
Melatonin Ameliorates Autophagy Impairment in a Metabolic Syndrome Model

 , and
, and 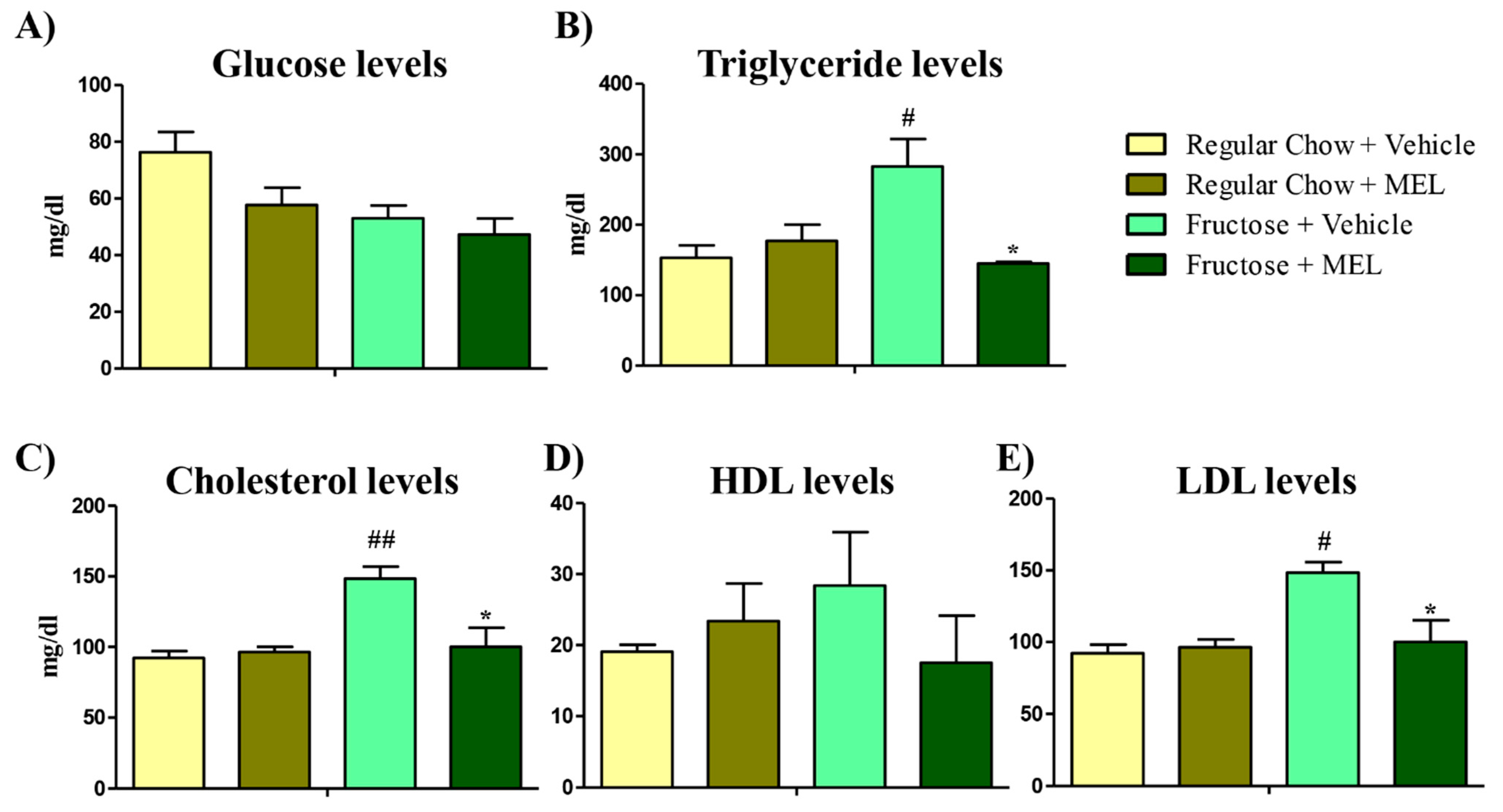{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Isolation of Proteins
