
Methionine Sulfoxide Reductases Contribute to Anaerobic Fermentative Metabolism in Bacillus cereus

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Growth Conditions
2.2. Analytical Procedures
2.3. Relative Quantification of msrA and msrAB Gene Expression
2.4. Construction of msrA Mutant Strains
2.5. Proteomics Analysis
2.6. Motility Assays
2.7. Statistical Analyses
3. Results
3.1. Expression Patterns for B. cereus msrA and msrAB RNA and Proteins under Fermentative Anaerobic Conditions
3.2. Effect of msrA and msrAB Mutations on Anaerobic Fermentative Metabolism
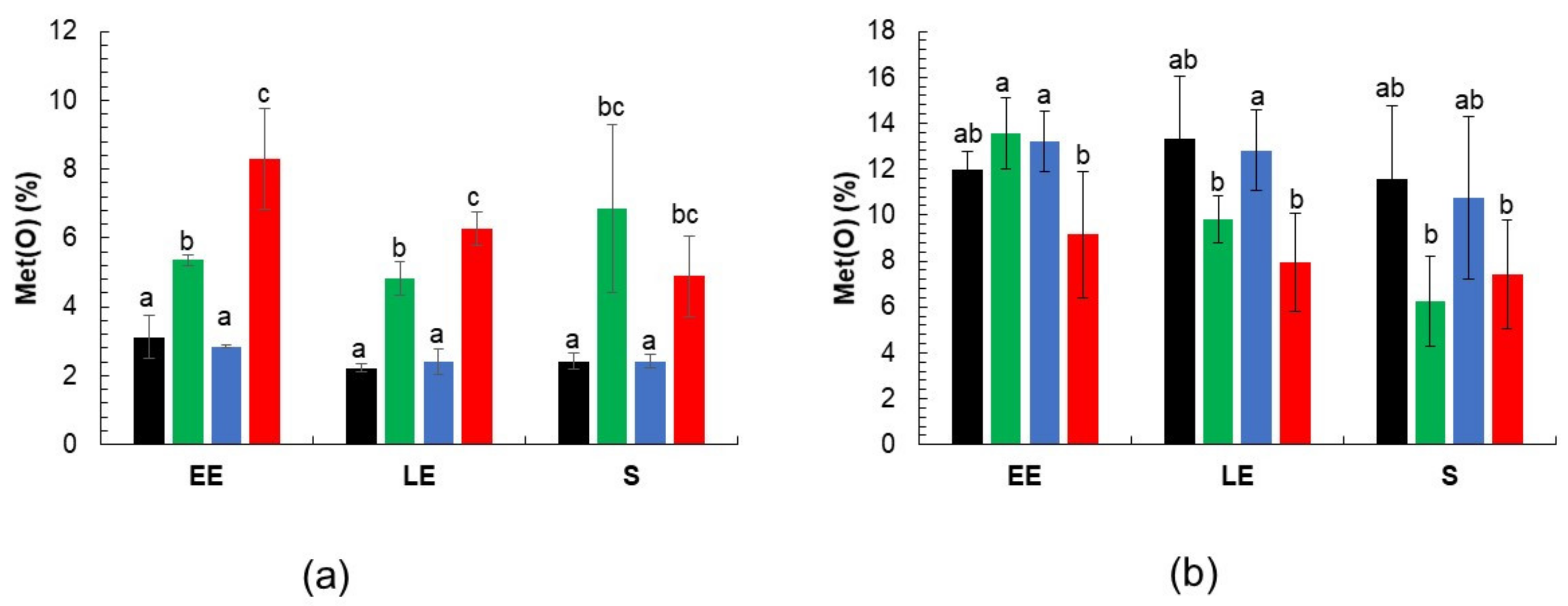
3.3. Met(O) Accumulation in msr Mutant Proteomes
3.4. Identification of Putative Msr Substrates
3.4.1. Putative Msr Substrates Identified in B. cereus Cellular Proteome
3.4.2. Putative Msr Substrates Identified in B. cereus Exoproteome
3.5. Swimming Motility of B. cereus Depends on MsrA Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duport, C.; Jobin, M.; Schmitt, P. Adaptation in Bacillus cereus: From stress to disease. Front. Microbiol. 2016, 7, 1550. [Google Scholar] [CrossRef] [Green Version]
- Laouami, S.; Messaoudi, K.; Alberto, F.; Clavel, T.; Duport, C. Lactate dehydrogenase A promotes communication between carbohydrate catabolism and virulence in Bacillus cereus. J. Bacteriol. 2011, 193, 1757–1766. [Google Scholar] [CrossRef] [Green Version]
- Rosenfeld, E.; Duport, C.; Zigha, A.; Schmitt, P. Characterization of aerobic and anaerobic vegetative growth of the food-borne pathogen Bacillus cereus F4430/73 strain. Can. J. Microbiol. 2005, 51, 149–158. [Google Scholar] [CrossRef]
- Zigha, A.; Rosenfeld, E.; Schmitt, P.; Duport, C. Anaerobic cells of Bacillus cereus F4430/73 respond to low oxidoreduction potential by metabolic readjustments and activation of enterotoxin expression. Arch. Microbiol. 2006, 185, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Clair, G.; Armengaud, J.; Duport, C. Restricting fermentative potential by proteome remodeling: An adaptive strategy evidenced in Bacillus cereus. Mol. Cell Proteom. 2012, 11, M111-013102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, J.P.; Alpha-Bazin, B.; Armengaud, J.; Duport, C. Time dynamics of the Bacillus cereus exoproteome are shaped by cellular oxidation. Front. Microbiol. 2015, 6, 342. [Google Scholar] [CrossRef] [PubMed]
- Mols, M.; Abee, T. Primary and secondary oxidative stress in Bacillus. Environ. Microbiol. 2011, 13, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Mols, M.; van Kranenburg, R.; van Melis, C.C.; Moezelaar, R.; Abee, T. Analysis of acid-stressed Bacillus cereus reveals a major oxidative response and inactivation-associated radical formation. Environ. Microbiol. 2010, 12, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, H.J.; Dupont, A.L.; Leivo, H.E.; Guillemette, J.G. Cloning, expression, and purification of a nitric oxide synthase-like protein from Bacillus cereus. Biochem. Res. Int. 2010, 2010, 489892. [Google Scholar] [CrossRef] [Green Version]
