossRef] [PubMed]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, Ł.; Lamperska, K. 2D and 3D cell cultures—a comparison of different types of cancer cell cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Atalay, S.; Domingues, P.; Skrzydlewska, E. The Differences in the Proteome Profile of Cannabidiol-Treated Skin Fibroblasts following UVA or UVB Irradiation in 2D and 3D Cell Cultures. Cells 2019, 8, 995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, C.; Rubio, L.; Touriño, S.; Martí, M.; Barba, C.; Fernández-Campos, F.; Coderch, L.; Luís Parra, J. Antioxidative effects and percutaneous absorption of five polyphenols. Free Radic. Biol. Med. 2014, 75, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Rozman, B.; Gasperlin, M.; Tinois-Tessoneaud, E.; Pirot, F.; Falson, F. Simultaneous absorption of vitamins C and E from topical microemulsions using reconstructed human epidermis as a skin model. Eur. J. Pharm. Biopharm. 2009, 72, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Wei, P.; Liu, W. Combined antioxidant effects of rutin and vitamin C in Triton X-100 micelles. J. Pharm. Biomed. Anal. 2007, 43, 1580–1586. [Google Scholar] [CrossRef] [PubMed]
- Godugu, C.; Singh, M. AlgimatrixTM-based 3d cell culture system as an in vitro tumor model: An important tool in cancer research. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2016; Volume 1379, pp. 117–128. [Google Scholar]
- Gęgotek, A.; Rybałtowska-Kawałko, P.; Skrzydlewska, E. Rutin as a Mediator of Lipid Metabolism and Cellular Signaling Pathways Interactions in Fibroblasts Altered by UVA and UVB Radiation. Oxid. Med. Cell. Longev. 2017, 2017, 4721352. [Google Scholar] [CrossRef]
- BLIGH, E.G.; DYER, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, E.M.; Lewis, D.H. Spectrophotometric determination of phosphate esters in the presence and absence of orthophosphate. Anal. Biochem. 1970, 36, 159–167. [Google Scholar] [CrossRef]
- Łuczaj, W.; Dobrzyńska, I.; Wroński, A.; Domingues, M.R.; Domingues, P.; Skrzydlewska, E. Cannabidiol-Mediated Changes to the Phospholipid Profile of UVB-Irradiated Keratinocytes from Psoriatic Patients. Int. J. Mol. Sci. 2020, 21, 6592. [Google Scholar] [CrossRef]
- Łuczaj, W.; Wroński, A.; Domingues, P.; Domingues, M.R.; Skrzydlewska, E. Lipidomic analysis reveals specific differences between fibroblast and keratinocyte ceramide profile of patients with psoriasis vulgaris. Molecules 2020, 25, 630. [Google Scholar] [CrossRef] [Green Version]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Marionnet, C.; Pierrard, C.; Lejeune, F.; Sok, J.; Thomas, M.; Bernerd, F. Different oxidative stress response in keratinocytes and fibroblasts of reconstructed skin exposed to non extreme daily-ultraviolet radiation. PLoS ONE 2010, 5, e12059. [Google Scholar] [CrossRef] [Green Version]
