
3H-1,2-Dithiole-3-Thione Protects Lens Epithelial Cells against Fructose-Induced Epithelial-Mesenchymal Transition via Activation of AMPK to Eliminate AKR1B1-Induced Oxidative Stress in Diabetes Mellitus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Reagents
2.3. Animals
2.4. Tissue Sample Collection
2.5. Immunoblotting Analysis
2.6. Blood Pressure Measurements
2.7. Immunofluorescence Staining Analysis
2.8. Statistical Analysis
3. Results
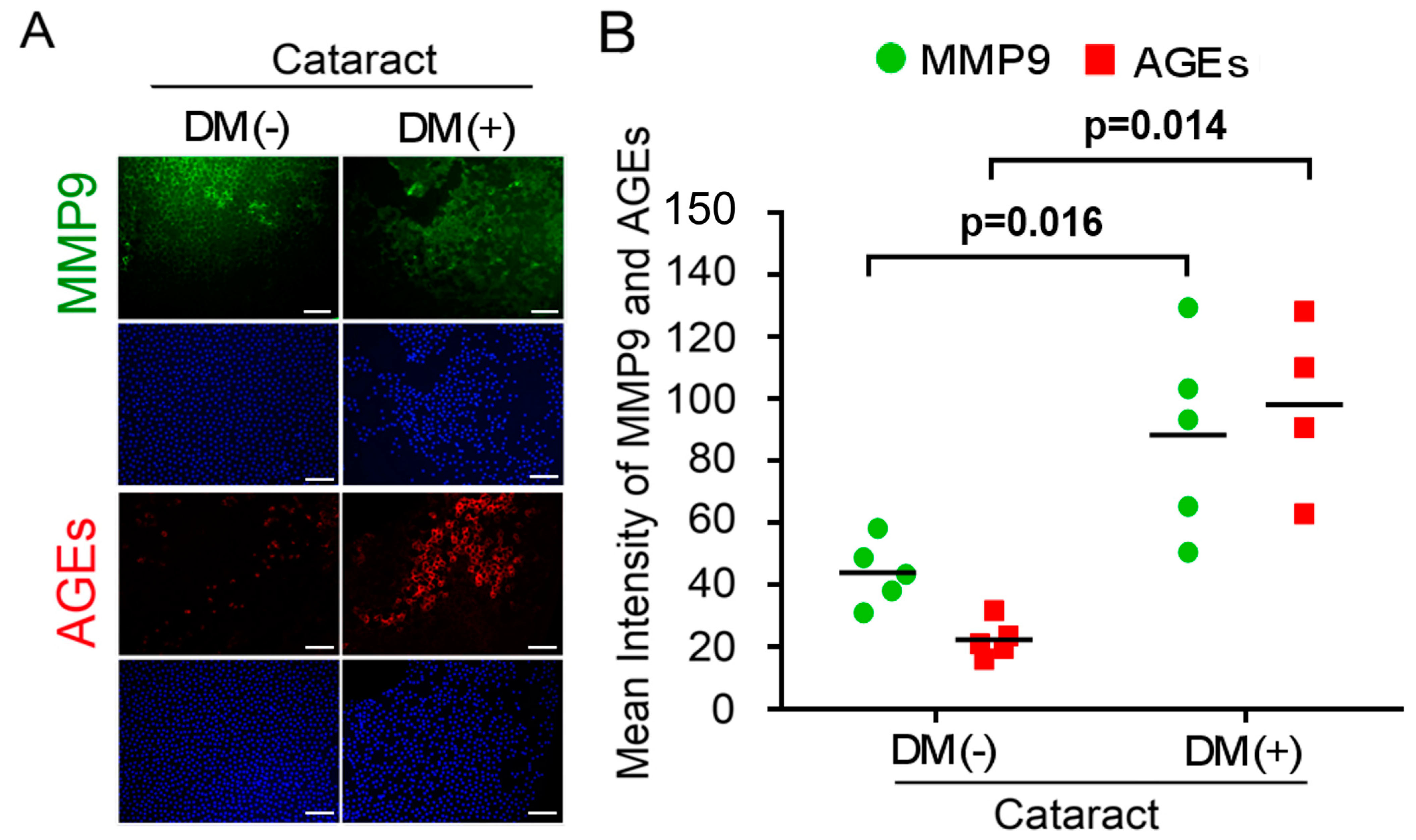
3.1. Activated AGEs and Enhanced MMP9 Production Are Involved in DM (+) Cataract LECs Pathogenesis
3.2. D3T Improves Fructose-Induced Hypertension, Does Not Prevent Fructose-Mediated Metabolic Defects
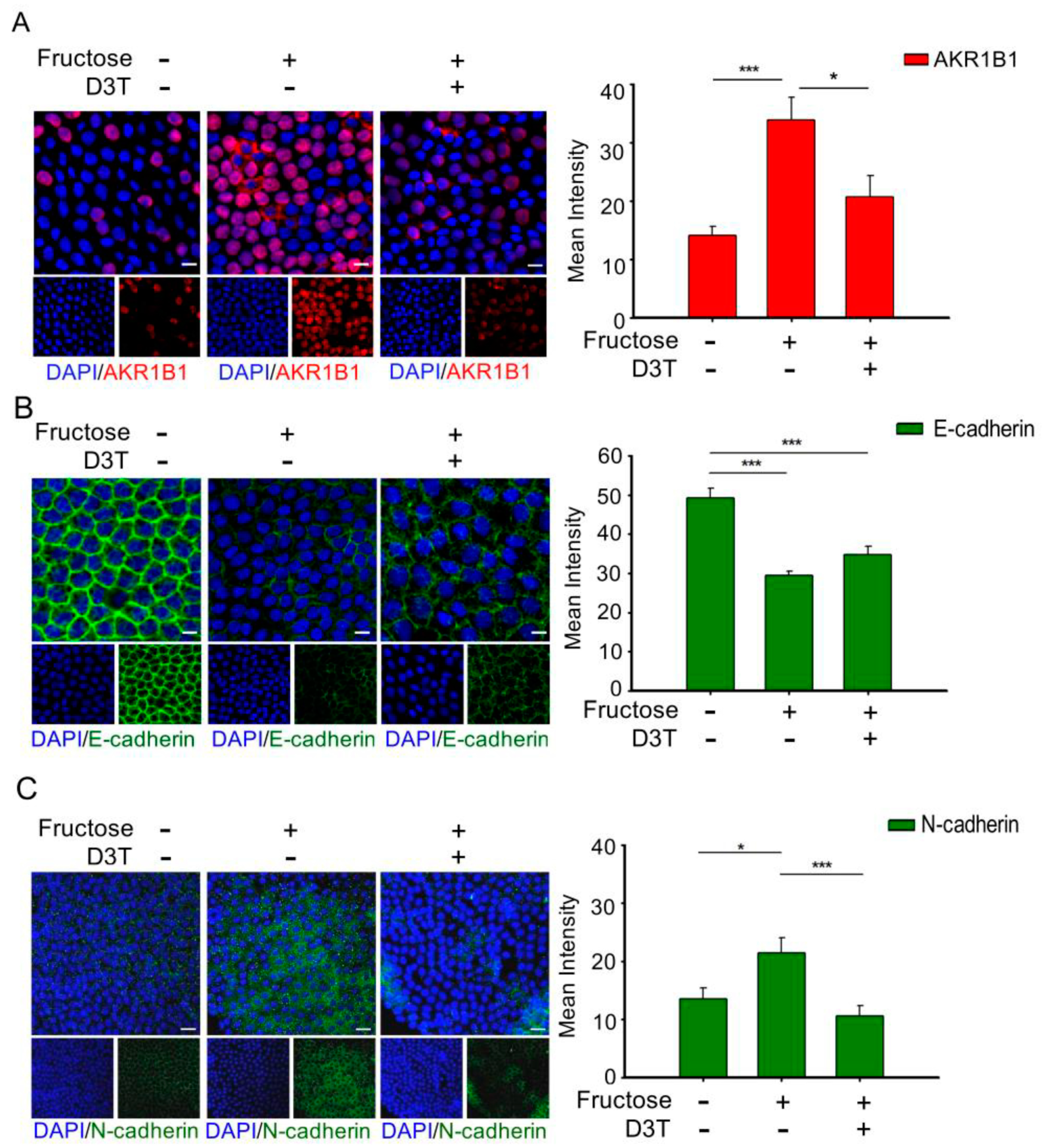
3.3. D3T Reduces AKR1B1-Induced ROS Generation and Decreases EMT in the LECs of Rats with Fructose-Induced Type 2 DM
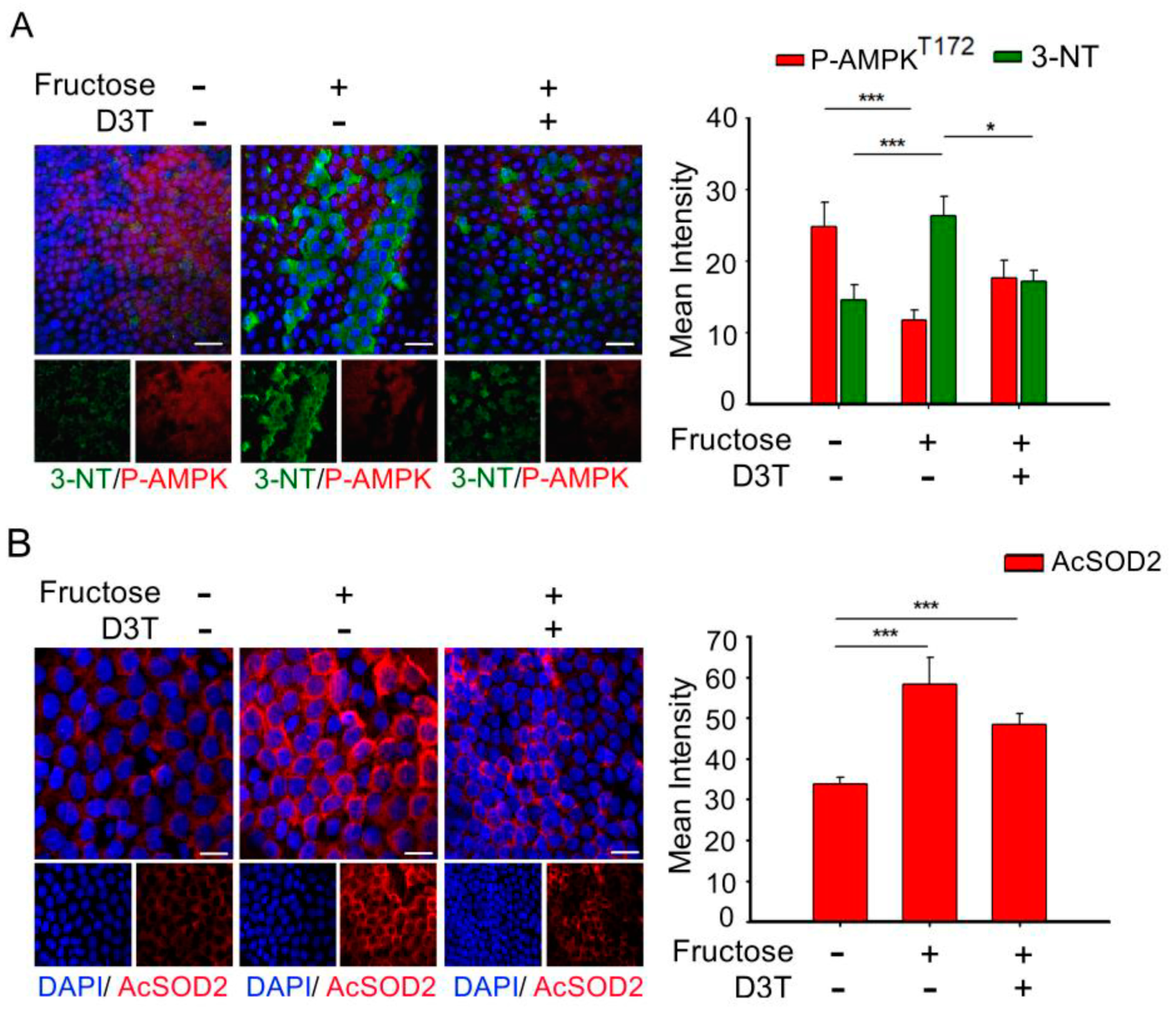
3.4. D3T Enhancement of AMP-Activated Protein Kinase (AMPK) Reduces ROS Production and SOD2 Acetylation in the Lens of Rats with Fructose-Induced Type 2 DM
3.5. D3T Reduces Snail/Slug Signaling Pathways to Ameliorate AKR1B1-Mediated Epithelial-to-Mesenchymal Transition through AMPK Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Skarbez, K.; Priestley, Y.; Hoepf, M.; Koevary, S.B. Comprehensive review of the effects of diabetes on ocular health. Expert Rev. Ophthalmol. 2010, 5, 557–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiziltoprak, H.; Tekin, K.; Inanc, M.; Goker, Y.S. Cataract in diabetes mellitus. World J. Diabetes. 2019, 10, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Boyle, J.P.; Thompson, T.J.; Gregg, E.W.; Barker, E.L.; Williamson, D.F. Projection of the year 2050 burden of diabetes in the US adult population: Dynamic modeling of incidence, mortality, and prediabetes prevalence. Popul. Health Metrics 2010, 8, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, S.; Mitchell, P.; Rochtchina, E. Five-year incidence of cataract in older persons with diabetes and pre-diabetes. Ophthalmic Epidemiol. 