- Popova, T.G.; Teunis, A.; Vaseghi, H.; Zhou, W.; Espina, V.; Liotta, L.A.; Popov, S.G. Nitric oxide as a regulator of B. anthracis pathogenicity. Front. Microbiol. 2015, 6, 921. [Google Scholar] [CrossRef]
- Luo, S.; Levine, R.L. Methionine in proteins defends against oxidative stress. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2009, 23, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Achilli, C.; Ciana, A.; Minetti, G. The discovery of methionine sulfoxide reductase enzymes: An historical account and future perspectives. BioFactors 2015, 41, 135–152. [Google Scholar] [CrossRef]
- Tarrago, L.; Grosse, S.; Lemaire, D.; Faure, L.; Tribout, M.; Siponen, M.I.; Kojadinovic-Sirinelli, M.; Pignol, D.; Arnoux, P.; Sabaty, M. Reduction of protein bound methionine sulfoxide by a periplasmic dimethyl sulfoxide reductase. Antioxidants 2020, 9, 616. [Google Scholar] [CrossRef]
- Brot, N.; Weissbach, L.; Werth, J.; Weissbach, H. Enzymatic reduction of protein-bound methionine sulfoxide. Proc. Natl. Acad. Sci. USA 1981, 78, 2155–2158. [Google Scholar] [CrossRef] [Green Version]
- Grimaud, R.; Ezraty, B.; Mitchell, J.K.; Lafitte, D.; Briand, C.; Derrick, P.J.; Barras, F. Repair of oxidized proteins. Identification of a new methionine sulfoxide reductase. J. Biol. Chem. 2001, 276, 48915–48920. [Google Scholar] [CrossRef] [Green Version]
- Tarrago, L.; Grosse, S.; Siponen, M.I.; Lemaire, D.; Alonso, B.; Miotello, G.; Armengaud, J.; Arnoux, P.; Pignol, D.; Sabaty, M. Rhodobacter sphaeroides methionine sulfoxide reductase P reduces R- and S-diastereomers of methionine sulfoxide from a broad-spectrum of protein substrates. Biochem. J. 2018, 475, 3779–3795. [Google Scholar] [CrossRef] [Green Version]
- Tarrago, L.; Kaya, A.; Weerapana, E.; Marino, S.M.; Gladyshev, V.N. Methionine sulfoxide reductases preferentially reduce unfolded oxidized proteins and protect cells from oxidative protein unfolding. J. Biol. Chem. 2012, 287, 24448–24459. [Google Scholar] [CrossRef] [Green Version]
- Laugier, E.; Tarrago, L.; Vieira Dos Santos, C.; Eymery, F.; Havaux, M.; Rey, P. Arabidopsis thaliana plastidial methionine sulfoxide reductases B, MSRBs, account for most leaf peptide MSR activity and are essential for growth under environmental constraints through a role in the preservation of photosystem antennae. Plant J. Cell Mol. Biol. 2010, 61, 271–282. [Google Scholar] [CrossRef]
- Lourenco Dos Santos, S.; Petropoulos, I.; Friguet, B. The oxidized protein repair enzymes methionine sulfoxide reductases and their roles in protecting against oxidative stress, in ageing and in regulating protein function. Antioxidants 2018, 7, 191. [Google Scholar] [CrossRef]