2.3. Lipid Peroxidation
2.4. Total Antioxidant Activity (TAA)
2.5. Immunoblotting
2.6. Statistical Analysis
3. Results and Discussion
3.1. Melatonin Treatment Ameliorates the Lipid Effect Caused by a Fructose Diet
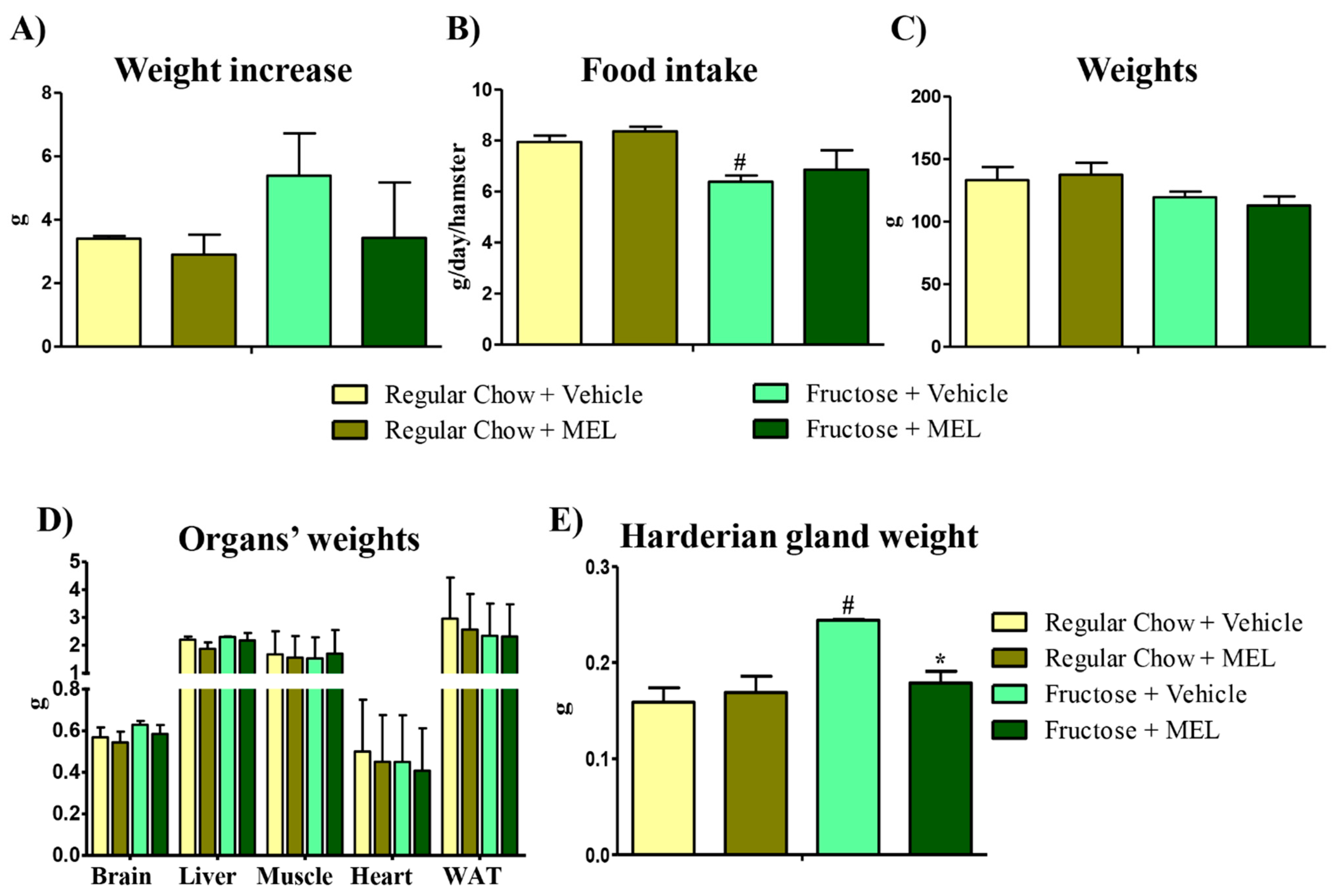
3.2. The Harderian Gland Is the Organ Most Affected by a Fructose Diet
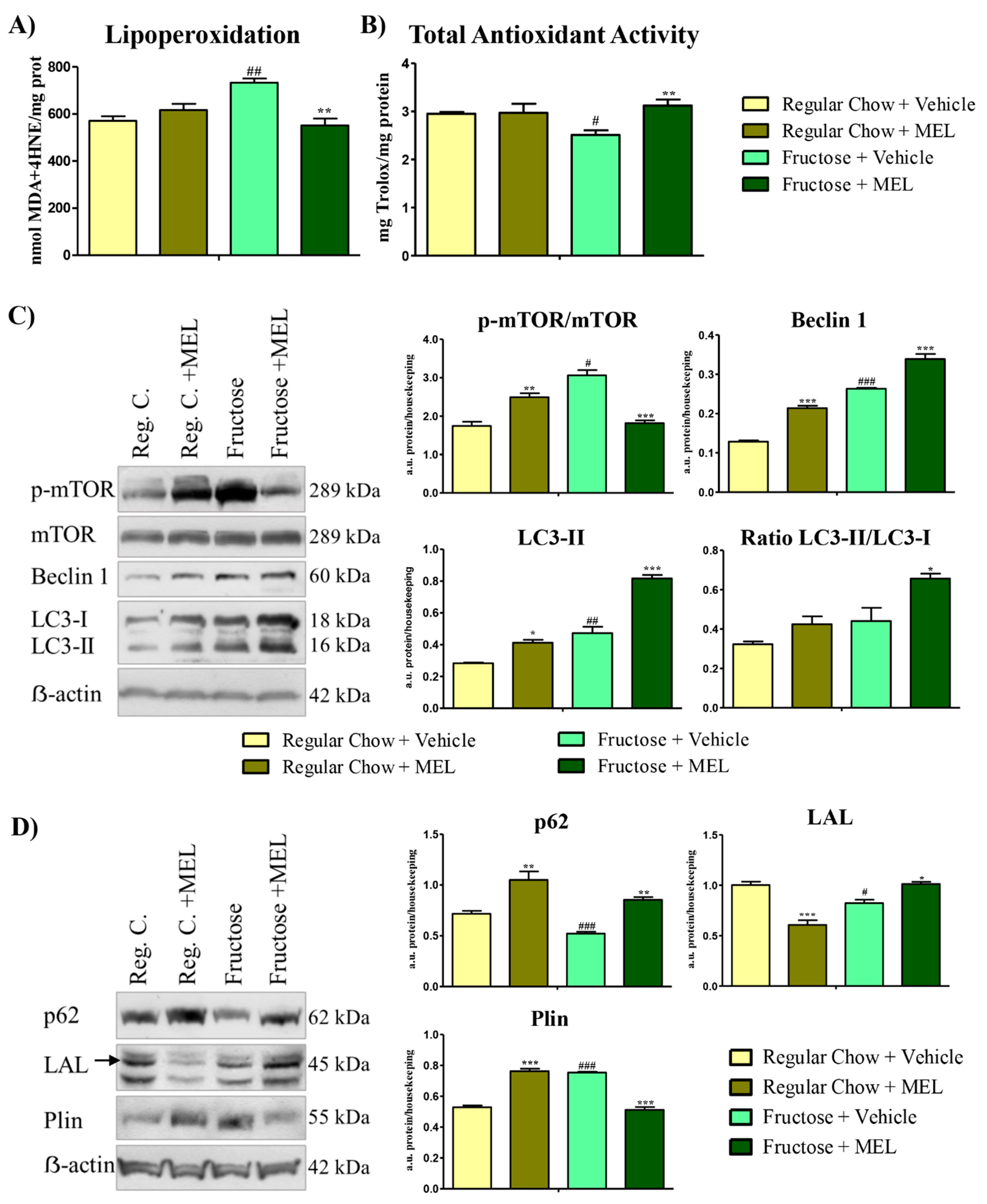
3.3. Melatonin Ameliorates Autophagic and Lipophagic Impairment in the Harderian Gland Caused by the Metabolic Syndrome.
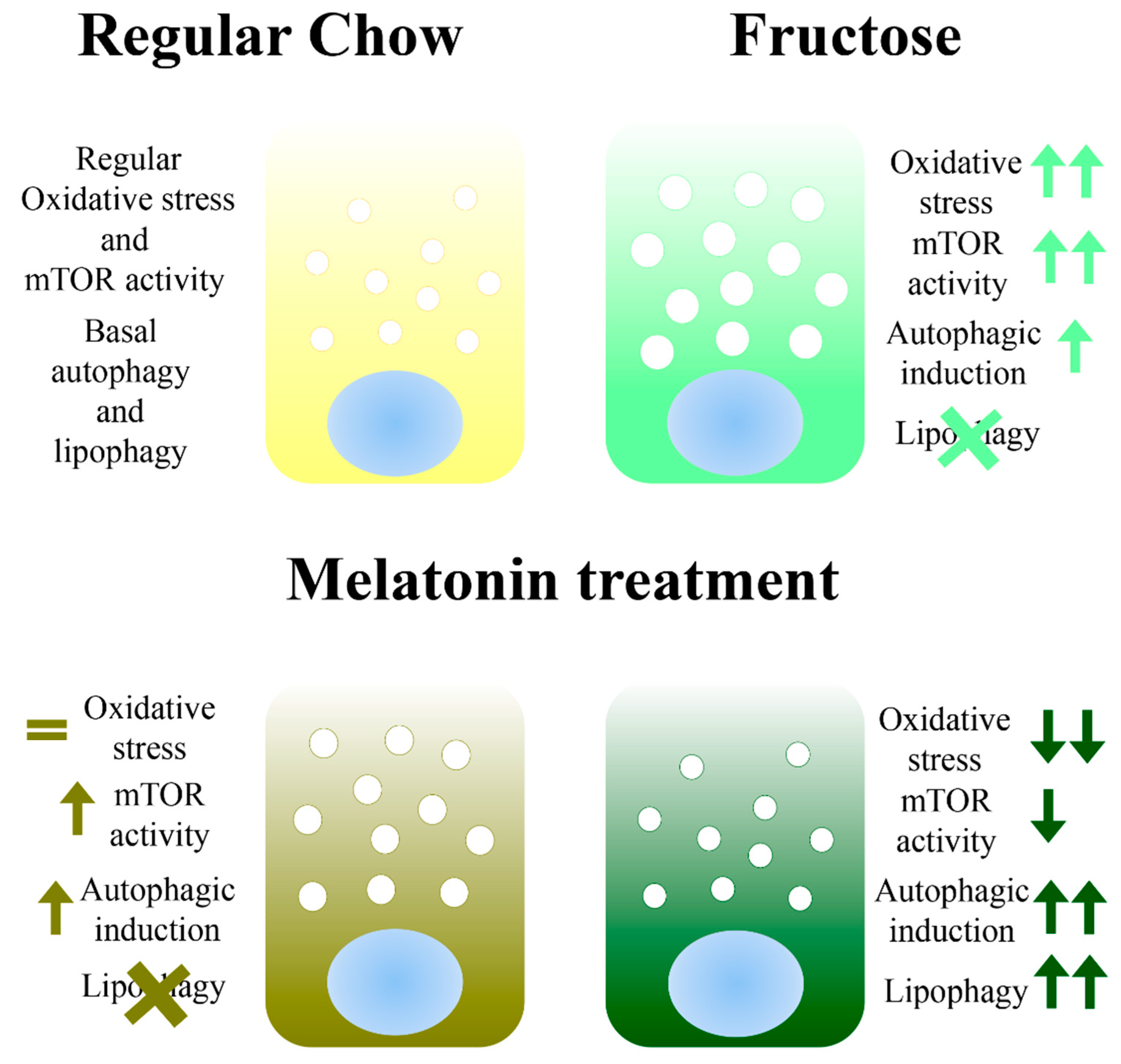
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agil, A.; Navarro-Alarcon, M.; Ruiz, R.; Abuhamadah, S.; El-Mir, M.Y.; Vazquez, G.F. Beneficial effects of melatonin on obesity and lipid profile in young Zucker diabetic fatty rats. J. Pineal Res. 2011, 50, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Demirtas, C.Y.; Pasaoglu, O.T.; Bircan, F.S.; Kantar, S.; Turkozkan, N. The investigation of melatonin effect on liver antioxidant and oxidant levels in fructose-mediated metabolic syndrome model. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1915–1921. [Google Scholar]
- Kozirog, M.; Poliwczak, A.R.; Duchnowicz, P.; Koter-Michalak, M.; Sikora, J.; Broncel, M. Melatonin treatment improves blood pressure, lipid profile, and parameters of oxidative stress in patients with metabolic syndrome. J. Pineal Res. 2011, 50, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J. The pineal and its hormones in the control of reproduction in mammals. Endocr. Rev. 1980, 1, 109–131. [Google Scholar] [CrossRef] [PubMed]
- Negi, G.; Kumar, A.; Sharma, S.S. Melatonin modulates neuroinflammation and oxidative stress in experimental diabetic neuropathy: Effects on NF-kappaB and Nrf2 cascades. J. Pineal Res. 2011, 50, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, A.; Ohta, Y.; Ohashi, K. Melatonin improves metabolic