2004, 11, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.-T.; Chen, Y.-Y.; Chang, H.-Y.; Kung, Y.-H.; Tseng, C.-J.; Cheng, P.-W. AKR1B1-Induced Epithelial–Mesenchymal Transition Mediated by RAGE-Oxidative Stress in Diabetic Cataract Lens. Antioxidants 2020, 9, 273. [Google Scholar] [CrossRef] [Green Version]
- Drinkwater, J.J.; Davis, W.A.; Davis, T.M. A systematic review of risk factors for cataract in type 2 diabetes. Diabetes/Metab. Res. Rev. 2019, 35, e3073. [Google Scholar] [CrossRef] [Green Version]
- Becker, C.; Schneider, C.; Aballéa, S.; Bailey, C.; Bourne, R.; Jick, S.; Meier, C. Cataract in patients with diabetes mellitus—incidence rates in the UK and risk factors. Eye 2018, 32, 1028–1035. [Google Scholar] [CrossRef] [Green Version]
- Guglielmotto, M.; Aragno, M.; Tamagno, E.; Vercellinatto, I.; Visentin, S.; Medana, C.; Catalano, M.G.; Smith, M.A.; Perry, G.; Danni, O.; et al. AGEs/RAGE complex upregulates BACE1 via NF-kappaB pathway activation. Neurobiol. Aging 2012, 33, 196.e13–196.e27. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.; Adeli, K. Dietary fructose and the metabolic syndrome. Curr. Opin. Gastroenterol. 2008, 24, 204–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandarakis, S.A.; Piperi, C.; Topouzis, F.; Papavassiliou, A.G. Emerging role of advanced glycation-end products (AGEs) in the pathobiology of eye diseases. Prog. Retin. Eye Res. 2014, 42, 85–102. [Google Scholar] [CrossRef]
- Sadi, G.; Eryilmaz, N.; Tütüncüoğlu, E.; Cingir, Şahika; Güray, T. Changes in expression profiles of antioxidant enzymes in diabetic rat kidneys. Diabetes/Metab. Res. Rev. 2012, 28, 228–235. [Google Scholar] [CrossRef]