- Salmon, A.B.; Perez, V.I.; Bokov, A.; Jernigan, A.; Kim, G.; Zhao, H.; Levine, R.L.; Richardson, A. Lack of methionine sulfoxide reductase A in mice increases sensitivity to oxidative stress but does not diminish life span. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2009, 23, 3601–3608. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.J.; Han, Z.Q.; Li, Z.Y. Modulating protein activity and cellular function by methionine residue oxidation. Amino Acids 2012, 43, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Slesak, I.; Slesak, H.; Zimak-Piekarczyk, P.; Rozpadek, P. Enzymatic antioxidant systems in early anaerobes: Theoretical considerations. Astrobiology 2016, 16, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Weissbach, H. Origin and evolution of the protein-repairing enzymes methionine sulphoxide reductases. Biol. Rev. Camb. Philos. Soc. 2008, 83, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Koc, A.; Gasch, A.P.; Rutherford, J.C.; Kim, H.Y.; Gladyshev, V.N. Methionine sulfoxide reductase regulation of yeast lifespan reveals reactive oxygen species-dependent and -independent components of aging. Proc. Natl. Acad. Sci. USA 2004, 101, 7999–8004. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.Y.; Zhang, Y.; Lee, B.C.; Kim, J.R.; Gladyshev, V.N. The selenoproteome of Clostridium sp. OhILAs: Characterization of anaerobic bacterial selenoprotein methionine sulfoxide reductase A. Proteins 2009, 74, 1008–1017. [Google Scholar] [CrossRef] [Green Version]
- Delaye, L.; Becerra, A.; Orgel, L.; Lazcano, A. Molecular evolution of peptide methionine sulfoxide reductases (MsrA and MsrB): On the early development of a mechanism that protects against oxidative damage. J. Mol. Evol. 2007, 64, 15–32. [Google Scholar] [CrossRef]
- Madeira, J.P.; Alpha-Bazin, B.M.; Armengaud, J.; Duport, C. Methionine residues in exoproteins and their recycling by methionine sulfoxide reductase ab serve as an antioxidant strategy in Bacillus cereus. Front. Microbiol. 2017, 8, 1342. [Google Scholar] [CrossRef]
- Madeira, J.P.; Omer, H.; Alpha-Bazin, B.; Armengaud, J.; Duport, C. Deciphering the interactions between the Bacillus cereus linear plasmid, pBClin15, and its host by high-throughput comparative proteomics. J. Proteom. 2016, 146, 25–33. [Google Scholar] [CrossRef]
- Zwietering, M.; Jongenburger, I.; Rombouts, F.; Van’t Riet, K. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef] [Green Version]
- Madeira, J.P.; Alpha-Bazin, B.; Armengaud, J.; Duport, C. Time-course proteomics dataset to monitor protein-bound methionine oxidation in Bacillus cereus ATCC 14579. Data Brief 2018, 18, 394–398. [Google Scholar] [CrossRef]