- Fagot, D.; Asselineau, D.; Bernerd, F. Direct role of human dermal fibroblasts and indirect participation of epidermal keratinocytes in MMP-1 production after UV-B irradiation. Arch. Dermatol. Res. 2002, 293, 576–583. [Google Scholar] [CrossRef]
- Jobe, N.P.; Živicová, V.; Mifková, A.; Rösel, D.; Dvořánková, B.; Kodet, O.; Strnad, H.; Kolář, M.; Šedo, A.; Smetana, K.; et al. Fibroblasts potentiate melanoma cells in vitro invasiveness induced by UV-irradiated keratinocytes. Histochem. Cell Biol. 2018, 149, 503–516. [Google Scholar] [CrossRef]
- Brenner, M.; Degitz, K.; Besch, R.; Berking, C. Differential expression of melanoma-associated growth factors in keratinocytes and fibroblasts by ultraviolet A and ultraviolet B radiation. Br. J. Dermatol. 2005, 153, 733–739. [Google Scholar] [CrossRef]
- Pines, A.; Backendorf, C.; Alekseev, S.; Jansen, J.G.; de Gruijl, F.R.; Vrieling, H.; Mullenders, L.H.F. Differential activity of UV-DDB in mouse keratinocytes and fibroblasts: Impact on DNA repair and UV-induced skin cancer. DNA Repair 2009, 8, 153–161. [Google Scholar] [CrossRef]
- Fernandez, T.L.; Van Lonkhuyzen, D.R.; Dawson, R.A.; Kimlin, M.G.; Upton, Z. In vitro investigations on the effect of dermal fibroblasts on keratinocyte responses to ultraviolet B radiation. Photochem. Photobiol. 2014, 90, 1332–1339. [Google Scholar] [CrossRef]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [Green Version]
- Dalmau, N.; Andrieu-Abadie, N.; Tauler, R.; Bedia, C. Phenotypic and lipidomic characterization of primary human epidermal keratinocytes exposed to simulated solar UV radiation. J. Dermatol. Sci. 2018, 92, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Reich, A.; Schwudke, D.; Meurer, M.; Lehmann, B.; Shevchenko, A. Lipidome of narrow-band ultraviolet B irradiated keratinocytes shows apoptotic hallmarks. Exp. Dermatol. 2009, 19, e103–e110. [Google Scholar] [CrossRef]
- Eckes, B.; Nischt, R.; Krieg, T. Cell-matrix interactions in dermal repair and scarring. Fibrogenes. Tissue Repair 2010, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Bielawska, K.; Biernacki, M.; Dobrzyńska, I.; Skrzydlewska, E. Time-dependent effect of rutin on skin fibroblasts membrane disruption following UV radiation. Redox Biol. 2017, 12, 733–744. [Google Scholar] [CrossRef]
- Farrell, A.M.; Uchida, Y.; Nagiec, M.M.; Harris, I.R.; Dickson, R.C.; Elias, P.M.; Holleran, W.M. UVB irradiation up-regulates serine palmitoyltransferase in cultured human keratinocytes. J. Lipid Res. 1998, 39, 2031–2038. [Google Scholar] [CrossRef]
- Uchida, Y.; Di Nardo, A.; Collins, V.; Elias, P.M.; Holleran, W.M. De novo ceramide synthesis participates in the ultraviolet B irradiation-induced apoptosis in undifferentiated cultured human keratinocytes. J. Investig. Dermatol. 2003, 120, 662–669. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Kim, Y.; Lee, Y.; Chung, J.H. Ceramide accelerates ultraviolet-induced MMP-1 expression through JAK1/STAT-1 pathway in cultured human dermal fibroblasts. J. Lipid Res. 2008, 49, 2571–2581. [Google Scholar] [CrossRef] [Green Version]