- Snow, A.; Shieh, B.; Chang, K.-C.; Pal, A.; Lenhart, P.; Ammar, D.; Ruzycki, P.; Palla, S.; Reddy, G.B.; Petrash, J.M. Aldose reductase expression as a risk factor for cataract. Chem. Interact. 2015, 234, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Grewal, A.S.; Bhardwaj, S.; Pandita, D.; Lather, V.; Sekhon, B.S. Updates on Aldose Reductase Inhibitors for Management of Diabetic Complications and Non-diabetic Diseases. Mini-Rev. Med. Chem. 2015, 16, 120–162. [Google Scholar] [CrossRef] [PubMed]
- Zablocki, G.J.; Ruzycki, P.A.; Overturf, M.A.; Palla, S.; Reddy, G.B.; Petrash, J.M. Aldose reductase-mediated induction of epitheli-um-to-mesenchymal transition (EMT) in lens. Chem. Biol. Interact. 2011, 191, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wormstone, I.M.; Wang, L.; Liu, C.S. Posterior capsule opacification. Exp. Eye Res. 2009, 88, 257–269. [Google Scholar] [CrossRef]
- Lin, S.-C.; Hardie, D.G. AMPK: Sensing Glucose as well as Cellular Energy Status. Cell Metab. 2018, 27, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.; Bissa, B.; Brecht, L.; Allers, L.; Choi, S.W.; Gu, Y.; Zbinden, M.; Burge, M.R.; Timmins, G.; Hallows, K.; et al. AMPK, a Regulator of Metabolism and Autophagy, Is Activated by Lysosomal Damage via a Novel Galectin-Directed Ubiquitin Signal Transduction System. Mol. Cell 2020, 77, 951–969.e9. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Li, C.X.; Wang, H.W.; Jiang, W.J.; Li, G.C.; Zhang, Y.D.; Luo, C.H.; Li, X.C. The Inhibition of Aldose Reductase Accelerates Liver Regen-eration through Regulating Energy Metabolism. Oxid. Med. Cell. Longev. 2020, 2020, 3076131. [Google Scholar]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G.; Group NCRRGW. Animal research: Reporting in vivo experi-ments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef]