syndrome induced by high fructose intake in rats. J. Pineal Res. 2012, 52, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Rios-Lugo, M.J.; Cano, P.; Jimenez-Ortega, V.; Fernandez-Mateos, M.P.; Scacchi, P.A.; Cardinali, D.P.; Esquifino, A.I. Melatonin effect on plasma adiponectin, leptin, insulin, glucose, triglycerides and cholesterol in normal and high fat-fed rats. J. Pineal Res. 2010, 49, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.; Reiter, R.J. Melatonin: Roles in influenza, Covid-19, and other viral infections. Rev. Med. Virol. 2020, 30, e2109. [Google Scholar] [CrossRef] [PubMed]
- Balakumar, M.; Raji, L.; Prabhu, D.; Sathishkumar, C.; Prabu, P.; Mohan, V.; Balasubramanyam, M. High-fructose diet is as detrimental as high-fat diet in the induction of insulin resistance and diabetes mediated by hepatic/pancreatic endoplasmic reticulum (ER) stress. Mol. Cell. Biochem. 2016, 423, 93–104. [Google Scholar] [CrossRef]
- Sakai, T. The mammalian Harderian gland: Morphology, biochemistry, function and phylogeny. Arch. Histol. Jpn. 1981, 44, 299–333. [Google Scholar] [CrossRef] [Green Version]
- Payne, A.P.; McGadey, J.; Moore, M.H.; Thompson, G.G. Changes in Harderian gland activity in the female golden hamster during the oestrous cycle, pregnancy and lactation. Biochem. J. 1979, 178, 597–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, R.A.; Johnson, L.B.; Reiter, R.J. Harderian glands of golden hamsters: Temporal and sexual differences in immunoreactive melatonin. J. Pineal Res. 1985, 2, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Pelaez, A.; Lopez-Gonzalez, M.A.; Guerrero, J.M. Melatonin binding sites in the Harderian gland of Syrian hamsters: Sexual differences and effect of castration. J. Pineal Res. 1993, 14, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Santillo, A.; Chieffi Baccari, G.; Minucci, S.; Falvo, S.; Venditti, M.; Di Matteo, L. The Harderian gland: Endocrine function and hormonal control. Gen. Comp. Endocrinol. 2020, 297, 113548. [Google Scholar] [CrossRef]