- Rozenova, K.A.; Deevska, G.M.; Karakashian, A.A.; Nikolova-Karakashian, M.N. Studies on the role of acid sphingomyelinase and ceramide in the regulation of tumor necrosis factor α (TNFα)-converting enzyme activity and TNFα secretion in macrophages. J. Biol. Chem. 2010, 285, 21103–21113. [Google Scholar] [CrossRef] [Green Version]
- Uchida, Y.; Hara, M.; Nishio, H.; Sidransky, E.; Inoue, S.; Otsuka, F.; Suzuki, A.; Elias, P.M.; Holleran, W.M.; Hamanaka, S. Epidermal sphingomyelins are precursors for selected stratum corneum ceramides. J. Lipid Res. 2000, 41, 2071–2082. [Google Scholar] [CrossRef]
- Grether-Beck, S.; Bonizzi, G.; Schmitt-Brenden, H.; Felsner, I.; Timmer, A.; Sies, H.; Johnson, J.P.; Piette, J.; Krutmann, J. Non-enzymatic triggering of the ceramide signalling cascade by solar UVA radiation. EMBO J. 2000, 19, 5793–5800. [Google Scholar] [CrossRef] [Green Version]
- Grether-Beck, S.; Timmer, A.; Felsner, I.; Brenden, H.; Brammertz, D.; Krutmann, J. Ultraviolet A-induced signaling involves a ceramide-mediated autocrine loop leading to ceramide de novo synthesis. J. Investig. Dermatol. 2005, 125, 545–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, Y. Ceramide signaling in mammalian epidermis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2014, 1841, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, J.G.; Grinstein, S. Sensing Phosphatidylserine in Cellular Membranes. Sensors 2011, 11, 1744–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vance, J.E. Phosphatidylserine and phosphatidyletha-nolamine in mammalian cells: Two metabolically-related aminophospholipids. J. Lipid Res. 2008, 49, 1377–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagan, V.E.; Bayır, H.; Tyurina, Y.Y.; Bolevich, S.B.; Maguire, J.J.; Fadeel, B.; Balasubramanian, K. Elimination of the unnecessary: Intra- and extracellular signaling by anionic phospholipids. Biochem. Biophys. Res. Commun. 2017, 482, 482–490. [Google Scholar] [CrossRef]
- Pristov, J.B.; Jovanović, S.V.; Mitrović, A.; Spasojević, I. UV-irradiation provokes generation of superoxide on cell wall polygalacturonic acid. Physiol. Plant. 2013, 148, 574–581. [Google Scholar] [CrossRef]
- Wang, H.Q.; Quan, T.; He, T.; Franke, T.F.; Voorhees, J.J.; Fisher, G.J. Epidermal Growth Factor Receptor-dependent, NF-κB-independent Activation of the Phosphatidylinositol 3-Kinase/Akt Pathway Inhibits Ultraviolet Irradiation-induced Caspases-3, -8, and -9 in Human Keratinocytes. J. Biol. Chem. 2003, 278, 45737–45745. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.B.; Jerome, D.; Yeung, J. Diagnosis and management of psoriasis. Can. Fam. Physician 2017, 63, 278–285. [Google Scholar]
- Zhang, P.; Wu, M.X. A clinical review of phototherapy for psoriasis. Lasers Med. Sci. 2018, 33, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Milde, J.; Elstner, E.F.; Graßmann, J. Synergistic inhibition of low-density lipoprotein oxidation by rutin, γ-terpinene, and ascorbic acid. Phytomedicine 2004, 11, 105–113. [Google Scholar] [CrossRef]