- McGrath, J.C.; Lilley, E. Implementing guidelines on reporting research using animals (ARRIVE etc.): New requirements for publication in BJP. Br. J. Pharmacol. 2015, 172, 3189–3193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthoud, V.M.; Beyer, E. Oxidative Stress, Lens Gap Junctions, and Cataracts. Antioxid. Redox Signal. 2009, 11, 339–353. [Google Scholar] [CrossRef] [Green Version]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell. Longev. 2016, 2016, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Korol, A.; Pino, G.; Dwivedi, D.; Robertson, J.V.; Deschamps, P.A.; West-Mays, J.A. Matrix metalloproteinase-9-null mice are resistant to TGF-beta-induced anterior subcapsular cataract formation. Am. J. Pathol. 2014, 184, 2001–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, T.-C.; Liu, C.-P.; Cheng, W.-H.; Chen, B.-R.; Lu, P.-J.; Cheng, P.-W.; Ho, W.-Y.; Sun, G.-C.; Liou, J.-C.; Tseng, C.-J. Caffeine Intake Improves Fructose-Induced Hypertension and Insulin Resistance by Enhancing Central Insulin Signaling. Hypertension 2014, 63, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Cheng, P.W.; Lin, Y.T.; Ho, W.Y.; Lu, P.J.; Chen, H.H.; Lai, C.C.; Sun, G.C.; Yeh, T.C.; Hsiao, M.; Tseng, C.J.; et al. Fructose induced neurogenic hypertension mediated by overac-tivation of p38 MAPK to impair insulin signaling transduction caused central insulin resistance. Free Radic. Biol. Med. 2017, 112, 298–307. [Google Scholar] [CrossRef]
- Misra, P.; Chakrabarti, R. The role of AMP kinase in diabetes. Indian J. Med. Res. 2007, 125, 389–398. [Google Scholar]
- Mahjoub, S.; Masrour-Roudsari, J. Role of oxidative stress in pathogenesis of metabolic syndrome. Casp. J. Intern. Med. 2012, 3, 386–396. [Google Scholar]
- Li, Y.R.; Jia, Z.; Zhu, H. 3H-1,2-Dithiole-3-Thione as a Potentially Novel Therapeutic Compound for Sepsis Intervention. React. Oxyg. Species (Apex.) 2019, 8, 202–212. [Google Scholar] [CrossRef]
- Sterneck, E.; Poria, D.K.; Balamurugan, K. Slug and E-Cadherin: Stealth Accomplices? Front. Mol. Biosci. 2020, 7, 138. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Kim, S.-G.; Kim, J.-Y.; Heo, Y.-K.; Park, J.-C.; Oh, J.-S. Histologic evaluation of a retrieved endosseous implant: A case report. Int. J. Periodontics Restor. Dent. 2013, 33, e32–e36. [Google Scholar] [CrossRef] [Green Version]