- Murphy, E. Nucleotide sequence of a spectinomycin adenyltransferase AAD(9) determinant from Staphylococcus aureus and its relationship to AAD(3”) (9). Mol. Gen. Genet. MGG 1985, 200, 33–39. [Google Scholar] [CrossRef]
- Arnaud, M.; Chastanet, A.; Debarbouille, M. New vector for efficient allelic replacement in naturally nontransformable, low-GC-content, gram-positive bacteria. Appl Environ. Microbiol. 2004, 70, 6887–6891. [Google Scholar] [CrossRef] [Green Version]
- Guerout-Fleury, A.M.; Shazand, K.; Frandsen, N.; Stragier, P. Antibiotic-resistance cassettes for Bacillus subtilis. Gene 1995, 167, 335–336. [Google Scholar] [CrossRef]
- Arantes, O.; Lereclus, D. Construction of cloning vectors for Bacillus thuringiensis. Gene 1991, 108, 115–119. [Google Scholar] [CrossRef]
- Hartmann, E.M.; Allain, F.; Gaillard, J.C.; Pible, O.; Armengaud, J. Taking the shortcut for high-throughput shotgun proteomic analysis of bacteria. Methods Mol. Biol. 2014, 1197, 275–285. [Google Scholar] [CrossRef]
- Clair, G.; Roussi, S.; Armengaud, J.; Duport, C. Expanding the known repertoire of virulence factors produced by Bacillus cereus through early secretome profiling in three redox conditions. Mol. Cell Proteom. 2010, 9, 1486–1498. [Google Scholar] [CrossRef] [Green Version]
- De Groot, A.; Dulermo, R.; Ortet, P.; Blanchard, L.; Guerin, P.; Fernandez, B.; Vacherie, B.; Dossat, C.; Jolivet, E.; Siguier, P.; et al. Alliance of proteomics and genomics to unravel the specificities of Sahara bacterium Deinococcus deserti. PLoS Genet. 2009, 5, e1000434. [Google Scholar] [CrossRef]
- Madeira, J.P.; Omer, H.; Alpha-Bazin, B.; Armengaud, J.; Duport, C. Cellular and exoproteome dynamics of Bacillus cereus ATCC 14579 with and without pBClin15. Data Brief. 2016. submitted. [Google Scholar] [CrossRef]
- Dupierris, V.; Masselon, C.; Court, M.; Kieffer-Jaquinod, S.; Bruley, C. A toolbox for validation of mass spectrometry peptides identification and generation of database: IRMa. Bioinformatics 2009, 25, 1980–1981. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Smits, A.H.; van Tilburg, G.B.; Ovaa, H.; Huber, W.; Vermeulen, M. Proteome-wide identification of ubiquitin interactions using UbIA-MS. Nat. Protoc. 2018, 13, 530–550. [Google Scholar] [CrossRef]
- Senesi, S.; Celandroni, F.; Salvetti, S.; Beecher, D.J.; Wong, A.C.L.; Ghelardi, E. Swarming motility in Bacillus cereus and characterization of a fliY mutant impaired in swarm cell differentiation. Microbiology 2002, 148, 1785–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.M.; Coleman, S.; Wilkinson, L.; Smith, M.L.; Hoang, T.; Niyah, N.; Mukherjee, M.; Huynh, S.; Parker, C.T.; Kovac, J.; et al. Unique inducible filamentous motility identified in pathogenic Bacillus cereus group species. ISME J. 2020, 14, 2997–3010. [Google Scholar] [CrossRef]