- Serrano, G.; Almudéver, P.; Serrano, J.M.; Milara, J.; Torrens, A.; Expósito, I.; Cortijo, J. Phosphatidylcholine liposomes as carriers to improve topical ascorbic acid treatment of skin disorders. Clin. Cosmet. Investig. Dermatol. 2015, 8, 591–599. [Google Scholar]
- Babazadeh, A.; Ghanbarzadeh, B.; Hamishehkar, H. Phosphatidylcholine-rutin complex as a potential nanocarrier for food applications. J. Funct. Foods 2017, 33, 134–141. [Google Scholar] [CrossRef]
- Ravi, G.S.; Charyulu, R.N.; Dubey, A.; Prabhu, P.; Hebbar, S.; Mathias, A.C. Nano-lipid Complex of Rutin: Development, Characterisation and In Vivo Investigation of Hepatoprotective, Antioxidant Activity and Bioavailability Study in Rats. AAPS PharmSciTech 2018, 19, 3631–3649. [Google Scholar] [CrossRef]
- Kim, K.P.; Shin, K.O.; Park, K.; Yun, H.J.; Mann, S.; Lee, Y.M.; Cho, Y. Vitamin C stimulates epidermal ceramide production by regulating its metabolic enzymes. Biomol. Ther. 2015, 23, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Uchida, Y.; Behne, M.; Quiec, D.; Elias, P.M.; Holleran, W.M. Vitamin C stimulates sphingolipid production and markers of barrier formation in submerged human keratinocyte cultures. J. Investig. Dermatol. 2001, 117, 1307–1313. [Google Scholar] [CrossRef] [Green Version]
- Mizutani, Y.; Mitsutake, S.; Tsuji, K.; Kihara, A.; Igarashi, Y. Ceramide biosynthesis in keratinocyte and its role in skin function. Biochimie 2009, 91, 784–790. [Google Scholar] [CrossRef]
- Pettus, B.J.; Chalfant, C.E.; Hannun, Y.A. Ceramide in apoptosis: An overview and current perspectives. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2002, 1585, 114–125. [Google Scholar] [CrossRef]
- Kippenberger, S.; Loitsch, S.M.; Grundmann-Kollmann, M.; Simon, S.; Dang, T.A.; Hardt-Weinelt, K.; Kaufmann, R.; Bernd, A. Activators of peroxisome proliferator-activated receptors protect human skin from ultraviolet-B-light-induced inflammation. J. Invest. Dermatol. 2001, 117, 1430–1436. [Google Scholar] [CrossRef] [Green Version]
- Siskind, L.J. Mitochondrial ceramide and the induction of apoptosis. J. Bioenerg. Biomembr. 2005, 37, 143–153. [Google Scholar] [CrossRef]
- Stancevic, B.; Kolesnick, R. Ceramide-rich platforms in transmembrane signaling. FEBS Lett. 2010, 584, 1728–1740. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phospholipid Species | Log2 (Fold-Change) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rut vs. Ctr | Asc vs. Ctr | Rut+Asc vs. Ctr | UVA vs. Ctr | UVB vs. Ctr | UVA+Rut vs. UVA | UVA+Asc vs. UVA | UVA+Rut+Asc vs. UVA | UVB+Rut vs. UVB | UVB+Asc vs. UVB | UVB+Rut +Asc vs. UVB | |
| PCp(44:9)/PCo(44:10) | - | - | 1.08 ↑ | 0.58 ↑ | 1.00 ↑ | - | - | - | 1.94 ↑ | 2.08↑ | 2.90↑ |
| PCp(44:11)/PCo(44:12) | - | - | 2.13↑ | 1.26 ↑ | 1.86 ↑ | - | - | - | 1.88 ↑ | 1.15 ↑ | 2.68↑ |