- Tan, A.G.; Kifley, A.; Holliday, E.G.; Klein, B.E.K.; Iyengar, S.K.; Lee, K.E.; Jun, G.R.; Cumming, R.G.; Zhao, W.; Wong, T.Y.; et al. Aldose Reductase Polymorphisms, Fasting Blood Glucose, and Age-Related Cortical Cataract. Investig. Opthalmol. Vis. Sci. 2018, 59, 4755–4762. [Google Scholar] [CrossRef] [PubMed]
- Stitt, A.W. Advanced glycation: An important pathological event in diabetic and age related ocular disease. Br. J. Ophthalmol. 2001, 85, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Ramana, K.V. Aldose reductase: New insights for an old enzyme. Biomol. Concepts 2011, 2, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashim, Z.; Zarina, S. Advanced glycation end products in diabetic and non-diabetic human subjects suffering from cataract. AGE 2010, 33, 377–384. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Wu, T.-T.; Ho, C.-Y.; Yeh, T.-C.; Sun, G.-C.; Kung, Y.-H.; Wong, T.-Y.; Tseng, C.-J.; Cheng, P.-W. Dapagliflozin Prevents NOX- and SGLT2-Dependent Oxidative Stress in Lens Cells Exposed to Fructose-Induced Diabetes Mellitus. Int. J. Mol. Sci. 2019, 20, 4357. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Hao, M.; Li, C.; Wu, W.; Wang, W.; Ma, Z.; Yang, T.; Zhang, N.; Isaac, A.T.; Zhu, X.; et al. Quercetin inhibited epithelial mesenchymal transition in diabetic rats, high-glucose-cultured lens, and SRA01/04 cells through transforming growth factor-beta2/phosphoinositide 3-kinase/Akt pathway. Mol. Cell Endocrinol. 2017, 452, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Li, K.R.; Yang, S.Q.; Gong, Y.Q.; Yang, H.; Li, X.M.; Zhao, Y.X.; Yao, J.; Jiang, Q.; Cao, C. 3H-1,2-dithiole-3-thione protects retinal pigment epithelium cells against Ultra-violet radiation via activation of Akt-mTORC1-dependent Nrf2-HO-1 signaling. Sci. Rep. 2016, 6, 25525. [Google Scholar] [CrossRef]