- Coto-Montes, A.; Boga, J.A.; Tomas-Zapico, C.; Rodriguez-Colunga, M.J.; Martinez-Fraga, J.; Tolivia-Cadrecha, D.; Menendez, G.; Hardeland, R.; Tolivia, D. Porphyric enzymes in hamster Harderian gland, a model of damage by porphyrins and their precursors. A chronobiological study on the role of sex differences. Chem. Biol. Interact. 2001, 134, 135–149. [Google Scholar] [CrossRef]
- Vega-Naredo, I.; Caballero, B.; Sierra, V.; Huidobro-Fernandez, C.; de Gonzalo-Calvo, D.; Garcia-Macia, M.; Tolivia, D.; Rodriguez-Colunga, M.J.; Coto-Montes, A. Sexual dimorphism of autophagy in Syrian hamster Harderian gland culminates in a holocrine secretion in female glands. Autophagy 2009, 5, 1004–1017. [Google Scholar] [CrossRef] [Green Version]
- Vega-Naredo, I.; Coto-Montes, A. Physiological autophagy in the Syrian hamster Harderian gland. Methods Enzymol. 2009, 452, 457–476. [Google Scholar]
- Kasim-Karakas, S.E.; Vriend, H.; Almario, R.; Chow, L.C.; Goodman, M.N. Effects of dietary carbohydrates on glucose and lipid metabolism in golden Syrian hamsters. J. Lab. Clin. Med. 1996, 128, 208–213. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Garcia-Macia, M.; Vega-Naredo, I.; De Gonzalo-Calvo, D.; Rodriguez-Gonzalez, S.M.; Camello, P.J.; Camello-Almaraz, C.; Martin-Cano, F.E.; Rodriguez-Colunga, M.J.; Pozo, M.J.; Coto-Montes, A.M. Melatonin induces neural SOD2 expression independent of the NF-kappaB pathway and improves the mitochondrial population and function in old mice. J. Pineal Res. 2011, 50, 54–63. [Google Scholar] [CrossRef]