| PC(40:6) | - | - | 1.69 ↑ | 2.05↑ | 2.76↑ | - | - | - | 0.70 ↓ | 0.51 ↑ | 2.34↑ |
| PC(38:3) | - | - | 2.34↑ | 2.42↑ | 3.49↑ | - | - | - | 0.64 ↑ | 0.53 ↑ | 1.68 ↑ |
| PS(40:1) | - | - | - | 1.31 ↑ | 2.15↓ | - | - | - | - | - | 4.98↑ |
| PS(44:4) | - | - | - | 1.20 ↑ | 2.42↓ | - | - | - | - | - | 4.93↑ |
| SM(d40:1) | 1.09 ↓ | 1.36 ↓ | 1.88 ↓ | 2.25↓ | 1.18 ↓ | - | - | 4.46↓ | 2.85↓ | 0.59 ↓ | 3.99↓ |
| PI(40:10) | - | - | - | 0.57 ↑ | 0.90 ↓ | 0.62 ↓ | 0.66 ↓ | 1.22 ↑ | 1.71 ↓ | 2.42↓ | 2.80↓ |
| PI(40:3) | - | - | - | 1.15 ↓ | 2.94↓ | 0.86 ↑ | - | 2.76↑ | 0.88 ↓ | 0.57 ↓ | 1.34 ↓ |
| PI(40:8) | - | - | - | 1.00 ↑ | 0.93 ↓ | 0.81 ↓ | 2.89↓ | 2.53↑ | 2.34↓ | 1.97 ↓ | 1.95 ↓ |
| CER Class | Ceramide Species | Log2 (Fold-Change) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rut vs. Ctr | Asc vs. Ctr | Rut +Asc vs. Ctr | UVA vs. Ctr | UVB vs. Ctr | UVA +Rut vs. UVA | UVA +Asc vs. UVA | UVA +Rut +Asc vs. UVA | UVB +Rut vs. UVB | UVB +Asc vs. UVB | UVB +Rut +Asc vs. UVB | ||
| CER[ADS] | Cer(d18:0/20:0(2OH)) | 2.51↑ | 3.57↑ | - | - | 3.81↑ | - | 3.54↑ | 1.85 ↓ | 5.40↓ | 0.79 ↑ | 2.13↑ |
| CER[ADS] | Cer(d18:0/22:0(2OH)) | 0.48 ↓ | 1.50 ↓ | 0.65 ↓ | 0.31 ↑ | 2.72↓ | - | 3.51↓ | - | 2.68↑ | - | 2.22↑ |
| CER[AS] | Cer(d16:2/24:0(2OH)) | 0.75 ↓ | 1.95 ↓ | 0.62 ↓ | 1.17 ↓ | - | 1.75 ↑ | - | 0.98 ↓ | 1.02 ↑ | 0.28 ↑ | 0.86 ↑ |
| CER[NP] | Cer(t18:0/22:0) | - | - | - | 0.75 ↓ | - | 0.28 ↓ | 1.32 ↓ | 4.39↑ | - | - | 1.70 ↑ |
| CER[NS] | Cer(d18:2/15:0) | - | - | 1.86 ↑ | 3.10↑ | 1.15 ↓ | 2.77↓ | 5.36↓ | 1.60 ↓ | - | - | 2.84↑ |
| CER[NS] | Cer(d18:1/24:0) | - | - | 1.79 ↑ | 1.69 ↑ | 2.34↓ | 1.30 ↓ | 1.13 ↓ | 1.85 ↑ | 3.54↑ | - | 4.47↑ |
| CER[NS] | Cer(d18:1/17:0) | - | - | 0.86 ↑ | 1.13 ↑ | - | 2.12 ↓ | 1.47 ↓ | 2.82↑ | 2.37↑ | - | 2.92↑ |
| CER[NDS] | Cer(d18:0/18:0) | - | - | 1.79 ↑ | 0.58 ↓ | 1.86 ↑ | 1.99 ↓ | 0.45 ↑ | 4.03↑ | - | 2.77↓ | 0.32 ↑ |
| CER[NDS] | Cer(d18:0/13:0) | - | 2.39↓ | 0.94 ↑ | 1.70 ↑ | 0.79 ↑ | 2.71↓ | 2.29↓ | 2.27↑ | 1.24 ↑ | 0.60 ↓ | 2.28↑ |
| CER[NDS] | Cer(d18:0/20:0) | - | - | - | - | - | - | - | 1.97 ↑ | 1.17 ↑ | 1.86 ↓ | 2.66↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gęgotek, A.; Łuczaj, W.; Skrzydlewska, E. Effects of Natural Antioxidants on Phospholipid and Ceramide Profiles of 3D-Cultured Skin Fibroblasts Exposed to UVA or UVB Radiation. Antioxidants 2021, 10, 578. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040578
Gęgotek A, Łuczaj W, Skrzydlewska E. Effects of Natural Antioxidants on Phospholipid and Ceramide Profiles of 3D-Cultured Skin Fibroblasts Exposed to UVA or UVB Radiation. Antioxidants. 2021; 10(4):578. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040578
Chicago/Turabian StyleGęgotek, Agnieszka, Wojciech Łuczaj, and Elżbieta Skrzydlewska. 2021. "Effects of Natural Antioxidants on Phospholipid and Ceramide Profiles of 3D-Cultured Skin Fibroblasts Exposed to UVA or UVB Radiation" Antioxidants 10, no. 4: 578. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040578