- Dukan, S.; Nystrom, T. Oxidative stress defense and deterioration of growth-arrested Escherichia coli cells. J. Biol. Chem. 1999, 274, 26027–26032. [Google Scholar] [CrossRef] [Green Version]
- Sigler, P.B.; Xu, Z.; Rye, H.S.; Burston, S.G.; Fenton, W.A.; Horwich, A.L. Structure and function in GroEL-mediated protein folding. Ann. Rev. Biochem. 1998, 67, 581–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khor, H.K.; Fisher, M.T.; Schoneich, C. Potential role of methionine sulfoxide in the inactivation of the chaperone GroEL by hypochlorous acid (HOCl) and peroxynitrite (ONOO-). J. Biol. Chem. 2004, 279, 19486–19493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahawar, M.; Tran, V.; Sharp, J.S.; Maier, R.J. Synergistic roles of Helicobacter pylori methionine sulfoxide reductase and GroEL in repairing oxidant-damaged catalase. J. Biol. Chem. 2011, 286, 19159–19169. [Google Scholar] [CrossRef] [Green Version]
- Harvey, K.L.; Jarocki, V.M.; Charles, I.G.; Djordjevic, S.P. The diverse functional roles of elongation factor Tu (EF-Tu) in microbial pathogenesis. Front. Microbiol. 2019, 10, 2351. [Google Scholar] [CrossRef] [PubMed]
- Trchounian, A. Escherichia coli proton-translocating F0F1-ATP synthase and its association with solute secondary transporters and/or enzymes of anaerobic oxidation-reduction under fermentation. Biochem. Biophys. Res. Commun. 2004, 315, 1051–1057. [Google Scholar] [CrossRef]
- Kaya, A.; Lee, B.C.; Gladyshev, V.N. Regulation of protein function by reversible methionine oxidation and the role of selenoprotein MsrB1. Antioxid. Redox Signal. 2015, 23, 814–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, S.; Minamino, T. Flagella-driven motility of bacteria. Biomolecules 2019, 9, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parish, C.R.; Stanley, P. Chemical and biological properties of bacterial flagellin following iodination and oxidation by chloramine-T. Immunochemistry 1972, 9, 853–872. [Google Scholar] [CrossRef]
- Duport, C.; Rousset, L.; Alpha-Bazin, B.; Armengaud, J. Bacillus cereus decreases NHE and CLO exotoxin synthesis to maintain appropriate proteome dynamics during growth at low temperature. Toxins 2020, 12, 645. [Google Scholar] [CrossRef]
- Senesi, S.; Ghelardi, E. Production, secretion and biological activity of Bacillus cereus enterotoxins. Toxins 2010, 2, 1690–1703. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, K.; Baum, K. The role of methionine sulfoxide reductases in oxidative stress tolerance and virulence of Staphylococcus aureus and other bacteria. Antioxidants 2018, 7, 128. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B. cereus Strains | ||||
|---|---|---|---|---|
| WT | ΔmsrA | ΔmsrAB | ΔmsrAΔmsrAB | |
| µmax (h−1) | 1.0 ± 0.1 a | 1.0 ± 0.1 a | 0.9 ± 0.1 a | 1.0 ± 0.1 a |
| Final OD600 | 2.0 ± 0.1 a | 1.4 ± 0.2 b | 2.0 ± 0.1 a | 1.1 ± 0.1 c |