- Kumar, H.; Kim, I.-S.; More, S.V.; Kim, B.; Choi, D.-K. Natural product-derived pharmacological modulators of Nrf2/ARE pathway for chronic diseases. Nat. Prod. Rep. 2014, 31, 109–139. [Google Scholar] [CrossRef]
- Wautier, M.-P.; Chappey, O.; Corda, S.; Stern, D.M.; Schmidt, A.M.; Wautier, J.-L. Activation of NADPH oxidase by AGE links oxidant stress to altered gene expression via RAGE. Am. J. Physiol. Metab. 2001, 280, E685–E694. [Google Scholar] [CrossRef]
- Stochelski, M.A.; Wilmanski, T.; Walters, M.; Burgess, J.R. D3T acts as a pro-oxidant in a cell culture model of diabetes-induced peripheral neuropathy. Redox Biol. 2019, 21, 101078. [Google Scholar] [CrossRef]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhang, M.; Liang, B.; Xu, J.; Xie, Z.; Liu, C.; Viollet, B.; Yan, D.; Zou, M.H. AMPKalpha2 deletion causes aberrant expression and activation of NAD(P)H oxidase and consequent endothelial dysfunction in vivo: Role of 26S proteasomes. Circ Res. 2010, 106, 1117–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, S.; Ozawa, Y.; Kurihara, T.; Sasaki, M.; Yuki, K.; Miyake, S.; Noda, K.; Ishida, S.; Tsubota, K. Roles of AMP-Activated Protein Kinase in Diabetes-Induced Retinal Inflammation. Investig. Opthalmol. Vis. Sci. 2011, 52, 9142–9148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xu, Z.; Ji, L.; Guo, L.; Liu, J.; Feng, K.; Xu, Y.; Zhu, D.; Jia, W.; Ran, X.; et al. Direct medical costs for patients with type 2 diabetes in 16 tertiary hospitals in urban China: A multicenter prospective cohort study. J. Diabetes Investig. 2018, 10, 539–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, M.S.; Kim, W.D.; Lee, K.Y.; Kim, J.H.; Koo, J.H.; Kim, S.G. AMPK Facilitates Nuclear Accumulation of Nrf2 by Phosphorylating at Serine 550. Mol. Cell. Biol. 2016, 36, 1931–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinman, B. The International Diabetes Federation World Diabetes Congress 2015. Eur. Endocrinol. 2015, 11, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Group (n = 6) | DM Group (n = 6) | p Value | |