- Vega-Naredo, I.; Caballero, B.; Sierra, V.; Garcia-Macia, M.; de Gonzalo-Calvo, D.; Oliveira, P.J.; Rodriguez-Colunga, M.J.; Coto-Montes, A. Melatonin modulates autophagy through a redox-mediated action in female Syrian hamster Harderian gland controlling cell types and gland activity. J. Pineal Res. 2012, 52, 80–92. [Google Scholar] [CrossRef]
- Vega-Naredo, I.; Poeggeler, B.; Sierra-Sanchez, V.; Caballero, B.; Tomas-Zapico, C.; Alvarez-Garcia, O.; Tolivia, D.; Rodriguez-Colunga, M.J.; Coto-Montes, A. Melatonin neutralizes neurotoxicity induced by quinolinic acid in brain tissue culture. J. Pineal Res. 2005, 39, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Dalboge, L.S.; Pedersen, P.J.; Hansen, G.; Fabricius, K.; Hansen, H.B.; Jelsing, J.; Vrang, N. A Hamster Model of Diet-Induced Obesity for Preclinical Evaluation of Anti-Obesity, Anti-Diabetic and Lipid Modulating Agents. PLoS ONE 2015, 10, e0135634. [Google Scholar] [CrossRef] [Green Version]
- Horne, R.G.; Yu, Y.; Zhang, R.; Abdalqadir, N.; Rossi, L.; Surette, M.; Sherman, P.M.; Adeli, K. High Fat-High Fructose Diet-Induced Changes in the Gut Microbiota Associated with Dyslipidemia in Syrian Hamsters. Nutrients 2020, 12, 3557. [Google Scholar] [CrossRef] [PubMed]
- Malo, E.; Saukko, M.; Santaniemi, M.; Hietaniemi, M.; Lammentausta, E.; Blanco Sequeiros, R.; Ukkola, O.; Kesaniemi, Y.A. Plasma lipid levels and body weight altered by intrauterine growth restriction and postnatal fructose diet in adult rats. Pediatr. Res. 2013, 73, 155–162. [Google Scholar] [CrossRef]
- Hussain, S.A. Effect of melatonin on cholesterol absorption in rats. J. Pineal Res. 2007, 42, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.Y.; Tang, P.L. Effect of melatonin on the maintenance of cholesterol homeostasis in the rat. Endocr. Res. 1995, 21, 681–696. [Google Scholar] [CrossRef]