| Final biomass (g/L) | 0.8 ± 0.1 a | 0.6 ± 0.1 b | 0.8 ± 0.1 a | 0.4 ± 0.1 c |
| qglucose (mmol/g/h) | 77 ± 7 a | 96 ± 4 b | 103 ± 5 b | 168 ± 5 c |
| Yacetate (mol/mol) | 0.31 ± 0.01 a | 0.41 ± 0.01 b | 0.24 ± 0.01 c | 0.65 ± 0.04 d |
| Ylactate (mol/mol) | 1.45 ± 0.06 a | 1.52 ± 0.01 a | 1.57 ± 0.06 a | 1.47 ± 0.42 a |
| Yformate (mol/mol) | 0.39 ± 0.08 a | 0.47 ± 0.01 a | 0.31 ± 0.08 a | 0.44 ± 0.07 a |
| Yethanol (mol/mol) | 0.10 ± 0.00 a | 0.07 ± 0.02 a | 0.10 ± 0.00 a | 0.08 ± 0.02 a |
| Ysuccinate (mol/mol) | 0.04 ± 0.00 a | 0.00 ± 0.00 b | 0.05 ± 0.00 a | 0.00 ± 0.00 b |
| Molecular Function | Gene No | Protein Name | Description | Met(O)/ Total Met | Met(O) Peptide Detected by LC MS/MS a | log2FC b | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| EE | LE | S | |||||||||
| ΔmsrA | ΔmsrAΔmsrAB | ΔmsrA | ΔmsrAΔmsrAB | ΔmsrA | ΔmsrAΔmsrAB | ||||||
| Amino acid metabolism | BC1238 | TrpA | Tryptophan synthase, alpha subunit | 1/5 | EVQMPFVLMTYLNPVLAFGK | 4.2 | |||||
| ATP synthesis | BC5308 | AtpA | F0F1 ATP synthase, alpha subunit | 1/10 | IMQVPVGK | 2.3 | |||||
| Chaperone proteins | BC0295 | GroEL | Chaperonin | 5/20 | SALQNAASVAAMFLTTEAVVADKPEPNAPAMPDMGGMGMGGMGGMM | 3.3 | |||||
| NVTAGANPMGLR | 2.0 | −3.0 | |||||||||
| SSIAQVAAISAADEEVGQLIAEAMER | 3.0 | 3.9 | 4.5 | 4.0 | |||||||
| AMLEDIAILTGGEVITEELGR | 4.5 | 5.0 | 4.3 | 4.6 | 4.7 | 4.3 | |||||
| Degradative enzymes | BC1991 | TgC | Putative murein endopeptidase | 1/9 | YKQSMDGTMQDIKK | −3.3 | |||||
| Fermentation | BC4365 | AdhE | bifunctional acetaldehyde-CoA alcohol dehydrogenase | 4/30 | QLMNHDGVALVLATGGAGMVK | 3.2 | |||||
| MIDTLVNNGQQALQALESFTQEEIDNIVHEMALAGVDQHMPLAK | 2.6 | 3.0 | |||||||||
| LPLISELKEIYMK | 2.8 | 3.2 | 4.3 | ||||||||
| BC4870 | Ldh2 | L-lactate dehydrogenase | 1/7 | GIMDSGFDGIFLIATNPVDILTYVTWK | 3.2 | 4.8 | 3.7 | ||||
| GlycolysisGluconeogenesis | BC4599 | PykI | Pyruvate kinase | 1/16 | AASTDEMLDTAIQTGMDAGLIGLGDTVVITAGVPVAETGTTNLMK | 2.7 | 3.4 | 4.0 | 3.8 | 3.4 | |
| BC4898 c | Pgi | Glucose-6-phosphate isomerase | 2/9 | FSVLTPVGLLPIAVSGLNIEEMMK | 3.1 | ||||||
| FSVLTPVGLLPIAVSGLNIEEMMK | 2.9 | 3.5 | 3.9 | 4.8 | 4.3 | 4.4 | |||||
| BC5135 | Eno | Phosphopyruvate hydratase | 5/11 | QLPTPMMNIINGGSHADNNVDFQEFMILPVGAPTFK | 2.6 | ||||||
| VNQIGTLTETFEAIEMAKR | 3.1 | 3.0 | |||||||||
| QLPTPMMNIINGGSHADNNVDFQEFMILPVGAPTFK | 2.5 | 3.4 | 3.3 | 3.9 | |||||||
| AMIELDGTPNKGK | −2.6 | −2.6 | -2.4 | ||||||||
| LGANAILGVSMAVAHAAADFVGLPLYR | 3.5 | 3.7 | 3.3 | 3.6 | 2.9 | ||||||
| AMIELDGTPNK | −2.9 | ||||||||||
| BC5140 | GapA2 | Glyceraldehyde-3-phosphate dehydrogenase | 4/9 | GMMTTIHSYTNDQQILDLPHK | 3.8 | ||||||
| GILGYSEEPLVSIDYNGCTASSTIDALSTMVMEGNMVK | 3.0 | ||||||||||
| GMMTTIHSYTNDQQILDLPHKDLR | 3.0 | ||||||||||
| GILGYSEEPLVSIDYNGCTASSTIDALSTMVMEGNMVK | 3.0 | 4.1 | 4.0 | ||||||||