|---|---|---|---|
| Age (years) | 66.0 ± 3.5 | 66.8 ± 6.1 | 0.871 a |
| Sex (male: female) | 2:4 | 2:4 | 1.000 b |
| BMI | 24.0 ± 2.7 | 25.3 ± 2.6 | 0.522 a |
| HBA1C | - | 9.0 ± 1.4 | - |
| Parameter/Group | Control | Fructose 8 W | Fructose 8 W + D3T |
|---|---|---|---|
| Systolic blood pressure (mm Hg) | 100.6 ± 2.0 | 161.4 ± 4.2 ** | 114.1 ± 2.9 ## |
| Fasting serum glucose (mg/dL) | 98.7 ± 6.1 | 150.0 ± 2.0 ** | 159.7 ± 6.8 ** |
| dHDL (mg/dL) | 67.8 ± 1.1 | 58.8 ± 2.2 | 74.8 ± 0.9 ## |
| Triglycerides (mg/dL) | 77.8 ± 7.8 | 156.0 ± 8.7 ** | 148.7 ± 9.0 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, T.-T.; Chen, Y.-Y.; Ho, C.-Y.; Yeh, T.-C.; Sun, G.-C.; Tseng, C.-J.; Cheng, P.-W. 3H-1,2-Dithiole-3-Thione Protects Lens Epithelial Cells against Fructose-Induced Epithelial-Mesenchymal Transition via Activation of AMPK to Eliminate AKR1B1-Induced Oxidative Stress in Diabetes Mellitus. Antioxidants 2021, 10, 1086. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10071086
Wu T-T, Chen Y-Y, Ho C-Y, Yeh T-C, Sun G-C, Tseng C-J, Cheng P-W. 3H-1,2-Dithiole-3-Thione Protects Lens Epithelial Cells against Fructose-Induced Epithelial-Mesenchymal Transition via Activation of AMPK to Eliminate AKR1B1-Induced Oxidative Stress in Diabetes Mellitus. Antioxidants. 2021; 10(7):1086. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10071086
Chicago/Turabian StyleWu, Tsung-Tien, Ying-Ying Chen, Chiu-Yi Ho, Tung-Chen Yeh, Gwo-Ching Sun, Ching-Jiunn Tseng, and Pei-Wen Cheng. 2021. "3H-1,2-Dithiole-3-Thione Protects Lens Epithelial Cells against Fructose-Induced Epithelial-Mesenchymal Transition via Activation of AMPK to Eliminate AKR1B1-Induced Oxidative Stress in Diabetes Mellitus" Antioxidants 10, no. 7: 1086. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10071086