- Martinez-Lopez, N.; Garcia-Macia, M.; Sahu, S.; Athonvarangkul, D.; Liebling, E.; Merlo, P.; Cecconi, F.; Schwartz, G.J.; Singh, R. Autophagy in the CNS and Periphery Coordinate Lipophagy and Lipolysis in the Brown Adipose Tissue and Liver. Cell Metab. 2016, 23, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Macia, M.; Santos-Ledo, A.; Caballero, B.; Rubio-Gonzalez, A.; de Luxan-Delgado, B.; Potes, Y.; Rodriguez-Gonzalez, S.M.; Boga, J.A.; Coto-Montes, A. Selective autophagy, lipophagy and mitophagy, in the Harderian gland along the oestrous cycle: A potential retrieval effect of melatonin. Sci. Rep. 2019, 9, 18597. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Vigo, D.E. Melatonin, mitochondria, and the metabolic syndrome. Cell. Mol. Life Sci. 2017, 74, 3941–3954. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Macia, M.; Rubio-Gonzalez, A.; de Luxan-Delgado, B.; Potes, Y.; Rodriguez-Gonzalez, S.; de Gonzalo-Calvo, D.; Boga, J.A.; Coto-Montes, A. Autophagic and proteolytic processes in the Harderian gland are modulated during the estrous cycle. Histochem. Cell Biol. 2014, 141, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Karsli-Uzunbas, G.; Poillet-Perez, L.; Sawant, A.; Hu, Z.S.; Zhao, Y.; Moore, D.; Hu, W.; White, E. Autophagy promotes mammalian survival by suppressing oxidative stress and p53. Genes Dev. 2020, 34, 688–700. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, A.; Amiri, F.; Khalighi, F.; Mohammadi Roushandeh, A.; Kuwahara, Y.; Bashiri, H.; Habibi Roudkenar, M. Cell Survival Effects of Autophagy Regulation on Umbilical Cord-Derived Mesenchymal Stem Cells Following Exposure to Oxidative Stress. Iran. J. Med. Sci. 2019, 44, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Rabanal-Ruiz, Y.; Korolchuk, V.I. mTORC1 and Nutrient Homeostasis: The Central Role of the Lysosome. Int. J. Mol. Sci. 2018, 19, 818. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, D.Z.; El-Sisi, A.E.E.; Sokar, S.S.; Shebl, A.M.; Abu-Risha, S.E. Targeting autophagy to modulate hepatic ischemia/reperfusion injury: A comparative study between octreotide and melatonin as autophagy modulators through AMPK/PI3K/AKT/mTOR/ULK1 and Keap1/Nrf2 signaling pathways in rats. Eur. J. Pharmacol. 2021, 897, 173920. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Yang, M.; Wang, Y.; Ren, J.; Lin, P.; Cui, C.; Song, J.; He, Q.; Hu, H.; Wang, K.; et al. Melatonin prevents diabetes-associated cognitive dysfunction from microglia-mediated neuroinflammation by activating autophagy via TLR4/Akt/mTOR pathway. FASEB J. 2021, 35, e21485. [Google Scholar] [CrossRef]
- Zoncu, R.; Efeyan, A.; Sabatini, D.M. mTOR: From growth signal integration to cancer, diabetes and ageing. Nat. Rev. Mol. Cell Biol. 2011, 12, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef] [Green Version]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition). Autophagy 2021, 1–382. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos-Ledo, A.; Luxán-Delgado, B.d.; Caballero, B.; Potes, Y.; Rodríguez-González, S.; Boga, J.A.; Coto-Montes, A.; García-Macia, M. Melatonin Ameliorates Autophagy Impairment in a Metabolic Syndrome Model. Antioxidants 2021, 10, 796. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050796
Santos-Ledo A, Luxán-Delgado Bd, Caballero B, Potes Y, Rodríguez-González S, Boga JA, Coto-Montes A, García-Macia M. Melatonin Ameliorates Autophagy Impairment in a Metabolic Syndrome Model. Antioxidants. 2021; 10(5):796. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050796
Chicago/Turabian StyleSantos-Ledo, Adrián, Beatriz de Luxán-Delgado, Beatriz Caballero, Yaiza Potes, Susana Rodríguez-González, José Antonio Boga, Ana Coto-Montes, and Marina García-Macia. 2021. "Melatonin Ameliorates Autophagy Impairment in a Metabolic Syndrome Model" Antioxidants 10, no. 5: 796. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050796