| AAAENMIPTSTGAAK | −2.3 | −2.3 | |||||||||
| Protein export | BC4410 | YajC | Preprotein translocase | 1/6 | AVAQMQSELAK | −3.2 | |||||
| Translation apparatus | BC0129 | Tuf | Elongation factor Tu | 10/12 | CDMVDDEELLELVEMEVR | 3.9 | |||||
| CDMVDDEELLELVEMEVRDLLSEYGFPGDDIPVIK | 3.9 | ||||||||||
| ETDKPFLMPVEDVFSITGR | 2.8 | 4.3 | 3.1 | ||||||||
| IIELMAEVDAYIPTPERETDKPFLMPVEDVFSITGR | 4.6 | 4.8 | |||||||||
| IIELMAEVDAYIPTPERETDKPFLMPVEDVFSITGR | 4.2 | 4.2 | 4.4 | 5.1 | |||||||
| NMITGAAQMDGGILVVSAADGPMPQTR | 3.4 | 3.9 | |||||||||
| QVGVPYIVVFLNKCDMVDDEELLELVEMEVR | 3.5 | ||||||||||
| TTDVTGIIQLPEGTEMVMPGDNIEMTIELIAPIAIEEGTK | 3.1 | 3.6 | 3.9 | 4.5 | 4.3 | ||||||
| TTDVTGIIQLPEGTEMVMPGDNIEMTIELIAPIAIEEGTK | 3.1 | 3.1 | 3.4 | 3.9 | 3.4 | ||||||
| TTDVTGIIQLPEGTEMVMPGDNIEMTIELIAPIAIEEGTK | 4.0 | 4.3 | 3.7 | 3.6 | |||||||
| TTDVTGIIQLPEGTEMVMPGDNIEMTIELIAPIAIEEGTK | 3.2 | 3.3 | |||||||||
| TTDVTGIIQLPEGTEMVMPGDNIEMTIELIAPIAIEEGTK | 2.9 | ||||||||||
| VGDVVEIIGLAEENASTTVTGVEMFR | 5.1 | 4.0 | |||||||||
| VGDVVEIIGLAEENASTTVTGVEMFRK | 6.1 | 6.1 | 4.4 | ||||||||
| BC5471 | RplI | 50S ribosomal protein L9 | 1/4 | QGLAAEATNSSMK | −5.0 | −4.6 | −4.7 | −5.6 | |||
| BC0155 | Rpm | 50S ribosomal protein L36 | 1/2 | VMVICENPK | −2.9 | ||||||
| Uncategorized | BC4182 c | Gls24 | Unknown | 1/6 | VEIAPEVIEVIAGIAAAEVEGVAAMR | 2.7 | 3.2 | 3.9 | 3.2 | ||
| Molecular Function | Gene No | Protein Name | Description | Met(O)/ Total Met | Met(O) Peptide Detected by LC MS/MSa | log2FCb | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EE | LE | S | ||||||||||||
| ΔmsrA | ΔmsrAB | ΔmsrAΔmsrAB | ΔmsrA | ΔmsrAB | ΔmsrAΔmsrAB | ΔmsrA | ΔmsrAB | ΔmsrAΔmsrAB | ||||||
| Cell surface biogenesis | BC5196 | CwlD | N-acetylmuramoylL-alanine amidase | 3/8 | MDTVVTSMTSTEGQLKELEK | −1.1 | −1.1 | −1.0 | ||||||
| ILESDEDIMK | −0.4 | −1.5 | −0.7 | |||||||||||
| Chaperone | BC0295 | GroEL | Chaperonin | 1/20 | AMLEDIAILTGGEVITEELGR | 0.8 | 1.5 | 0.2 | ||||||
| Degradative enzymes | BC0556 | ColG | Collagenase | 1/6 | GLEVVTQAMHMYPR | 1.1 | 1.1 | 1.3 | ||||||
| MKGQAIYDIMQGIDYDIQSYLTEAR | 0.0 | 0.0 | 0.0 | 9.4 | 1.2 | 9.9 | ||||||||
| BC0670 | PlcB | Phospholipase C | 1/6 | AEVTPMTGKR | 2.3 | −0.1 | 2.4 | 9.5 | 2.6 | 10.0 | ||||
| BC1193 | PepF1 | Oligoendopeptidase F | 1/11 | ALGLDELHMYDLYTPLVPEVK | 23.0 | 15.0 | 18.9 | |||||||
| BC1991 | TgC | Murein endopeptidase | 1/9 | NIMDQLYGEFNKIVDADEYVK | 9.4 | −0.4 | 8.7 | |||||||
| NIMDQLYGEFNKIVDADEYVKYNVASTR | −1.0 | −0.9 | −1.0 | |||||||||||
| BC2735 | NprP2 | Bacillolysin | 2/10 | GIGEDKMFDIFYYANTDELNMTSNFK | 6.5 | 1.0 | 6.6 | |||||||
| GIGEDKMFDIFYYANTDELNMTSNFK | 7.5 | 1.7 | 7.5 | |||||||||||
| BC5351 | NprB | Bacillolysin | 1/6 | NMSDIYDYFKK | −1.5 | −1.2 | −0.4 | |||||||
| GNGIYIYNANYADSLGGYSQAGYPGTLISSSTPNFADKEAAGAMK | 2.5 | 0.3 | 2.6 | 7.7 | 1.5 | 8.2 | ||||||||
| Flagella | BC1657 c | FlaA | Flagellin | 2/15 | ILNEAGISMLSQANQTPQMVSK | −3.1 | 0.0 | −3.1 | ||||||
| VQLSDASGDTMTIDSLNAK | −6.0 | 0.1 | −8.9 | |||||||||||
| BC1658 c | FlaB | Flagellin | 1/15 | TNFNGNSFLDTTATPPGKDIEIQLSDASGDTMTLK | −1.7 | −0.7 | −2.4 | |||||||
| BC1659 c | FlaC | Flagellin | 1/15 | LDHNLNNVTSQATNMAAAASQIEDADMAKEMSEMTK | −2.6 | −0.9 | −2.8 | |||||||
| Glycolysis Gluconeogenesis | BC4898 | Pgi | Glucose-6-phosphate isomerase | 1/9 | FSVLTPVGLLPIAVSGLNIEEMMK | 7.2 | 3.3 | 7.2 | ||||||
| BC5135 | Eno | Phosphopyruvate hydratase | 2/11 | LGANAILGVSMAVAHAAADFVGLPLYR | 9.5 | 2.2 | 9.6 | |||||||
| VNQIGTLTETFEAIEMAK | 1.8 | 1.6 | 1.4 | |||||||||||
| Exotoxins | BC3101 | HblB’ | Hemolysin BL, component B’ | 2/15 | GLDMVKIPFIPTLIAGGIMIGDAR | 1.5 | 0.9 | |||||||
| IPFIPTLIAGGIMIGDAR | 7.9 | 0.7 | 7.9 | |||||||||||
| BC3102 | HblB | Hemolysin BL, component B | 4/8 | SMNAYSYMLIKNPDVNFEGITINGYVDLPGR | 6.3 | 0.7 | 6.7 | |||||||
| MKETLQK | 0.3 | −1.0 | 0.4 | |||||||||||
| QLLDTLNGIVEYDTTFDNYYETMVEAINTGDGETLKEGITDLR | 8.4 | 2.2 | 7.7 | |||||||||||
| QLLDTLNGIVEYDTTFDNYYETMVEAINTGDGETLK | 7.9 | 0.9 | 7.4 | |||||||||||
| BC3103 | HblL1 | Hemolysin BL, component L1 | 3/8 | MLQDFKGK | 8.2 | 3.7 | 8.2 | |||||||
| IGELSMKADR | 1.7 | −0.3 | 2.1 | |||||||||||
| QWNTMGANYTDLLDNIDSMEDHKFSLIPDDLK | 1.9 | 0.8 | 1.6 | 8.0 | 1.5 | 7.4 | ||||||||
| BC3523 | HlyII | Hemolysin II | 1/7 | DSFNTFYGNQLFMK | 0.9 | −5.4 | 0.9 | |||||||
| BC1809 | NheA | Non hemolytic enterotoxin, A | 1/9 | VLNNNMIQIQTNVEEGTYTDSSLLQK | 7.6 | −0.3 | 8.2 | |||||||
| BC5101 | HlyI | Cereolysin | 2/7 | MTLDHYGAYVAQFDVSWDEFTFDQK | 0.8 | 1.0 | 0.4 | |||||||
| KVMVAAYK | 6.3 | 6.3 | ||||||||||||
| Toxin-like | BC5239 | EntA | Enterotoxin, cell wall binding | 1/5 | VLTAMGHDLTANPNMK | −1.4 | −1.4 | −0.6 | ||||||
| Uncategorized | BC5027 c | ErfK/srfK precursor | 1/4 | MYNNDIHWLFER | −1.6 | −0.9 | −1.0 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duport, C.; Madeira, J.-P.; Farjad, M.; Alpha-Bazin, B.; Armengaud, J. Methionine Sulfoxide Reductases Contribute to Anaerobic Fermentative Metabolism in Bacillus cereus. Antioxidants 2021, 10, 819. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050819
Duport C, Madeira J-P, Farjad M, Alpha-Bazin B, Armengaud J. Methionine Sulfoxide Reductases Contribute to Anaerobic Fermentative Metabolism in Bacillus cereus. Antioxidants. 2021; 10(5):819. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050819
Chicago/Turabian StyleDuport, Catherine, Jean-Paul Madeira, Mahsa Farjad, Béatrice Alpha-Bazin, and Jean Armengaud. 2021. "Methionine Sulfoxide Reductases Contribute to Anaerobic Fermentative Metabolism in Bacillus cereus" Antioxidants 10, no. 5: 819. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050819